
Abstract
Background: ISL1, as a member of the LIM homeodomain transcription factor family, is expressed in a distinct population of undifferentiated cardiac progenitors and plays a pivotal role in cardiogenesis. Lacking ISL1 expression results in growth arrest or displays profound defects in heart development, including atria, ventricle, and the inflow and outflow tracts, which constitute a major form of congenital heart disease (CHD). Recently, an important study by Stevens et al. found that genetic variation in ISL1 is associated with risk of CHD in white and black/African American populations; this observation led us to hypothesize that ISL1 common variants might influence susceptibility to sporadic CHD in our Chinese population. Methods: We conducted a case-control study of CHD in Chinese to test our hypothesis by genotyping ISL1 common variant rs1017 in 1003 CHD cases and 1012 non-CHD controls. Results: We found that rs1017 was not associated with the risk of CHD (p=0.213). When we performed stratified analyses according to subjects' age, sex, and CHD classifications, we found no overall heterogeneity of risk in different subgroups. Conclusions: This is the first study which indicates that ISL1 common variant rs1017 may not play a role in sporadic CHD susceptibility in the Chinese population.
Introduction
C
Recently, ISL1, a LIM homeodomain transcription factor, has become an important molecular marker for the second heart field, which is expressed early in the cardiac progenitor population and functions near the top of a transcriptional pathway essential for heart development (Kang et al., 2009). ISL1+ genetic marking observation suggested that embryonic ISL1-expressing cells are multipotent cardiac progenitors that give rise to three major cardiac lineages, including cardiomyocytes, smooth muscle, and endothelial cells (Laugwitz et al., 2005; Moretti et al., 2006). Moreover, lineage tracing analysis further demonstrated that ISL1 descendants contribute extensively to the heart, including to parts of the inflow tract region, the atria, and the left ventricle, as well as the outflow tract and the right ventricle (Cai et al., 2003).
Gain- and loss-of-function experiments revealed that ISL1 is required for normal heart development by regulating cardiac gene expression and morphogenesis of the heart (Dodou et al., 2004; Lin et al., 2006; Cohen et al., 2007; Lin et al., 2007; Ma et al., 2008). Ablation of ISL1 in mice results in embryonic growth arrest at E9.5 and lethality by E10.5 (Pfaff et al., 1996; Cai et al., 2003). Additionally, ISL1 mutant mice display several common forms of cardiovascular malformations, including persistent truncus arteriosus, transposition of the great arteries, tetralogy of Fallot, ventricular septal defect, and atrial septal defect, indicating their potential role in diverse forms of human CHD (Lin et al., 2006; Bu et al., 2009). Although previous evidence proved that dysregulation of ISL1 brings about the occurrence of CHD, the mechanism about which factors alter ISL1 expression remains unclear. Interestingly, a recent study gives us a clue that the most common genetic variation, single-nucleotide polymorphisms (SNPs) in ISL1 might be functional and modify ISL1 expression, causing significant increased risk of CHD in white and black/African American populations (Stevens et al., 2010).
Thus, it will be of particular interest to identify whether a common variant in ISL1 influences susceptibility to sporadic CHD in our Chinese population.
Materials and Methods
Study population
The study was approved by the institutional review board of Nanjing Medical University, Nanjing, China. The study population and subject characteristics were previously described (Xu et al., 2009, 2010). In brief, this hospital-based case-control study included 1003 confirmed CHD patients and 1012 non-CHD controls, frequency matched to the cases on age, sex, and residential areas. The CHD cases were consecutively recruited from the First Affiliated Hospital of Nanjing Medical University and the Affiliated Nanjing Children's Hospital of Nanjing Medical University, Nanjing, China, between March 2006 and July 2008. All the cases had nonsyndromic CHD diagnosed by ultrasound and confirmed in surgical operations. Cases who had structural malformations involving another organ system or known chromosomal abnormalities were excluded. Exclusion criteria also included a positive family history of CHD in a first-degree relative (parents, siblings, and children), maternal diabetes mellitus, phenyl ketonuria, maternal teratogen exposures (e.g., pesticides and organic solvents), and maternal therapeutic drug exposures during the intrauterine period. Controls were non-CHD outpatients from the same geographic area who were frequency matched to the cases on age and sex. They were recruited from the hospitals just referred to during the same time period; most had a diagnosis of trauma or infection. Controls with congenital anomalies were excluded. All subjects were genetically unrelated ethnic Han Chinese. After informed consent had been directly obtained or from the parents, subjects and/or their parents were personally interviewed by trained interviewers using a structured questionnaire. After the interview, ∼2 mL of venous blood were collected from each subject.
SNP selection and genotyping
We used the public SNP database (the NCBI dbSNP database, build 127; www.ncbi.nlm.nih.gov/projects/SNP/) to select potential functional common (i.e., minor allele frequency >0.05) SNP in the ISL1 gene including exon, 3′ UTR and 5′ UTR region in our Chinese population. According to these criteria, we identified only one SNP rs1017, which was located within the ISL1 gene 3′ UTR region.
Genomic DNA was isolated from leukocyte pellets of venous blood by proteinase K digestion and followed by phenol-chloroform extraction. SNPs were genotyped using the TaqMan assay on the ABI PRISM 7900 HT platform (Applied Biosystems, Inc.). Approximately equal numbers of case and control samples were assayed in each 384-well plate with two or no template controls. Primers and probes were available on request. Genotyping was performed by blinding the case or control status. Five percent of the samples were randomly selected for repeat genotyping as blind duplicates, and the reproducibility was 100%.
Statistical analyses
Differences in the distributions of selected variables, frequencies of the genotype of ISL1 rs1017 polymorphism between the cases and controls were evaluated using the χ2 or Student's t-test. The associations between the genotype and the risk of CHD were estimated by adjusted odds ratio (OR) and their 95% confidence interval (CI) from logistic regression analyses, with the adjustment for age and sex. The Hardy-Weinberg equilibrium was tested by a goodness-of-fit χ2 test to compare the observed genotype frequencies with the expected ones among control subjects. All of the statistical analyses were performed with Statistical Analysis System software (v.9.1.3e; SAS Institute).
Results
The characteristics of the CHD cases and controls included in the analysis are summarized in Table 1. There were no statistically significant differences between cases and controls in terms of age and sex. Traditionally, CHD was classified into five broad categories: cyanotic heart disease, left-sided obstruction defects, septation defects, patent ductus arteriosus, and other complex abnormalities. Of the 1003 CHD cases, 260 (25.9%) had cyanotic heart disease, 633 (63.1%) had septation defects, 49 (4.9%) had patent ductus arteriosus, 56 (5.6%) had other complex abnormalities, and only 5 (0.5%) had left-sided obstruction defects.
CHD, congenital heart disease.
The genotype distributions of the SNP rs1017 in the cases and the controls are shown in Table 2. The observed genotype frequency was in agreement with that expected under the Hardy-Weinberg equilibrium in the controls. Compared with rs1017 wild-type AA, rs1017 AT was not associated with the risk of CHD (OR=0.88, 95% CI=0.71-1.08; p=0.213). A similar result was also observed for rs1017 homozygote TT (OR=0.91, 95% CI=0.48-1.75; p=0.783). Taken together, there was no appreciable difference between CHD risk and variant genotypes of ISL1 rs1017 in the recessive genetic model (OR=0.88, 95% CI=0.72-1.08; p=0.213).
Adjusted by age and sex.
OR, odds ratio; CI, confidence interval.
To further evaluate the relevance of the rs1017 SNP, we performed stratified analyses according to subjects' age, sex, and CHD classifications, we found no overall heterogeneity of risk in different subgroups (two other CHD classification methods were also used; Fig. 1).
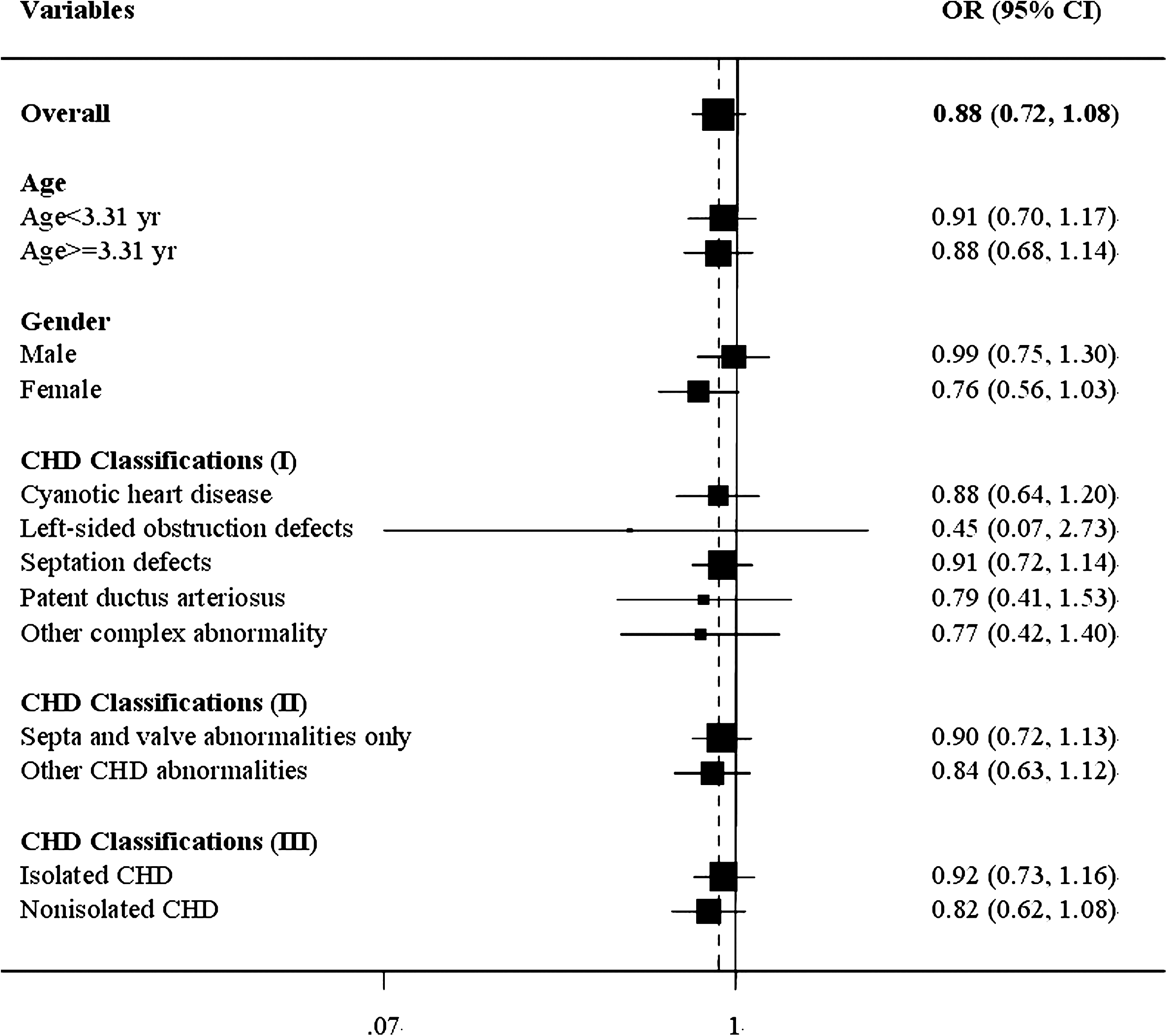
Forest plot representing the dichotomized effects of ISL1 rs1017 and CHD susceptibility.
Discussion
CHDs arises from abnormal heart development during embryogenesis. The regulatory mechanisms involved in establishing the early heart and regulating its morphogenesis have been extensively studied (Bajolle et al., 2009). The heart is derived from mesoderm that responds to signals from the surrounding tissue to develop into two different heart fields (Kelly and Buckingham, 2002; Buckingham et al., 2005). The earliest population of cardiac progenitors, referred to as the first heart field, gives rise to the linear heart tube and subsequently, parts of the atria and the left ventricle. Cells of the second heart field are derived from the pharyngeal mesoderm that lies anterior and dorsal to the heart tube and, subsequently, gives rise to the outflow tract, right ventricle, part of the atria, and a few portions of the left ventricle (Mjaatvedt et al., 2001; Waldo et al., 2001; Brand, 2003).
The LIM homeodomain transcription factor ISL1 marks a cell population that constitutes the second heart field and should, thus, represent a true endogenous cardiac progenitor cell (Bu et al., 2009). In vivo cell lineage tracing in mouse embryos has confirmed that ISL1+ progenitors contribute to more than two-thirds of the cells in the embryonic heart (Cai et al., 2003; Moretti et al., 2006; Sun et al., 2007) and suggested that purified populations of ISL1+ progenitor cells are capable of self-renewal and differentiation into cardiomyocytes, smooth muscle, and endothelial lineages (Qyang et al., 2007), and throughout the inflow tract region, the atria, and the left ventricle, as well as the outflow tract and the right ventricle (Genead et al., 2010).
Functional analysis showed that ISL1 is required for proliferation, survival, and migration of cardiac progenitors into the forming heart. Inhibition of ISL1 results in growth arrest or exhibits severely defects in the outflow tract, right ventricle, atria, and inflow tract (Brade et al., 2007). More importantly, a recent publication by Stevens et al. (2010) demonstrated that two different SNPs (rs1017, rs3762977) in ISL1 contribute to risk of CHD in white or black/African American population, respectively. Specifically, within ISL1, rs1017 significantly increased the risk of CHD in a white population, while no association was observed in a black/African American population. Similarly, increased CHD risk was observed for rs3762977 in a black/African American population but not for a white population (Stevens et al., 2010).
To investigate the relationship between an ISL1 common variant and CHD risk in the Chinese population, we searched the public SNP database (the NCBI dbSNP database, build 127; www.ncbi.nlm.nih.gov/projects/SNP/) to select a common (i.e., minor allele frequency >0.05) SNP in the ISL1 gene in our Chinese population and found only one SNP rs1017 agreement to these criteria. Logistic-regression analysis showed that no association was found between rs1017 and CHD risk. To diminish the potential confounding agents, we performed stratified analyses according to subjects' age, sex, and CHD classifications, and no overall heterogeneity of risk in different subgroups was observed.
Differences in risk estimate for ISL1 polymorphism rs1017 in association with CHD might be caused by multiple factors, including etiologic heterogeneity between populations, study design, type of heart defects, and lack of information on potential effect modifiers. In the present study, the case-control design reduced the possibility of finding a chance association. Strengths of the study also include a relatively large sample size, clear exclusion criteria for cases, and matching criteria between cases and controls.
In summary, cardiac development is a complicated process, involving the expression of many genes at different times, spaces, and orders. Now that some of the genes involved in the main forms of CHD have been identified, a new challenge is to find which common polymorphisms in these genes might be causal.
Footnotes
Acknowledgments
This work was supported in part by National Natural Science Foundation of China (30872544 and 81170158); The Ph. D Program Foundation of Ministry of Education of China (20093234110006); Jiangsu Provincial Natural Science Foundation of China (BK2007252); Jiangsu Province Health Department Program Grant (H200821); and Jiangsu Postdoctoral Sustentation Fund (0801025B).
Author Disclosure Statement
No competing financial interests exist.