
Abstract
Aims: Vascular endothelial growth factor (VEGF) protein plays an important role in tumor development and progression. Polymorphisms in the VEGF gene may lead to over- or underexpression of the protein and may be associated with either risk or progression of malignancy. The aim of this case-control study is to identify and quantify the correlation between VEGF polymorphisms and renal cell carcinoma (RCC). Results: Restriction fragment length polymorphism methods were used for the analysis of VEGF polymorphisms at −2578 and +936 positions in the promoter and 3′-untranslated regions, respectively. The VEGF −2578 A-allele was associated with an increased risk of RCC (odds ratio: 1.6; 95% CI: 1.2-2.3) and A-carrier genotypes were strongly correlated (odds ratio: 2.7; 95% CI: 1.5-4.7) with higher risk. Comparison of VEGF +936 C/T polymorphism between patient and control groups revealed no association with renal carcinoma. Both VEGF −2578 C/A and VEGF +936 C/T polymorphisms showed no significant association with the histopathological parameters of RCC. Conclusions: This study shows that VEGF −2578 A-allele and A-carrier genotypes are associated with an increased risk of RCC. In groups with higher incidence of RCC, a screening test for this polymorphism may be recommended in conjunction with other established markers.
Introduction
The cellular pathways involved in the pathogenesis of RCC lead to the overexpression of vascular endothelial growth factor (VEGF) protein. The protein itself is a potent mitogen and plays a critical role in angiogenesis (Ferrara and Davis-Smyth, 1997). Both in vitro and in vivo experiments have shown that increased VEGF expression is associated with tumor development and progression (Kawai et al., 2002). Some of the promising new medical treatments for metastatic renal cancer target VEGF (Patard et al., 2009). These targeted therapies include either monoclonal antibodies against VEGF (e.g., Bevacizumab) or they inhibit the VEGF receptor (e.g., Sorafenib) (Motzer and Bukowski, 2006).
The functional polymorphisms, that is, the gene variants that effect the expression and/or the function of the encoded proteins, are most likely to contribute to an individual's susceptibility to a disease. Further, these polymorphisms may also be responsible for the differences in severity of the disease and response to specific medical regimens (Balasubramanian et al., 2002a). Therefore, before the administration of targeted therapies it is essential to establish whether or not various polymorphisms of the gene play a role in increasing the risk for the disease and in its progression.
The VEGF gene has been localized on chromosome 6p21.3 and at least 30 single-nucleotide polymorphisms have been described (Watson et al., 2000). Polymorphisms in either the regulatory or coding sequence of VEGF may be responsible for the differential expression of the gene. Different studies have shown association of −634C/G in the 5′ untranslated region (UTR), −1154G/A and −2578C/A polymorphisms in the promoter region, and +936C/T in the 3′ UTR, with varied VEGF expression and, ultimately, with various cancers (Balasubramanian et al., 2002a).
In this study, the role of two of these polymorphisms, −2578 C/A and +936 C/T, in the etiology, pathology, and prognosis of RCC in the Pakistani population was investigated.
Materials and Methods
Sample collection
Blood samples of 143 RCC patients who underwent tumor nephrectomy at the Sindh Institute of Urology and Transplantation (SIUT), Karachi, Pakistan, were collected from March 2009 till July 2010. For controls, blood samples from 106 age-matched individuals were selected. Specifically, the controls had no personal history of RCC in the medical records. Five milliliters of blood samples in EDTA-coated tubes were stored at 4°C until DNA isolation.
Informed consent was obtained from all the participants and the project was approved by the Ethical Review Committee, SIUT.
Histopathological classification
The tumor tissues were histopathologically examined for tumor classification, and grading was carried out. Tumors were staged based upon the TNM staging of the Union Contre le Cancer as well as the Robson Stage for prognosis of RCC. The grading of the tumors was carried out according to the World Health Organization recommendations. The results were confirmed by a second blinded evaluation.
DNA isolation
DNA was isolated according to the standard phenol-chloroform method (Sambrook and Russell, 2001). The final DNA precipitation was carried out with an equal volume of isopropanol in the presence of 1/10th volume of 10 M ammonium acetate. The precipitates were washed with 70% ethanol and resuspended in 10 mM Tris-EDTA (TE) buffer. DNA purity and concentration were spectrophotometrically determined and samples were stored at −20°C.
Genotyping
The genotyping was carried out after the amplification of the target DNA fragment by PCR followed by restriction fragment length polymorphism.
In case of −2578 C/A, the DNA fragment was amplified using 65 ng of total genomic DNA in 20 μL PCRs containing 1.5 mM Mg++, 0.1 mM dNTPs, 1.2 U Taq and 2 μM of each primer (−2578 C/A: forward, 5′-GGA TGG GGC TGA CTA GGT AAG-3′; reverse, 5′-AGC CCC CTT TTC CTC CAA C-3′). PCR was carried out using a thermal cycler (GeneAmp® PCR System 9700; Applied Biosystems) and the reaction conditions were an initial incubation of 94°C for 5 min; 10 cycles of 94°C for 1 min, 69°C for 1 min, 72°C for 1 min; 20 cycles of 94°C for 1 min, 59°C for 1 min, and 72°C for 1 min; and a final extension at 72°C for 10 min. The PCR product was a fragment of 325 bp. About 15 μL of the amplified products were then digested overnight at 37°C with 2 units of BglII (Fermantas, Life Sciences). The digested products were separated on 2.5% agarose gel in the presence of ethidium bromide. The recognition site for BglII is A*GATCT; therefore, the presence of A-allele at the dimorphic site yielded two fragments after restriction digestion. Thus, the homozygous A alleles (i.e., AA genotype) were represented by DNA bands with 202- and 123-bp fragments; an uncut fragment of 325 bp indicated homozygous C alleles (i.e., CC genotype) and heterozygous genotype CA was displayed by a combination of 325-, 202-, and 123-bp fragments (Fig. 1a).
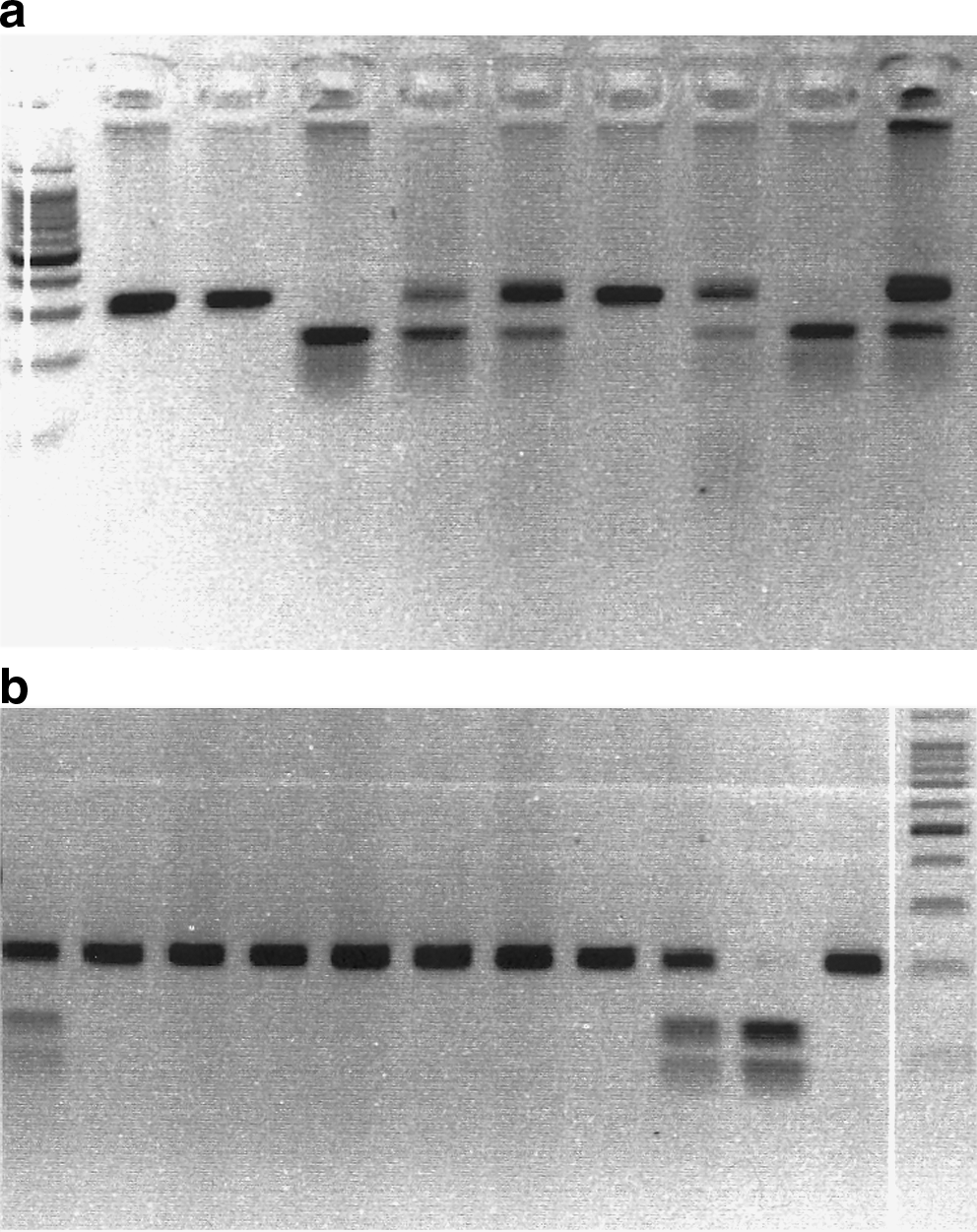
For +936C/T polymorphism, PCR used 60 ng of total genomic DNA in 15 μL PCR mixtures containing 1.5 mM Mg++, 0.1 mM dNTPs, 1.35 U Taq, and 0.6 μM of each primer (+936C/T: forward, 5′-AAG GAA GAG GAG ACT CTG CGC AGA GC-3′; reverse, 5′-TAA ATG TAT GTA TGT GGG TGG GTG TGT CTA CAG-3′). Reaction conditions were same as described earlier. The PCR resulted in a product of 208 bp. Ten microliters of amplified products were then digested overnight at 37°C with 4 units of NlaIII (Fermantas, Life Sciences). The digested products were resolved on 2.5% agarose gel as described earlier. The recognition site for NlaIII is CATG*; therefore, after restriction, the presence of the T-allele at the dimorphic site yielded two fragments. The homozygous T alleles (i.e., TT genotype) displayed DNA fragments of 122 and 86 bp, the heterozygous CT genotype was indicated by a combination of 208-, 122-, and 86-bp fragments, and an uncut 208-bp fragment represented homozygous C-allele (i.e., CC genotype) as shown in Figure 1b.
Statistical analysis
Data pertaining to participants were coded and analyzed using the Statistical Package for Social Sciences (SPSS®) version 13 for Windows®. Demographics of cases and controls were calculated and compared. The association of VEGF alleles and genotypes with RCC and clinical parameters were investigated using the chi-square test of independence with appropriate degree of freedom. The strength of a genetic association was measured by odds ratio (OR) and 95% confidence interval. For all statistical analyses, p-values <0.05 were considered significant.
Results
Baseline data of participants and clinicopathologic characteristics of RCC patients
The study comprised 249 participants, with 143 RCC patients and 106 controls. The latter had no personal history of RCC or any other cancer and were age matched. The data pertaining to participants are given in Table 1a. The tumor characteristics of RCC patients, consisting of size, grade, and histopathological classification are listed in Table 1b.
The incidence of RCC was predominant in males, with a ratio of 1.5:1; however, this difference was not statistically significant.
Association of VEGF −2578 A-allele and A-carrier genotypes with increased risk of RCC
The distributions of genotypes and allele frequencies of VEGF −2578C/A polymorphism are shown in Table 2. In both the controls and the patients, the genotypes were in Hardy-Weinberg equilibrium (controls: χ2=4.24, 2 df, p=0.08; RCC patients: χ2=1.28, 2 df, p=0.60).
RCC, renal cell carcinoma.
The frequencies of A-allele were 0.39±0.05 and 0.51±0.04 in controls and RCC patients, respectively. The chi-square test of independence showed that A-allele was significantly associated with the risk of RCC (χ2=6.5, 1 df, p=0.011), having an OR of 1.6 (95% CI: 1.2-2.3).
The percentages (numbers) of CC genotypes in controls and patients were 41.5% (44/106) and 20.1% (30/143), respectively. On the other hand, the percentages (numbers) of A-carrier genotypes in controls and patients were 58.5% (62/106) and 79.0% (113/143), respectively. The comparison of CC genotypes with A-carrier genotypes, that is, CC versus CA+AA, revealed that the presence of at least one A-allele in the genotype was strongly associated with an increased risk of RCC (χ2=12.3, 1 df, p=0.001) and conferred an OR of 2.7 (95% CI: 1.5-4.7).
No correlation of VEGF −2578C/A polymorphism with clinical variables of RCC
VEGF −2578 allelic and genotypic frequencies did not show any significant association with either the age of the patient, tumor size, tumor grade, or tumor stage (Table 3).
Tumor grade was unknown for 22 samples.
Tumor stage was unknown for 66 samples.
VEGF +936C/T polymorphism and RCC
VEGF +936C/T polymorphism lacked association with RCC (χ2=1.15, 1 df, p=0.36), having an OR of 1.5 (95% CI: 0.7-3.3). Also, the haplotype analyses in combination with −2578 C/A polymorphism did not reveal any significant result (data not shown).
Discussion
The variations in the susceptibility and severity of complex disorders, such as cancers, may be attributed to functional polymorphisms in the candidate genes. Association studies that identify such markers are influenced by the genetic make-up of target population. It is therefore necessary to analyze the putative genetic markers across different populations before they can be used as either prognostic markers or therapeutic targets.
The VEGF protein is considered to play a major role in cancer progression. Evidence shows that it stimulates proliferation and migration of endothelial cells resulting in angiogenesis (Veikkola and Alitalo, 1999). Certain polymorphisms in the promoter region, 5′ and 3′ UTR, have been shown to affect either the expression or the function of VEGF protein (Balasubramanian et al., 2002a). In this study, we have investigated the association of two of these polymorphisms, −2578C/A in the promoter region and +936C/T in the 3′ UTR, with RCC.
Our results show that the A-allele of −2578C/A polymorphism is associated with increased RCC risk in our population. Further, the analysis of genotypic frequencies revealed that subjects with at least one A-allele, that is, those with CA and AA genotypes, were 2.7 times more likely to suffer from RCC than those with CC genotype.
A correlation between +936C/T polymorphism and VEGF protein levels has been shown, wherein +936 T-allele is correlated with decreased VEGF plasma levels (Renner et al., 2000). Therefore, this polymorphism may also affect the incidence and severity of different cancers. A study from Korea reported a gender-specific association between this polymorphism and stomach cancer (Bae et al., 2008). However, a study from Japan (Abe et al., 2002) and two more studies from Europe reported no association with breast cancer (Balasubramanian et al., 2002b; Jin et al., 2005). Similarly, the present study did not find any correlation between this polymorphism and incidence/tumor characteristics of RCC.
To our knowledge, this is the first study that reports a highly significant association of the A-allele and A-carrier genotypes of VEGF −2578C/A polymorphism with an increased risk for RCC. Recently, a study from France reported strong association with another polymorphism, −460C/T, in the 5′ UTR of VEGF gene (Bruyère et al., 2010) with RCC. Interestingly, it appears that this locus (VEGF −460) is in linkage disequilibrium with the VEGF −2578 locus, at least in the Caucasian population. Thus, this study in combination with the aforementioned French work suggests the association of polymorphisms in the 5′ UTR and promoter region of VEGF gene with an increased risk of RCC.
Another study from Japan analyzed the effect of some polymorphisms on the characteristics and prognosis of RCC. They reported that the CA and CA+AA genotypes of −2578C/A polymorphism were weakly associated with less-frequent lymph node metastasis and favorable cancer-specific survival (Kawai et al., 2002). However, our results do not show any significant effect of this polymorphism on tumor characteristics, such as tumor stage, size, and grade.
The association of VEGF −2578 A-allele and A-carrier genotypes with increased risk of RCC is most likely due to the modification of VEGF protein expression. Indeed, a study published in 2004 reported that A-allele was correlated with higher VEGF levels (Koukourakis et al., 2004). However, in another study, C-allele was associated with higher VEGF levels (Shahbazi et al., 2002). Therefore, further studies, both in vitro and in vivo, need to be carried out to establish the effect of polymorphisms on protein levels.
As our study shall proceed, more data are likely to emerge in the next few years. Further, the analyses of −1154C/G polymorphism in the promoter region and −634C/G in the 5′ UTR of VEGF gene are also in progress.
The present study is a part of an ongoing effort to identify the genetic components involved in the pathogenesis and progression of RCC. Further studies are needed to establish the findings reported here and to uncover the mechanisms through which these genetic variations contribute to an increased risk for RCC.
Footnotes
Acknowledgments
This study was supported by a core grant of SIUT. The authors specially thank the patients who participated in this study.
Disclosure Statement
No competing financial interests exist.