
Abstract
Background: The distribution of polymorphisms in the CYP2D6 and CYP2C19 genes allows inferring the potential risk for specific adverse drug reactions and lack of therapeutic effects in humans. This variability shows differences among human populations. The aim of this study was to analyze single-nucleotide polymorphisms related to a poor metabolizer (PM) phenotype in nonpreviously studied Amerindian groups and Mestizos (general admixed population) from Mexico. Methods: We detected by SNaPshot® different polymorphisms located in CYP2D6 (*3, *4, *6, *7, and *8) and CYP2C19 (*2, *3, *4 and *5) in western Mestizos (n=145) and five Amerindian groups from Mexico: Tarahumaras from the North (n=88); Purépechas from the Center (n=101); and Tojolabales (n=68), Tzotziles (n=88), and Tzeltales (n=20) from the Southeast. Genotypes were observed by capillary electrophoresis. The genetic relationships among these populations were estimated based on these genes. Results and Discussion: The wild-type allele (*1) of both genes was predominant in the Mexican populations studied. The most widely observed alleles were CYP2C19*2 (range, 0%-31%) and CYP2D6*4 (range, 1.2%-7.3%), whereas CYP2D6*3 was exclusively detected in Mestizos. Conversely, CYP2C19*4 and *5, as well as CYP2D6*3, *6, *7, and *8, were not observed in the majority of the Mexican populations. The Tarahumaras presented a high frequency of the allele CYP2C19*2 (31%) and of homozygotes *2/*2 (10.7%), which represent a high frequency of potentially PM phenotypes in this Amerindian group. The genetic distances showed high differentiation of Tarahumaras (principally for CYP2C19 gene). In general, a relative proximity was observed between most of the Amerindian, Mexican-Mestizo, and Latin-American populations. Conclusion: In general, the wild-type allele (*1) predominates in Mexican populations, outlining a relatively homogeneous distribution for CYP2C19 and CYP2D6. The exception is the Tarahumara group that displays a potentially increased risk for adverse reactions to CYP2C19-metabolized drugs.
Introduction
T
The gene CYP2C19 codifies for the enzyme mephenytoin 4-prime-hydroxylase, which is of clinical importance for the metabolism of the anticonvulsant mephenytoin, antiulcer drugs (such as omeprazole), antidepressants, the antimalarial drug proguanil, the muscle relaxant carisoprodol, the antiplatelet clopidrogel, cyclophosphamide, and thalidomide, among others (Takada et al., 2004; Hulot et al., 2006; Li et al., 2007). Twenty eight allelic variants have been described for CYP2C19. The alleles CYP2C19*2 through CYP2C19*8 have been reported to be without enzyme activity and CYP2C19*9 through CYP2C19*11 showed decreased activity in vitro. Finally, CYP2C19*12 exhibits unstable activity in vitro, whereas CYP2C19*17 demonstrates increased transcriptional activity (Sibbing et al., 2010). The remaining alleles also exhibit the absence of enzyme activity (www.cypalleles.ki.se/cyp2c19.htm). Metabolizer phenotypes generated due to the variable enzyme activity of CYP2D6 and CYP2C19 include Ultrarapid (UM), Extensive (EM), Intermediate (IM), and Poor metabolizers (PM). The allele frequencies of these genes show differences among populations. For example, alleles CYP2D6*3, *4, *5, and *6 are responsible for 97% of PM in Caucasians, with CYP2D6*3 and *4 the most common alleles (Sachse et al., 1997), whereas in China, these alleles are rare and explain only ∼1% of PM phenotypes (Wang et al., 1993). For CYP2C19, CYP2C19*2 and *3 are the most common alleles and are those mainly responsible for the PM phenotype in Caucasian and Asian populations, respectively (Xie et al., 2001; Mizutani, 2003). PM frequency is 5% in Caucasian and African populations and ∼20% in Asians.
In Mexico, there are two main populations: Native groups (Amerindians) and Mestizos the result of post-Columbian admixture. Mexican Amerindians constitute >68 Native groups, >85 languages and variant dialects, and are geographically located mainly in the Center and Southeast of the country (Martinez-Cortes et al., 2010). Conversely, Mexican-Mestizos are widely distributed throughout the country, comprise ∼93% of the total population, and arose during and after European contact with the New World by admixture among Spaniards, Amerindians, and, to a lesser degree, with African slaves (Rubi-Castellanos et al., 2009). Presently, the distribution of alleles related with the PM phenotype for CYP2D6 and CYP2C19 is poorly known in both Mexican-Mestizos and Amerindians. CYP2D6 alleles CYP2D6*2, *3, *4, *5, *10, and *17 in Mestizos from Mexico City display a frequency of 19.3%, 1.4%, 11.2%, 2.6%, 12.4%, and 1.6%, respectively (Lopez et al., 2005). In Mestizos from Durango (Northwest), alleles CYP2D6*2, *3, *4, *5, and *10 were reported, with frequencies of 10.7%, 0.9%, 13.1%, 1.3%, and 2.3%, respectively, in addition to 6.8% of PM phenotypes, similar to those reported in Caucasians (10%) (Sosa-Macias et al., 2006; Sosa-Macias et al., 2010). In Mexican-Amerindian groups, this information is more limited; during the search for CYP2D6*3, *4, *6, and *10 in Tepehuanos from Durango (Northwest, Mexico), only CYP2D6*4 was found at a low frequency (0.6%), suggesting that the PM phenotype is absent in this Amerindian group (Sosa-Macias et al., 2006). In addition, CYP2D6*4 (0.21%) and CYP2D6*10 (0.05%) were also detected in Mennonites of Caucasian origin residing in Durango, Mexico, whereas CYP2D6*6 was not found (Alanis-Bañuelos et al., 2007). In Mexican-Americans, frequencies of alleles CYP2D6*4 and *6 were 10 and 0.4%, respectively (Luo et al., 2005). Recently, CYP2D6 resequencing data from two Mexican-Mestizo populations allowed detection of 14 novel variants, including allele CYP2D6*82 that was hypothesized as Amerindian because of its identification in three Mexican-Amerindian groups (Contreras et al., 2011). In this study, the frequencies of CYP2D6*1, *2, *4, *5, *10, *29, *53, *82, and its duplications were 50.0, 25.5, 14.1, 2.0, 2.6, 1.0, 0.5, 2.1, and 3.6%, respectively.
For CYP2C19, the first pharmacokinetic study was conducted in western Mexican-Mestizos, obtaining a frequency of 6% and 4% for PM and UM, respectively (González et al., 2003). Afterward, in a case-control study carried out in the state of Nuevo León (Northeast Mexico), estimated PM frequency of CYP2C19 was 8% in the general population and 11.9% in cases (Garza-González et al., 2007). Recently, in one gene-disease association study, the estimated frequency of CYP2C19*1 and *2 was 91.6% and 8.4% in healthy volunteers from Mexico City, respectively, whereas CYP2C19*3 was not found (Hoyo-Vadillo et al., 2010). To our knowledge, in the unique study of CYP2C19 in Amerindian groups, PM individuals were not found in the Cuna Amerindian group from Panama (Inaba et al., 1988). These studies in Native Americans are interesting, taking into account their particular lifestyle, customs, and geographic location, because these are related with the clinical impact of these alleles in terms of conferring risk for or susceptibility to adverse drug reactions. Because of the relationship between a genotype and a phenotype, at least to some extent, these analyses help to define the prevalence of different drug phenotype metabolizers in populations (Goldstein, 2001; Desta et al., 2002). The aim of this work was to define CYP2D6 and CYP2C19 allele frequency in five Native groups from different regions (North, West, and Southwest) of Mexico, in addition to Mestizos from western Mexico. We found a high frequency of the wild-type allele in the Mexican populations, but an elevated frequency of homozygous CYP2C19 *2/*2 in the Tarahumara group (North-Central Mexico) that could represent a higher risk for adverse drug reactions for CYP2C19-metabolized drugs.
Materials and Methods
DNA samples
A total of 416 and 506 unrelated individuals from five Amerindian groups and western Mestizos from Mexico were analyzed for CYP2D6 and CYP2C19, respectively. Difference in population sample sizes was due to the scarce quantity and bad quality (degradation) of some DNA samples. DNA was extracted from peripheral blood samples by the salting-out method (Miller et al., 1988) or with the Qiagen® FlexiGene DNA kit. The characteristics of the population samples analyzed here and those utilized for comparison purposes are described in Table 1. Written informed consent was obtained from all volunteers. The experimental protocol was approved by the Committee of Ethics in Research of the Centro Universitario de la Ciénega, Universidad de Guadalajara (CUCI-UdeG).
Genotyping
Five CYP2D6 alleles (CYP2D6*3, *4, *6, *7, and *8) and four CYP2C19 alleles (CYP2C19*2, *3, *4, and *5) were analyzed by SNapShot™ with previously described primers and conditions (Bender, 2005). However, the poor reproducibility of the Long PCR protocol to amplify the CYP2D6 fragment (5.1 kb) compelled us to employ an alternative PCR multiplex assay with the following primers sets previously reported (Gersdorff, 2005): (1) forward 5′-GGCTGGCAAGGTCCTACGC-3′ and reverse 5′-CATTCCTCCTGGGACGCTCA-3′ for CYP2D6*3 (196 bp); (2) forward 5′-GCCTGGGCAAGAAGTCGCT-3′ and reverse 5′-AGGGAGGCGATCACGTTGCT-3′ for CYP2D6*4/*6/*8 (227 bp); and 3) forward 5′-CGTTCTGTCCCGAGTATGCT-3’ and reverse 5’-GCCCTATCACGTCGTCGAT-3′ for CYP2D6*7 (420 bp). To validate SNaPshot assays, genotypes in 10 individuals were confirmed with commercially available qPCR assays using TaqMan probes for the most common variant allele CYP2D6*4 (Applied Biosystems).
PCR multiplexes were performed with the Qiagen Multiplex PCR kit (Qiagen) and with 10 ng of genomic DNA in a 10 μL of total volume. PCR products were observed by 6% (29:1) polyacrylamide gel electrophoresis followed by silver staining. After amplification, 0.2 μL of PCR product was purified with 1 μL of ExoSAP-IT (Amersham, Biosciences). The mix was incubated at 37°C for 15 min followed by 15 min at 80°C to inactive the enzyme. The minisequencing reaction was conducted in a total volume of 5 μL with 1 μL of purified PCR product, 2.5 μL of SNaPshot reaction mix (Applied Biosystems), and 0.7 μL of single base extension (SBE) primer mix. SBE primers and cycling conditions were those specified by Bender (2005). Five μL of the SBE product were treated with 1 μL of the SAP enzyme (Amersham Biosciences) for 60 min at 37°C, followed by 15 min at 80°C for enzyme inactivation. SNaPshot products were observed in the ABI Prism 3130® Genetic Analyzer (Applied Biosystems). One microliter of SBE product was mixed with 9 μL of HiDi™ formamide and 0.3 μL of size standard LIZ-120 (Applied Biosystems); these were injected for 16 s at 1.2 kV and 5 μA at 60°C in a 36-cm-long capillary filled with POP7®. CYP2D6 and CYP2C19 genotypes were interpreted with GeneMapper® v3.2 (Applied Biosystems) software. The presence of the wild-type allele (CYP2D6*1 and CYP2C19*1) was inferred when derived states were absent in all single-nucleotide polymorphisms (SNP) detected.
Statistical analysis
Allele and genotype frequencies were estimated by the gene counting method. Genotype distribution agreement with Hardy-Weinberg expectations was checked by exact tests. Genetic relatedness between populations based on CYP2D6 and CYP2C19 was evaluated by pairwise comparisons (exact tests) and genetic distances (FST), which additionally were represented in a multidimensional scaling plot (MDS) with SPSS 19.0.0 for Windows software. The Arlequin 2000 program was employed for genetic data analyses (Excoffier et al., 2005).
Results and Discussion
To our knowledge, this is the first report of CYP2C19 and CYP2D6 SNPs related to PM phenotypes in the Mexican populations here analyzed. Although a low allele diversity was observed in the majority of populations, some limitations must be considered regarding these findings: (1) European or Asian prevalence of the alleles identified, which means that alleles of Amerindian origin are not represented, such as the variant CYP2D6*82 recently described (Contreras et al., 2011). (2) Deletions related with the PM phenotype were not included, particularly CYP2D6*5, which has been described with a low frequency (0.5%) in Tepehuanos, one Amerindian group from Northwest, Mexico (Sosa-Macias et al., 2010). (3) Alleles related to the UM phenotype were not included in this study, such as CYP2C19*17 (Sim et al., 2006) and CYP2D6 duplications. (4) The small population sample of some Amerindian groups, such as Tzeltales (n=19), which is explained by the difficulty to obtain samples from these isolated communities. These limitations should be considered to design additional studies in Mexican populations, principally Amerindian groups, in the near future.
Allele frequencies
In general, low allele diversity was observed due to the predominance of the wild-type allele for both genes (>92%) (Table 2). The exception was the Tarahumara group that exhibited a lower CYP2C19*1 frequency (69%). The alleles observed in the majority of Mexican populations were CYP2C19*2 (range, 0%-6.9%) and CYP2D6*4 (range, 1.2%-7.3%). Conversely, alleles CYP2D6*6, *7, and *8, as well as CYP2C19*3, *4, and *5, were not observed in the majority of Mexican population samples. It was particularly noteworthy that CYP2D6*3 was observed exclusively in Mestizos, and that CYP2C19*2 had an elevated frequency in Tarahumaras (31%), but was absent in Tzeltales, who solely presented the wild-type allele CYP2C19*1 (Table 2).
These frequencies in Mexican populations are similar to those reported in European populations in which CYP2C19*2 and CYP2D6*4 are the modal alleles (Sachse et al., 1997; Mizutani, 2003). In particular, CYP2C19*2 frequency in Mestizos (6.9%) is similar to that reported in Mexico City (8.4%) (Hoyo-Vadillo et al., 2010) and Mexican-Americans (9.7%) (Luo et al., 2006). Regarding the high frequency of CYP2C19*2 in Tarahumaras, this could be related with the high differentiation by genetic drift described for this group, in addition to their proximity to non-Amerindian groups, such as the NaDene groups located in North America (Rangel-Villalobos et al., 2000; Rangel-Villalobos et al., 2008). With respect to the allele CYP2D6*4 frequency in Mestizos (5.6%), although this was lower than those reported in Mexico City (11.2%) and Durango (13.1%) (17-18), this difference was not significant (p>0.1268). Finally, all of the SNPs analyzed in these Mexican populations were in Hardy-Weinberg equilibrium (p>0.05) (data not shown).
CYP2C19 genotype frequencies
The scarce presence of a sole additional allele (CYP2C19*2) in the majority of Mexican populations explains the high frequency of wild-type homozygous *1/*1 (range, 48.8%-100%). Therefore, presence of heterozygous *1/*2 (range, 0%-11.3%) and homozygous *2/*2 (range, 0%-1.4%) also was low, except in Tarahumaras (40.5% and 10.7%, respectively). This suggests, in general, limited frequency of PM phenotypes in Mexican populations. Compared with previous reports, the frequency of homozygous *2/*2 in Mexican-Mestizos (1.4%) is similar to those observed in Mestizos of Colombia and Bolivia (Bravo-Villalta et al., 2005; Isaza et al., 2007). However, a previous study also carried out in western Mexican Mestizos, but based on the metabolic ratio (MR log) omeprazole/hydroxy-omeprazole, estimated a frequency of 6% for PM (González et al., 2003). This constitutes a difference with respect to the PM frequency predicted by the genotypes *2/*2 detected in our study (6% vs. 1.4%) (p=0.047±0.0019). Although this difference could be explained by the limited number of SNPs detected here, and to additional factors that influence the CYP2C19 pharmacokinetic phenotype (Shah, 2005; Jacob et al., 2009), the answer to this finding would require further research.
As previously described, the main finding in this work regarding CYP2C19 comprises the high frequency in Tarahumaras of heterozygous *1/*2 (40.5%) and homozygous *2/*2 (10.7%). This result deserves additional pharmacokinetic studies to confirm the possible increment of PM phenotypes in this Native American group that—presumably—would increase the risk for adverse drug reactions. In psychiatry, for instance, the CYP2C19 phenotype affects the pharmacokinetics of antidepressants, including monoamine oxidase inhibitor moclobemide, tricyclic antidepressants amitriptyline and clomipramine, and selective serotonin reuptake inhibitors (SSRI) sertraline and citalopram (Ingelman-Sundberg et al., 2007); in addition to anxiolytics, such as diazepam and clonazepam (Desta et al., 2002) and antiepileptics, such as clobazam (Seo et al., 2008). Another potential effect of the high frequency of PM for CYP2C19 would be a lesser clinical response to prodrugs, such as clopidogrel, thalidomide, and cyclophosmamide (Takada et al., 2004; Hulot et al., 2006; Li et al., 2007). However, the final impact of CYP2C19-based adverse drug reactions in Tarahumaras would be limited because of the rare application of these drugs in this Native group. This is due to the Tarahumaras' natural lifestyle and high physical activity in the mountains and canyons of the Sierra Madre of Chihuahua (North, Mexico), which additionally complicates the presence of public health services (Pintado-Cortina, 2004). Conversely, modification of the Tarahumara lifestyle by their incorporation into urban areas—and into social health services—probably will increase their exposure to environmental factors and to CYP2C19-metabolized drugs (Yampey, 1981; Torres et al., 2003). This should be considered to avoid adverse drug reactions in Tarahumaras currently incorporated into urban centers. In addition, CYP2C19 has been described as a possible risk factor for lung (odds ratio [OR]=3.23), esophageal (OR=3.18), and stomach cancer (OR=2.86) (Tsuneoka et al., 1996; Shi and Chen, 2004). Therefore, research on CYP2C19 genotypes and phenotypes and prevalence of the previously mentioned cancers in Tarahumaras will probably offer interesting results on this topic. This is of particular note considering additional studies that report weak or no association between CYP2C19 alleles and cancer (Brockmoller et al., 1996; Wadelius et al., 1999). On the other hand, CYP2C19 allele variants do not always represent a risk factor. For example, proton pump inhibitors (PPI) in PM achieve better suppression of gastric acid and efficient eradication of Helicobacter pylori (Furuta et al., 2007; Chaudhry et al., 2008). Based on the generally accepted idea that therapeutic efficiency of omeprazole is influenced by the CYP2C19 genotype, some authors have suggested that this also affects the effectiveness of different PPIs. However, the less clear influence of the CYP2C19 genotype over further PPIs, support that this relationship actually depends on the kind of PPI (Chang et al., 1995; Sapone et al., 2003).
CYP2D6 genotype frequencies
The frequency of the wild-type homozygous *1/*1 was even greater for CYP2D6 (range, 85.3%-97.6%) than for CYP2C19. This was a result of the limited presence of additional alleles CYP2D6*4 and CYP2D6*3; the latter allele only observed in Mestizos. Therefore, the low frequency of heterozygous *1/*4 (range, 2.4%-14.7%) and *1/*3 (2.5% in Mestizos) and the absence of compounds heterozygous *3/*4 or homozygous for these alleles suggest a low frequency of PM for CYP2D6 in Mexican populations. Previous reports of CYP2D6 in Mestizos from Mexico City and Durango (Lopez et al., 2005; Sosa-Macias et al., 2006) showed a similar allele distribution to that of the western Mestizos studied here (p=0.1564 and p=0.5268, respectively). This is a result of the relatively similar frequency of the heterozygous *1/*3 (0.41 and 1.13 vs. 2.5%, respectively) and heterozygous *1/*4 (10.7 and 15.9 vs. 11.2%, respectively). The impact of the heterozygotes is difficult to establish because of wide inter-individual variation in enzyme activity for these genotypes (Carcillo et al., 2003). Conversely, the unique antecedent of CYP2D6 in Mexican-Amerindians derives from Tepehuanos (Northwest) (Sosa-Macias et al., 2006), who exhibited a similar allele frequency to that of all five Native groups and the western Mestizos studied here (p>0.1229). However, it was particularly noteworthy that the frequency of genotype *1/*4 (1.2%) in Tepehuanos was the lowest with respect to the five Amerindian groups (range, 2.4%-14.7%).
Genetic relationship
We attempted to describe genetic relationships among Mexican populations based on these two genes, adding certain populations to the analysis (Table 1). For CYP2C19, the similarity was evident among Mestizos, Latin-American, and Amerindian populations (Fig. 1A). The exception was the Tarahumara group that was similar to populations from Brazil and China (Mongolia). These populations formed a cluster (p>0.01) based on their high CYP2C19*2 frequency. As previously mentioned, separation of Tarahumaras could be explained by gene flow received from nearby Native North Americans, such as the NaDene groups, with a particular gene pool regarding Amerindians located geographically from Mesoamerica to South America (Schurr, 2004), for example, the haplogroup C* of the Y-chromosome, which is restricted to Native populations from North America (Zegura et al., 2004). This hypothesis is in agreement with the genetic differentiation described for Tarahumaras by means of autosomal short tandem repeats and Y-chromosome markers (Rangel-Villalobos et al., 2000; Rangel-Villalobos et al., 2008). For CYP2D6, Mexican-Mestizos presented similarity (p>0.01) (data not shown) with the majority of populations due to their intermediate position between Amerindian groups and Latin-American/Spanish populations (Fig. 1B). Interestingly, Amerindian groups were closer to China, but only China displayed a consistent differentiation with Latin-American and Spanish populations for CYP2D6 (p>0.01) (data not shown).
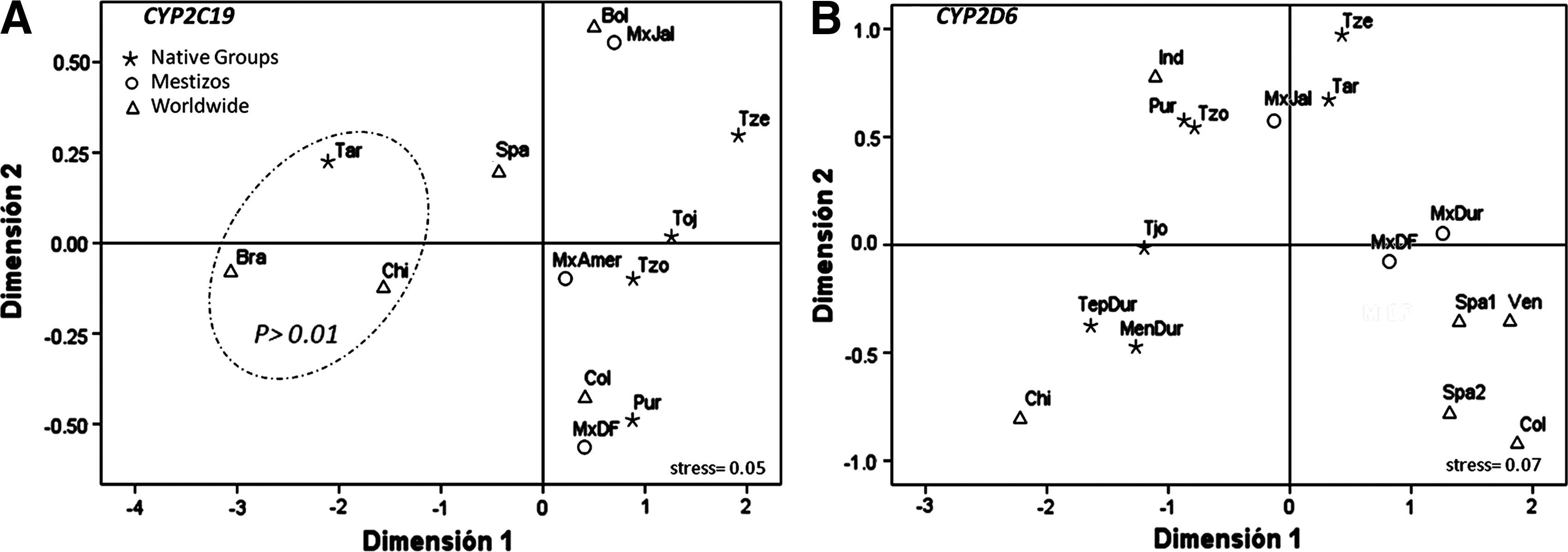
Multidimensional scaling plot that shows genetic distances between Mexican and reference populations based on:
Conclusion
The wild-type allele for PM phenotypes in CYP2D6 and CYP2C19 genes was prevalent in western Mestizos and five Amerindian groups from Mexico. CYP2C19*2 and CYP2D6*4 were widely observed, but CYP2D6*3 was only observed in Mestizos. Tarahumaras were distinguished by their high frequency of CYP2C19*2, which potentially represents a risk factor for this Amerindian group. In general, based on these SNPs, a relative homogeneity was observed among these Mexican populations.
Footnotes
Acknowledgments
We thank the support for this project to the Consejo Estatal de Ciencia y Tecnología de Jalisco (COECyT-JAL; grant 25-3008-32) and Agencia Española de Cooperación Internacional para el Desarrollo (AECID; grant A/017012/08) to H.R.-V. We also thank the Doctoral fellowship of CONACyT to J.S.-F.
Author Disclosure Statement
No competing financial interests exist.