
Abstract
The genetic polymorphisms in DNA repair genes might affect the repair activities of the enzymes, predisposing individuals to cancer risk. Due to these genetic variants, interethnic differences in DNA repair capacity were observed in various populations. Hence, our study aimed to determine the prevalence of three nonsynonymous single-nucleotide polymorphisms (SNPs) in an X-ray repair cross-complementation group 1 gene (XRCC1) (Arg194Trp, Arg280His, and Arg399Gln) in a healthy Tunisian population (TUN) and to compare that with HapMap (www.hapmap.org) populations. Also, we predicted their eventual functional effect based on the protein conservation analysis by
Introduction
DNA
X-ray repair cross-complementation gene 1 (XRCC1), one of the DNA repair proteins, plays an important role in base excision repair (BER). XRCC1 is a scaffold protein that interacts with many other components of BER as DNA polymerase β, APE1, hOGG1, poly-(ADP-ribose) polymerase, and DNA ligase III in the NH2-terminal, central, and COOH-terminal regions, respectively, as summarized in Figure 1 (Vidal et al., 2001; Marsin et al., 2003; Audebert et al., 2004; Brem and Hall, 2005). Eight common single-nucleotide polymorphisms (SNPs) leading to an amino acid substitution have been described in XRCC1 (Ruttan and Glickman, 2002); three of which are located in or near important protein domains (Shen et al., 1998) (Fig. 1).
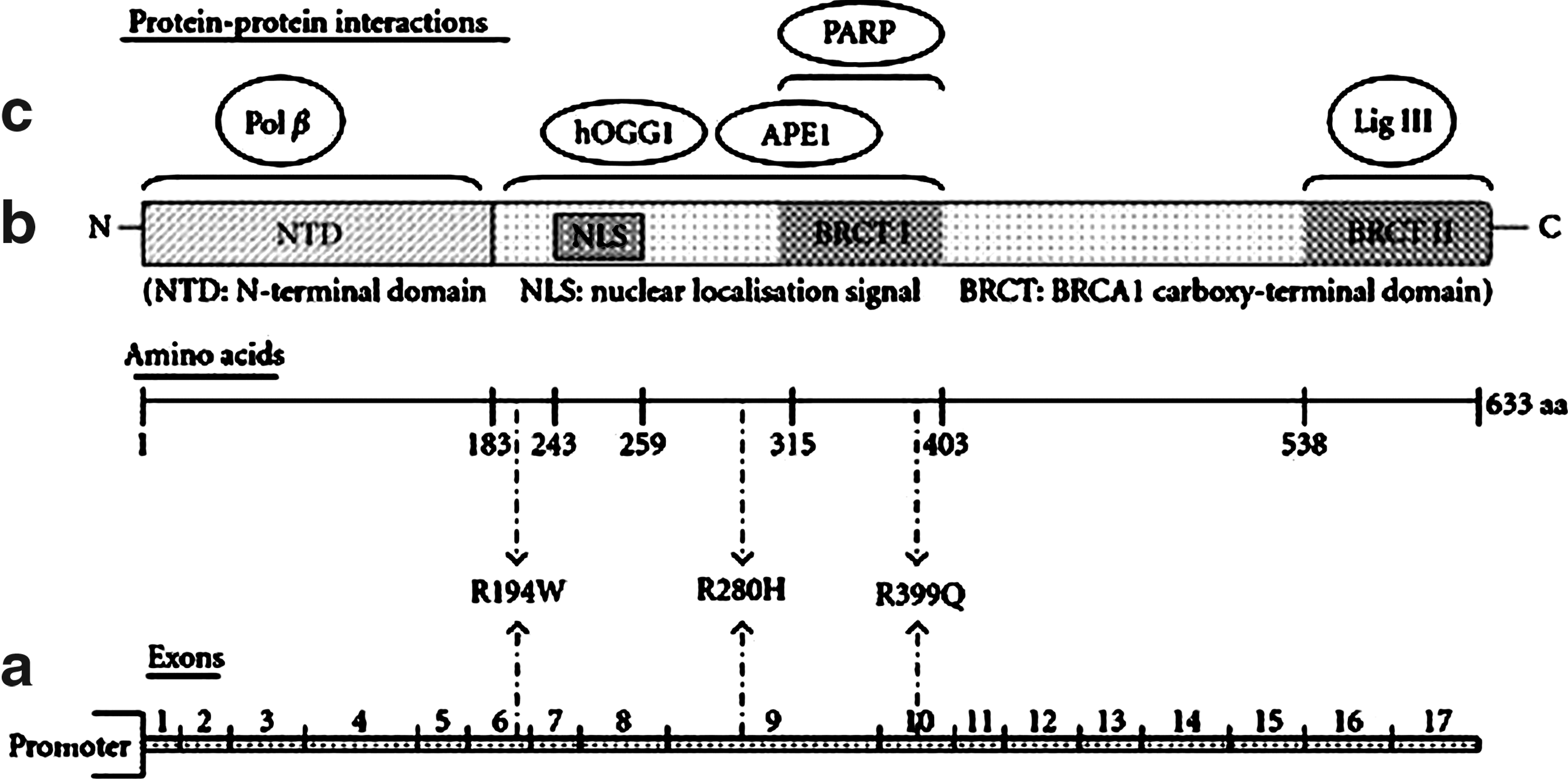
Linear schematic of human XRCC1 gene and protein structure.
The C>T (rs1799782) polymorphism causing arginine to tryptophane substitution at codon 194 of exon 6 (Arg194Trp) occurs near the N-terminal domain. The 194Trp variant allele has been associated with a decrease in cancer risk (Goode et al., 2002; Hung et al., 2005a, 2005b). This unexpected protective effect has been confirmed by experimental studies (Wang et al., 2003; Takanami et al., 2005).
The G>A (rs25489) polymorphism causing arginine to histidine substitution at codon 280 of exon 9 (Arg280His) occurs near the BRCT1 domain. It has been reported that the 280His variant allele was associated with a slight increase in cancer risk (Hu et al., 2005). However, an approximate 3.5-fold increase in skin cancer risk has been, recently, reported in meta-analysis study (Zhang et al., 2011).
The G>A (rs25487) polymorphism causing arginine to glutamine substitution at codon 399 of exon 10 (Arg399Gln) occurs at a conserved residue in the poly (ADP-ribose) polymerase-binding domain of XRCC1. The Arg399Gln polymorphism has been extensively investigated. A positive association between the 399Gln variant allele and cancer risk has been reported in several studies (Divine et al., 2001; Huang et al., 2011; Samulak et al., 2011; Wu et al., 2011; Zhou et al., 2011; Yin et al., 2012). However, no overall association has been seen by others (Smith et al., 2003a, 2003b; Wang et al., 2009; Xue et al., 2011).
The variant allele frequencies of Arg194Trp, Arg280His, and Arg399Gln polymorphisms in the human population are 0.131, 0.061, and 0.266, respectively, according to the NCBI dbSNP (www.ncbi.nlm.nih.gov/projects/SNP/) database. However, a difference in their prevalence has been reported among ethnic groups as well as within the same ethnic group, according to the HapMap(www.hapmap.org) project database. To the best of our knowledge, there is no study in this regard in the Tunisian population (TUN) and, therefore, further investigation is merited to establish our genotype profile.
Hence, our present study aimed to determine the genotype and the allele frequencies of XRCC1 polymorphisms Arg194Trp, Arg280His, and Arg399Gln in the healthy TUN, and to compare them with those reported in the HapMap (www.hapmap.org) populations from other ethnic groups. In addition, we predicted their eventual functional effect based on the protein conservation analysis by SIFT (http://sift.jcvi.org/www/SIFT_dbSNP.html) software, in order to distinguish between neutral and deleterious amino acid substitutions for further clinical and genetic studies.
Materials and Methods
Studied population and DNA extraction
In the present study, 154 healthy unrelated individuals (56 women and 98 men) from Tunisia were enrolled after approval from the ethics committee of the institute. Their age ranged from 20 to 60 years, with a mean±SD of 37.5±11. Information about social habits and health problems of each individual were gathered through standardized questionnaire. We excluded those who had a history of cancer. Blood samples (5 mL) were collected after getting proper written informed consent, and processed for DNA extraction using phenol-chloroform standard procedure (Lewin and Stewart-Haynes, 1992).
Analysis of XRCC1 polymorphisms
The analysis of XRCC1 polymorphisms was carried out with polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) using primers F: 5′-GCCCCGTCCCAGGTA-3′ and R: 5′-AGCCCCAAGCCCTTTCACT-3′ for exon 6 (491 bp) (Ruth et al., 1999); F: 5′-TGGGGCCTGGATTGCTGGGTCTG-3′ and R: 5′-CAGCACCACTACCACACCCTGGCA-3′ for exon 9 (280 bp) (Tuimala et al., 2002); and F: 5′-GCCCCTCAGATCACACCTAC-3′ and R: 5′-CATTGCCCAGCACAGGATAA-3′ for exon 10 (214 bp) (Vodicka et al. 2004).
These fragments were amplified separately under the following conditions: a reaction mixture (50 μL) contained approximately 50 ng of genomic DNA, 10 μM of each primer, 2 mM MgCl2, 10 mM mix dNTP, 1×PCR buffer, and 1.5 U Taq DNA polymerase. The mixtures were amplified with a GeneAmp PCR System 9700 Thermal cycler (Applied Biosystems). The PCR profile consisted of an initial melting step of 95°C for 5 min, followed by 35 cycles with melting at 94°C for 50 s, annealing at 62°C (exon 6) for 60 s, 71.5°C (exon 9) for 50 s, and 61°C (exon 10) for 30 s, and elongation at 72°C for 50 s with a final elongation step of 72°C for 10 min. The PCR products were checked on a 2% agarose gel and photographed using Gel doc and were then subjected to RFLP analysis.
The restriction enzyme MspI (Fermentas) was used to distinguish the codon 194 C→T polymorphism of exon 6, in which a loss of MspI restriction site occurs in the polymorphic allele. PCR-RFLP patterns resulted in three bands of 178, 292, and 21 bp in the homozygous wild type (Arg/Arg), whereas in the heterozygous mutant type (Arg/Trp), four bands at 313, 292, 178, and 21 bp were produced. The same restriction enzyme MspI was used to distinguish the codon 399 G→A polymorphism at exon 10, in which a created MspI restriction site occurs in the polymorphic allele. The homozygous wild type (Arg/Arg) has the PCR product band 214 bp, whereas the homozygous mutant type (Gln/Gln) has two bands of 160 and 54 bp. The restriction enzyme RsaI (Fermentas) was used to distinguish the codon 280 G→A polymorphism of exon 9, in which a created RsaI restriction site occurs in the polymorphic allele. The homozygous wild type (Arg/Arg) has the PCR product band (280 bp), and the homozygous mutant type (His/His) resulted in two fragments of 140 and 140 bp.
Digestion of the PCR products was carried out using 10 U of MspI or RsaI and the 10×buffer supplied with each restriction enzyme at 37°C overnight. The digestion products were separated on a 2.5% and 3.5% agarose gel and photographed using Gel doc film (Bio-Rad).
Bioinformatics analysis
The genotype and the allele frequencies of XRCC1 polymorphisms (Arg194Trp, Arg280His, and Arg399Gln) in the other ethnic groups were obtained from the HapMap (www.hapmap.org) database. The pairwise linkage disequilibrium (LD) (r2) was calculated using the Haploview software (version 4.2) and HapMap (www.hapmap.org) data. The possible haplotypes in our population and their estimated frequencies were performed using PHASE software (version 2.1).
Evolutionary protein conservation was examined by SIFT (http://sift.jcvi.org/www/SIFT_dbSNP.html) software (version 1.03) developed by Ng and Henikoff (2001, 2002). SIFT (http://sift.jcvi.org/www/SIFT_dbSNP.html) predicts whether an amino acid substitution affects protein function. SIFT (http://sift.jcvi.org/www/SIFT_dbSNP.html) prediction is based on the degree of conservation of amino acid residues in sequence alignments derived from closely related sequences, collected through PSI-BLAST. The median sequence conservation ranges from 0 to 4.32; ideally the number would be between 2.75 and 3.5, the default median sequence conservation in the range of 3.0. The amino acid substitution is predicted damaging if the SIFT score is ≤0.05, and tolerated if the score is >0.05 (The SIFT score ranges from 0 to 1.).
Statistical analysis
The allele frequency was calculated from the genotype frequency. Hardy-Weinberg equilibrium was examined for all SNPs, using a Chi-square (χ2) test with one degree of freedom. The 95% confidence interval (CI) of the genotype frequencies was calculated based on a binomial distribution. The comparison of genotype frequency distributions as a gender function was performed by χ2 test. The difference between TUN and HapMap populations was examined by χ2 test, using the allele frequencies, and a p-value<0.05 was considered statistically significant. These calculations were conducted by the computer SPSS program (Version 17.0).
Results and Discussion
The discordance in genetic variation observed between ethnicities highlights the necessity for researchers to establish a specific genotype profile for each population, in order to prioritize substitutions for further studies. In the present one, we studied the major three SNPs in XRCC1 gene as Arg194Trp (C>T, rs1799782), Arg280His (G>A, rs25489), and Arg399Gln (G>A, rs25487).
The genotype and the allele frequencies of XRCC1 polymorphisms in our population are summarized in Table 1. The genotype distributions showed no statistical difference as a gender function (p>0.05; Table 1). The observed genotype frequencies are consistent with Hardy-Weinberg expectations (p>0.05; Table 1).
n, Number of subjects.
Genotype frequencies.
There was no deviation from the expected Hardy-Weinberg frequencies (p>0.05).
p>0.05, no difference as a function of gender.
Genotype and allele frequencies of XRCC1 (C>T) Arg194Trp
The observed CC, CT, and TT genotype frequencies were 0.863, 0.137, and 0.000, respectively (Table 1). The HapMap data for this SNP were only available for the ethnic groups, Caucasian Europeans (CEU), Asians (HCB and JPT), and Sub-Saharan Africans (YRI). The distribution of genotype frequencies with 95% of CI for Tunisians (TUN) and the other ethnic groups is shown graphically in Figure 2. The wild-type (C) and variant allele (T) frequencies for Tunisians (TUN) and the other HapMap groups are shown in Table 2. The prevalence of Arg194Trp polymorphism in the Tunisian population (TUN) is consistent with those reported in both Caucasian (CEU) and Sub-Saharan African (Negroid YRI) populations (p>0.05; Table 2). However, a clear discordance was observed compared with those observed in Asian populations (JPT and HCB) (p<0.05; Table 2).
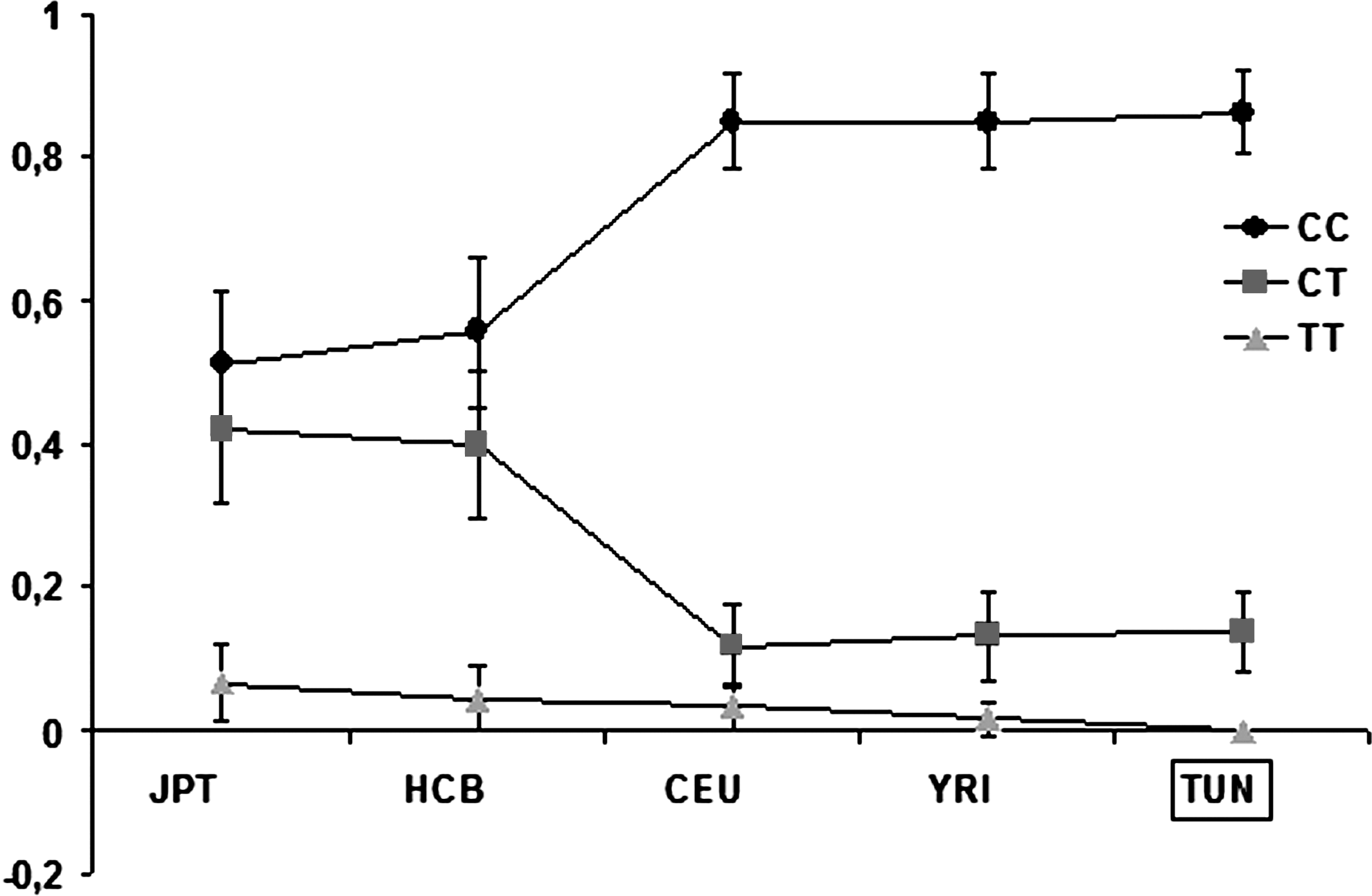
Genotype frequencies of C>T (Arg194Trp) polymorphism in Tunisian (TUN) and HapMap populations (JPT, HCB, CEU, and YRI) (www.hapmap.org). Circle: frequency of homozygous wild-type (CC); square: frequency of heterozygous (CT); and triangle: frequency of homozygous mutant type (TT); bars are 95% confidence intervals.
n, Number of individuals.
Population data source: (www.hapmap.org).
CEU, Utah residents with Northern and Western European ancestry from CEPH collection; HCB, Han Chinese in Beijing, China; JPT, Japanese in Tokyo, Japan; YRI, Yoruban in Ibadan, Nigeria.
p<0.05, a significant difference between TUN and other populations.
p>0.05, no significant difference between TUN and other populations.
Genotype and allele frequencies of XRCC1 (G>A) Arg280His
The observed GG, GA, and AA genotype frequencies were 0.643, 0.305, and 0.052, respectively (Table 1). The distribution of genotype frequencies with 95% of CI for Tunisians (TUN) and the other ethnic groups is shown graphically in Figure 3. The wild-type (G) and variant allele (A) frequencies for Tunisians (TUN) and the other HapMap groups are shown in Table 2. The Tunisian population appears to have a significantly higher prevalence compared with all other ethnic groups included in our study, such as Caucasians (CEU and TSI), Asians (HCB, CHD, GIH, and JPT), Africans Sub-Saharan (LWK, MKK, and YRI), American-Africans (ASW), and American-Mexicans (MEX) (p<0.05; Table 3).
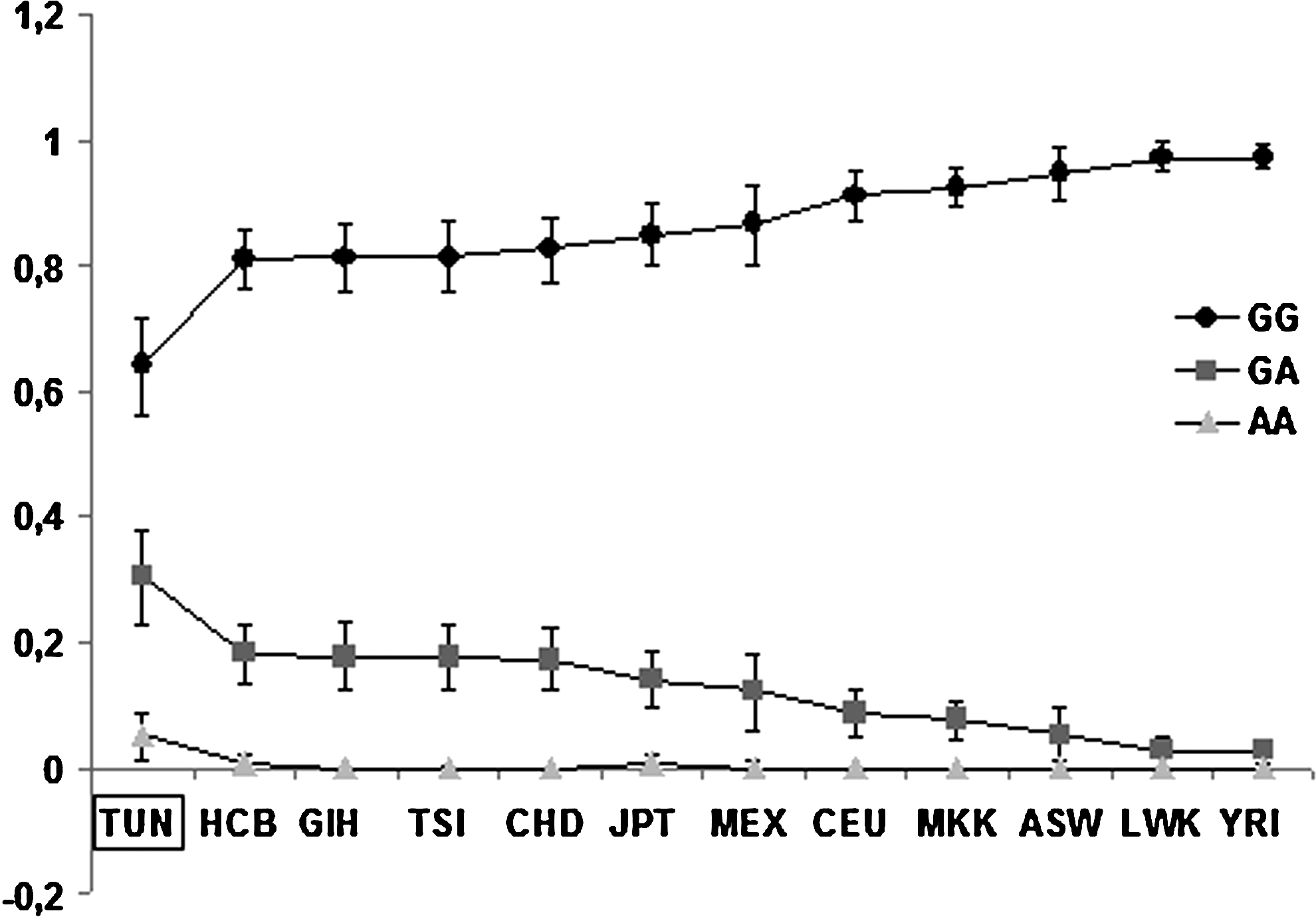
Genotype frequencies of G>A (Arg280His) polymorphism in Tunisian (TUN) and HapMap populations (HCB, GIH, TSI, CHD, JPT, MEX, CEU, MKK, ASW, LWK, and YRI) (www.hapmap.org). Circle: frequency of homozygous wild-type (GG); square: frequency of heterozygous (GA); and triangle: frequency of homozygous mutant type (AA); bars are 95% confidence intervals.
n, Number of individuals.
Population data source: (www.hapmap.org).
CEU, Utah residents with Northern and Western European ancestry from the CEPH collection; TSI, Tuscan in Italy; HCB, Han Chinese in Beijing, China; CHD, Chinese in Metropolitan Denver, Colorado; GIH, Gujarati Indians in Houston, Texas; JPT, Japanese in Tokyo, Japan; LWK, Luhya in Webuye, Kenya; MKK, Maasai in Kinyawa, Kenya; YRI, Yoruban in Ibadan, Nigeria; MEX, Mexican ancestry in Los Angeles, California; ASW, African ancestry in Southwest United States.
p<0.05, significant difference between TUN and other populations.
p>0.05, no significant difference between TUN and other populations.
This unexpected data could be explained by the high rate of inbreeding in the Tunisian population being the most important effect. It has been reported that the unions between Tunisian relatives represent more than 32%, which may reach 60% in rural areas (Riou et al., 1989; Ben Arab et al., 2004; Kerkeni et al., 2007). This endogamy creates groups in the population where chances that two gametes carrying the same genetic information meet each other are more elevated than in the general population. Therefore, endogamy has direct consequences on distribution, structure, heterogeneity, and homozygosity tendency in Tunisian population (Ben Mrad and Chalbi, 2004).
Genotype and allele frequencies of XRCC1 (G>A) Arg399Gln
The observed GG, GA, and AA genotype frequencies were 0.416, 0.448, and 0.136, respectively (Table 1). The distribution of genotype frequencies with 95% of CI for Tunisians (TUN) and the other ethnic groups is shown graphically in Figure 4. The wild-type (G) and variant allele (A) frequencies for Tunisians (TUN) and the other HapMap groups are shown in Table 2. Unlike 194 and 280 polymorphisms, the Arg399Gln polymorphism showed a relatively high prevalence in Tunisian population and in all HapMap groups. Compared with other groups, a similarity was seen between Tunisian populations and either Caucasian (CEU and TSI), Asian (GIH, HCB, CHD, and JPT), and American-Mexican (MEX) populations (p>0.05; Table 2). However, a significant difference was shown compared with American-Africans (ASW) and African Sub-Saharans (LWK and MKK and YRI) (p<0.05; Table 2).

Genotype frequencies of G>A (Arg399Gln) polymorphism in Tunisian (TUN) and HapMap populations (GIH, CEU, TSI, MEX, CHD, JPT, HCB, MKK, ASW, YRI, and LWK) (www.hapmap.org). Circle: frequency of homozygous wild-type (GG); square: frequency of heterozygous (GA); triangle: frequency of homozygous mutant type (AA); bars are 95% confidence intervals.
LD and haplotype analysis
Calculations of pairwise LD showed an equilibrium status for XRCC1 polymorphisms; r2 was 0, 0.01, and 0.01 for (rs25489/rs25487), (rs1799782/rs25489), and (rs1799782/rs25487), respectively. Haplotype distributions are shown in Table 4. Four out of 8 possible haplotypes were observed in our population with frequencies greater than 5%. The most frequent haplotype was GGC (0.455±0.009) that is composed of three wild-type alleles (Table 4), followed by GAT haplotype (0.279±0.008) that contained the variant alleles of G>A (rs25489) and C>T (rs1799782). To elucidate the influence of XRCC1 haplotypes on cancer risk, Saadat evaluated whether the Arg194Trp and Arg399Gln haplotypes contribute to breast cancer susceptibility among Caucasoid subjects (Saadat, 2010). Saadat reported a borderline increased risk of breast cancer associated with the Arg194-Gln399 haplotype versus the Arg194-Arg399 haplotype. However, a higher association has been observed between head and neck cancer and XRCC1 haplotypes, especially with Arg194Trp-Arg399Arg haplotype (Kowalski et al., 2009). To the best of our knowledge, there is no study pertaining to the XRCC1 haplotypes in our population; therefore, these data would be useful in epidemiologic and experimental studies. However, largest population studies might be more suitable to assess the effect of each haplotype.
The major haplotypes are in bold (frequency>5%).
Effect of Arg194Trp substitution on protein function
The arginine (R) to tryptophane (W) substitution at 194 codon was predicted as damaging with a SIFT score of 0.00 (Table 5). In our analysis, the SIFT prediction was based on 98 homologous proteins in the alignments. The Arg194Trp substitution occurs near the N-terminal domain of the XRCC1 gene (Shen et al., 1998; Marintchev et al., 1999), within possible binding domains of several interactive protein partners, including APE1, polymerase β, hOGG1, and PARP, as described in Figure 1 (Ladiges, 2006). This area is rich in proline, serine, arginine, and lysine residues, and, therefore, the transition from positively charged arginine to a hydrophobic tryptophan may affect binding and DNA repair efficiency (Ladiges, 2006).
Amino acid.
SIFT score: The amino acid substitution is predicted DAMAGING if the SIFT score ≤0.05, and TOLERATED if SIFT score >0.05.
Median Info: This is used to measure the diversity of the sequences used for prediction; it ranges from 0 to 4.32; ideally the number would be between 2.75 and 3.5.
The number of sequences at position: The number of homologue sequences that have an amino acid at the position prediction (SIFT automatically chooses the sequences).
Effect of Arg280His substitution on protein function
The arginine (R) to histidine (H) substitution at 280 codon was predicted as damaging with a SIFT score of 0.05 (Table 5). The Arg280His substitution occurs near the BRCT1 domain (breast cancer susceptibility protein-1 domain) of the XRCC1 gene (Fig. 1) (Shen et al., 1998), and lies in between the DNA polymerase β and PARP binding areas (Thompson and West, 2000). According to Caldecott (2003), the transition from a positively charged arginine to histidine within the conserved region could potentially alter XRCC1 function and its ability to interact with the apurinic/apyrimidinque endonuclease (APE)-binding domain. All these findings highlight the deleterious effect of Arg280His polymorphism on XRCC1 function. In this regard, the 280His variant allele has been suggested to confer increased mutagen sensitivity (Tuimala et al., 2002).
Effect of Arg399Gln substitution on protein function
The arginine (R) to glutamine (Q) substitution at 399 codon was predicted as damaging with a SIFT score of 0.05 (Table 5). This substitution occurs within the breast cancer C-terminal BRCT domain at a conserved residue in the poly (ADP-ribose) polymerase PARP binding domain (Zhang et al., 1998; Ladiges, 2006). Taylor et al. (2002) showed that although BRCT1 domain is critical for efficient single-strand break repair and cell survival, Arg399Gln polymorphism located within this domain did not appear to significantly affect XRCC1 function. On the contrary, by using molecular dynamics techniques, Monaco et al. (2007) predicted the structure of wild-type and polymorphic form of BRCT1 domain of XRCC1 demonstrating that the polymorphism in exon 10 changed the XRCC1's secondary structure (Taylor et al., 2002; Monaco et al., 2007). Therefore, any change in the BRCT structure has the potential to lead to defects in the detection of the DNA damage and hence the activation of the BER pathway. In fact, several reports suggested that 399 polymorphism may result in deficient DNA repair and individuals with Gln variant allele exhibited higher level of DNA damage than those with homozygous wild-type (Arg/Arg) and heterozygous (Arg/Gln) genotypes (Lunn et al., 1999; Abdel-Rahman and El-Zein, 2000; Miller et al., 2001; Matullo et al., 2001).
In the overall, our findings revealed an interethnic variability in prevalence of XRCC1 polymorphisms. Tunisia with its high levels of endogamy could provide a unique resource for dissecting complex disease etiology. Our findings lead us to suggest that the Tunisian population could be more varied in the capacity of DNA repair than other populations in the world, and therefore the susceptibility to disease incidences due to environmental pollutants will vary; thus, extrapolated results from other populations cannot be applied on the Tunisian population. The present study establishes the prevalence and the possible effects of XRCC1 polymorphisms. This data will provide a basic database for the future clinical and genetic studies pertaining to variability and the defect of DNA repair capacity.
Footnotes
Acknowledgments
The authors would like to thank the volunteers for their cooperation. We were also grateful to Pr. Jamil JAOUA, Founder and former head of the English Department at the Sfax Faculty of Science, for proofreading this article. This work was supported by the Ministry of Higher Education and Scientific Research in Tunisia and Chemical Tunisian Group.
Author Disclosure Statement
The authors declare that they have no actual or potential conflict of interest, including any financial, personal, or other relationships, with other people or organizations concerning this work.