
Abstract
Background:
Many studies have focused on the association between the apolipoprotein A5 (ApoA5) polymorphism and the risk of metabolic syndrome (MetS). However, these studies drew inconsistent conclusions. The aim of this study was to evaluate the exact association between the ApoA5 polymorphism and MetS in a large-scale meta-analysis.
Methods:
The PubMed, Embase, and Science Citation Index (ISI Web of Science) databases were searched to collect all publications on the association between the ApoA5 polymorphism and MetS. Two common variants of ApoA5 (namely −1131T>C in the promoter region and c.56C>G in the coding region) with the risk of MetS were analyzed. The overall odd ratios (ORs) and 95% confidence intervals (CIs) for −1131T>C (CC+TC) versus TT genotype and c.C56G (GG+GC) versus CC were assessed between the MetS and control group. Subgroup analysis was further performed by ethnicity. The meta-analysis was performed by Stata11.0.
Results:
Twelve studies from 10 publications were chosen in our meta-analysis. The combined results showed that C allele carriers (CC+TC) of −1131T>C had a significantly higher risk of MetS for the overall (OR=1.32; 95% CI: 1.14-1.53; p=0.000) with moderate heterogeneity (I2=54.9%, p=0.014). Subgroup analysis was further performed according to ethnicity, and the association was still significant in Asians (OR=1.42; 95% CI: 1.25-1.62; p=0.000), but not in white populations (OR=1.25; 95% CI: 0.97-1.61; p=0.087). When analyzing the association between c.C56G and MetS, the G allele carrier (GG+GC) genotype significantly increased the risk of MetS (OR=1.32; 95% CI: 1.15-1.50; p=0.000) in white populations. No significant publication bias was observed in either −1131T>C or c.C56G.
Conclusions:
Our study suggested that the ApoA5 −1131T>C polymorphism was significantly associated with the risk of MetS in Asians, but not in white populations. However, the c.C56G polymorphism was significantly associated with MetS in white populations.
Introduction
M
Apolipoprotein A5 (ApoA5) is a novel apolipoprotein discovered by two study groups in 2001 (Pennacchio et al., 2001; van der Vliet et al., 2001), and it is located on human chromosome 11q23, proximal to the ApoA1/C3/A4 gene cluster. Function study in ApoA5 knockout and overexpressed mice shows that ApoA5 can significantly reduce plasma triglyceride levels (Pennacchio et al., 2001; van der Vliet et al., 2002). To date, more than 10 polymorphic sites of ApoA5 have been reported (Hubacek, 2005). Among them, −1131T>C in the promoter region and c.56C>G in the coding region are of special interest, for several studies have found these two variants of ApoA5 were closely associated with higher plasma triglyceride levels (Baum et al., 2003; Matsunaga et al., 2007). As higher triglyceride is one crucial risk factor in the development of many diseases, including MetS, the polymorphism of the ApoA5 gene may be involved in MetS. Indeed, several studies in different populations have found the association between the ApoA5 polymorphism and the risk of MetS (Grallert et al., 2007; Yamada et al., 2007; Dallongeville et al., 2008; Hsu et al., 2008; Komurcu-Bayrak et al., 2008; Mattei et al., 2009; Kisfali et al., 2010; Niculescu et al., 2010; Ong et al., 2011; Vasilopoulos et al., 2011). However, these results are not quite consistent; therefore, we conducted a meta-analysis to clarify the exact association between the ApoA5 polymorphism and the risk of MetS.
Methods
Study selection
The PubMed, Embase, and Science Citation Index (ISI Web of Science) databases were searched to collect all publications on the association between the ApoA5 polymorphism and MetS (last search update: 11th February 2012). The language was limited to English only. References lists were further retrieved from identified publications to avoid missing any relevant articles. The following key words were used: (“Apolipoprotein A5” OR ApoA5); (Polymorphism OR SNPs OR “polymorphism, single nucleotide” OR variants OR genotype OR mutation); and (“Metabolic Syndrome X” OR “Metabolic Syndrome” OR MetS). Then, the three themes were combined by the Boolean operator AND. The included studies should meet the following criteria: (1) a case-control study; (2) only MetS as the outcome, and the investigated site should be −1131T>C and/or c.56C>G; (3) the genotype distribution of MetS and control can be obtained from articles directly or by calculation; (4) the control group without established diagnosis related to MetS; and (5) the genotype distribution of the control group should meet the Hardy-Weinberg equilibrium.
Data extraction
Two authors extracted data independently, and disagreements were resolved by discussion and consensus. The following information was extracted from publications: first author's name, year of publication, country of origin, ethnicity of population, definition of MetS, mean age of the subjects, gender component, and numbers of case and control with various genotypes.
Statistical analysis
Stata11.0 software (Stata; College Station, Texas) was used for meta-analyses. We pooled the overall odd ratios (ORs) and 95% confidence intervals (CIs) for −1131T>C (CC+TC) versus TT genotype and c.C56G (GG+GC) versus CC between the MetS and control group. Subgroup analysis was further performed by ethnicity. Heterogeneity among studies was evaluated by Chi-square-based Q test and I2 test. If there was no significant heterogeneity (p-value>0.05 and I2<25%), the ORs and 95% CIs would be estimated in a fixed-effect model. Otherwise, a random-effect model was chosen. Begg's funnel plot and Egger's test were used to evaluate the significance of the publication bias, with a p-value>0.05 considered as not significant.
Results
Characteristics of the included studies
The literature search identified 86 potential relevant articles (after deleting 81 duplicate articles). Sixty-two articles were excluded after initial screening. The remaining 24 studies were further reviewed by reading the full text, and 14 additional articles were excluded (see Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/gtmb). Because the articles by Grallert et al. (2007) and Ong et al. (2011) included two separate groups, respectively, these groups were considered as two independent studies. Finally, 12 studies from 10 articles (Grallert et al., 2007; Yamada et al., 2007; Dallongeville et al., 2008; Hsu et al., 2008; Komurcu-Bayrak et al., 2008; Mattei et al., 2009; Kisfali et al., 2010; Niculescu et al., 2010; Ong et al., 2011; Vasilopoulos et al., 2011) were included in our meta-analysis. Among 10 publications, only Ong’ study (Ong et al., 2011) chose the updated harmonized definition of MetS (Alberti et al., 2009). All other nine articles used the definition of NCEP ATPIII (Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults, 2001). A summary of characteristics of 10 articles is shown in Table 1.
ApoA5, apolipoprotein A5; MetS, metabolic syndrome; NA, not applicable.
Among the 12 studies, 11 studies from 9 articles (Grallert et al., 2007; Yamada et al., 2007; Hsu et al., 2008; Komurcu-Bayrak et al., 2008; Mattei et al., 2009; Kisfali et al., 2010; Niculescu et al., 2010; Ong et al., 2011; Vasilopoulos et al., 2011) involving 4374 MetS patients and 4628 no-MetS controls were included for the analysis of the −1131T>C polymorphism and MetS. 7 studies from 6 articles (Grallert et al., 2007; Komurcu-Bayrak et al. 2008; Mattei et al., 2009; Kisfali et al., 2010; Niculescu et al., 2010; Vasilopoulos et al., 2011) involving 3405 MetS patients and 5329 no-MetS controls were included for the analysis of the c.C56G polymorphism and MetS.
Meta-analysis
Figure 1 shows the results of the meta-analysis on the association of the −1131C>T polymorphism with MetS. A random-effect model was selected to assess the ORs and 95% CI. Compared to the TT genotype, the (CC+TC) genotype was significantly associated with MetS for the overall (OR=1.32; 95% CI: 1.14-1.53; p=0.000) with moderate heterogeneity (I2=54.9%, p=0.014). Subgroup analysis was further performed according to ethnicity. The association was still significant in Asians (OR=1.42; 95% CI: 1.25-1.62; p=0.000). However, we did not find a significant association in white populations (OR=1.25; 95% CI: 0.97-1.61; p=0.087).
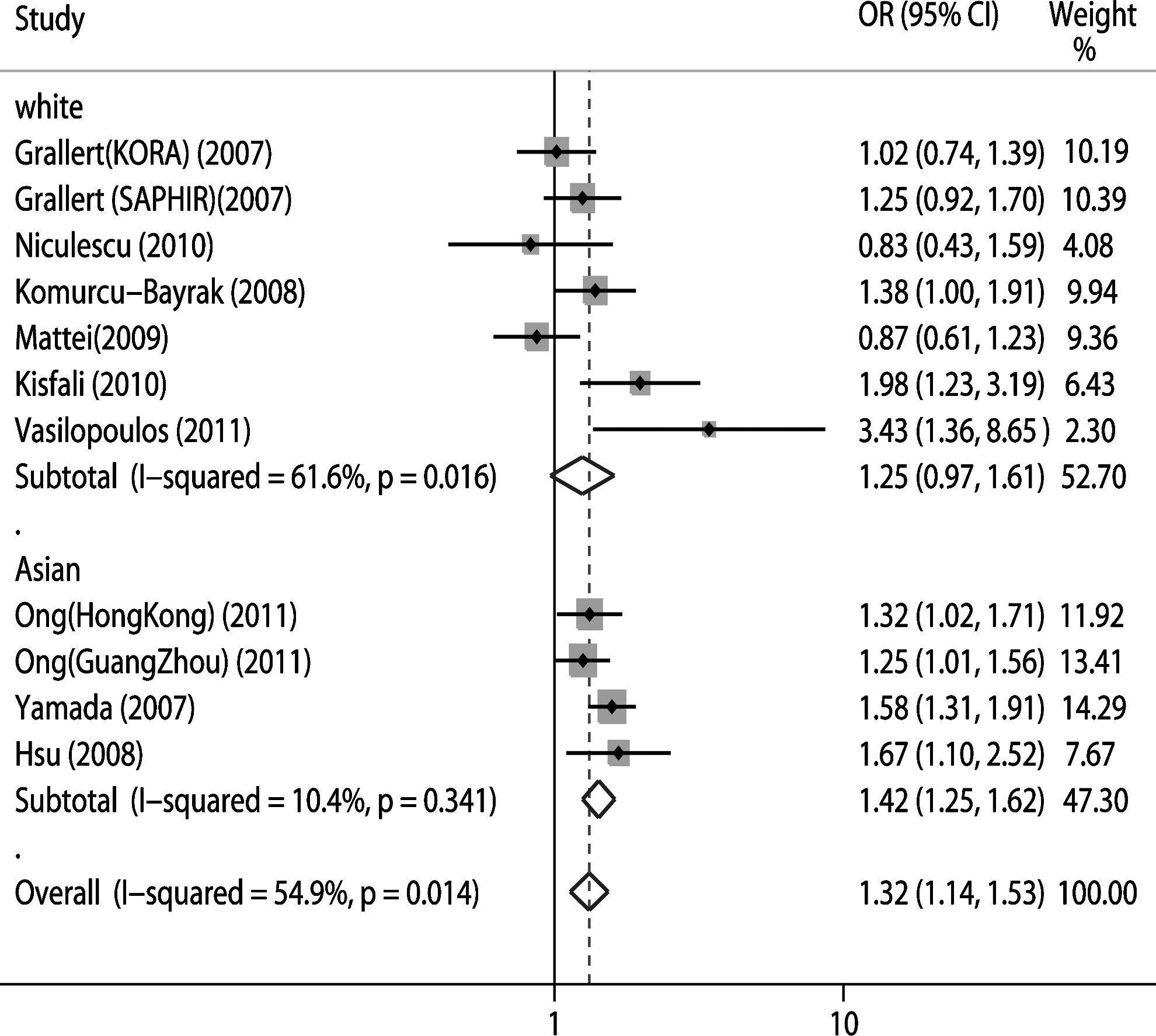
Forest plot of association between −1131T>C and metabolic syndrome (MetS). OR, odds ratio; CI, confidence interval.
Seven studies in white populations provided results on association of the c.C56G polymorphism with MetS. As is shown in Figure 2, the (GG+GC) genotype significantly increased the risk of MetS (OR=1.32; 95% CI: 1.15-1.50; p=0.000), and there was no evident heterogeneity (I2=0%, p=0.674).
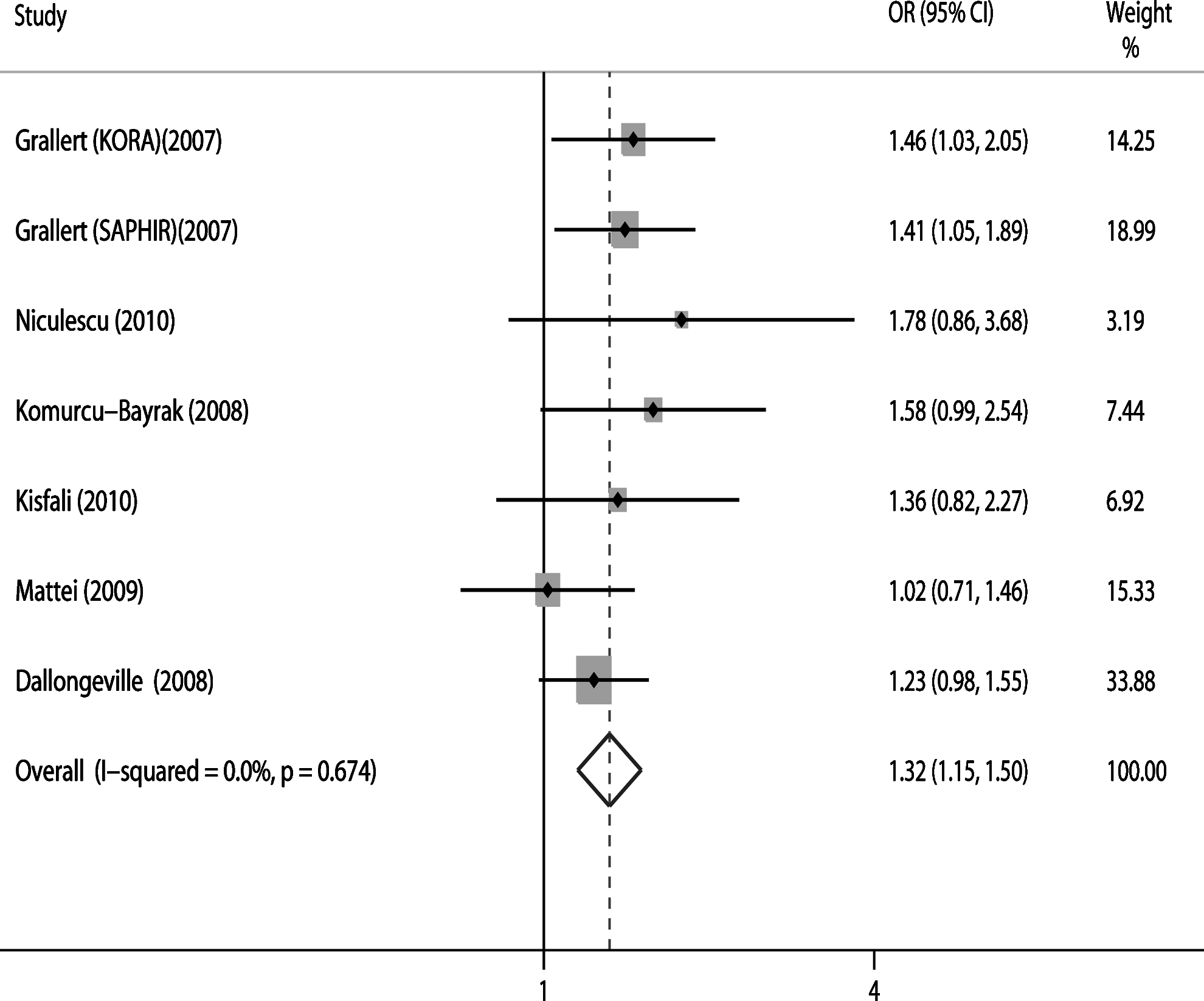
Forest plot of association between c.C56>G and MetS.
Begg's funnel plot and Egger's test were used to assess the publication bias of the studies. No significant publication biases were observed in either −1131T>C or c.C56G (Supplementary Figs. S2 and S3; Egger's test p=0.889; 0.289, respectively).
Discussion
ApoA5 was a new apolipoprotein gene first identified by comparative sequencing in 2001 (Pennacchio et al., 2001). The human ApoA5 gene consists of four exons and encodes a 366-amino-acid protein (apolipoprotein A-V). Function study in animals indicates that ApoA5 is a factor that can significantly reduce plasma triglyceride levels. Studies in humans (Baum et al., 2003; Matsunaga et al., 2007) further validated the strong association between the polymorphism of ApoA5 and plasma triglyceride levels. Recently, the association between ApoA5 variants and MetS was deeply investigated in different populations, but these studies drew inconsistent conclusions (Grallert et al., 2007; Yamada et al., 2007; Dallongeville et al., 2008; Hsu et al., 2008; Komurcu-Bayrak et al., 2008; Mattei et al. 2009; Kisfali et al., 2010; Niculescu et al., 2010; Ong et al., 2011; Vasilopoulos et al., 2011).
In our present study, we comprehensively investigated two common variants of ApoA5 with the risk of MetS. The polymorphism of −1131T>C seems significantly associated with MetS in Asians (OR=1.42; 95% CI: 1.25-1.62, p=0.000), but not in white populations (OR=1.25; 95% CI: 0.97-1.61, p=0.087), whereas the c.C56G polymorphism was significantly associated with MetS in white populations (OR=1.32; 95% CI: 1.15-1.50; p=0.000). Our analysis is in accordance with allelic frequency distributions in different ethnicities. As the rare allele frequency for −113T>C was at a low value (6%) in European Americans (Pennacchio et al., 2002) and at a relatively high value (34%) in Asians (Hubacek, 2005), the rare allele frequency for c.C56G was at high value (15%) in the white population (Pennacchio et al., 2002). No significant publication biases were observed in either −1131T>C or c.C56G (Supplementary Figs. S2 and S3; Egger's test p=0.889; 0.289, respectively).
The exact mechanism of how the ApoA5 gene polymorphism affects MetS is not fully clear. Maybe the association is due to the effect of the ApoA5 polymorphism on the dyslipidemic components of MetS (Dallongeville et al., 2008; Hsu et al., 2008). Possible mechanisms of ApoA5 on modulating triglyceride are as follows: first, ApoA5 might reduce hepatic very low-density lipoprotein production and secretion, as ApoA5 showed high lipid affinity and low solubility (Beckstead et al., 2003; Weinberg et al., 2003). Second, ApoA5 could accelerate plasma triglyceride hydrolysis by stimulating lipoprotein lipase activity or by modifying the effects or the concentrations of other apolipoprotein (Schaap et al., 2004; van Dijk et al., 2004). Third, ApoA5 could act as a ligand to lipoprotein receptors or proteoglycans and facilitate receptor-mediated endocytosis of lipoprotein particles (Lookene et al., 2005; Merkel et al., 2005; Nilsson et al., 2007).
Some limitations in our meta-analysis should be pointed out: firstly, the effects of gene-gene or gene-environment interactions were not evaluated. In fact, many other genes have been confirmed to be related with MetS, such as FTO (Zhou et al., 2012) and IL-6 (Hamid et al., 2005). Environmental factors, such as diets and physical inactivity, could also affect the development of MetS. Secondly, the results of −1131T>C in a white population subgroup should be interpreted with a caution for evident heterogeneity in between-studies. Thirdly, articles in other languages (not English) were not included in this meta-analysis. That may lead to bias, although the Begg's funnel plot and Egger's test showed no significant bias. Lastly, as our study mainly focused on MetS, we did not evaluate the ApoA5 genotype association with various parameters of MetS. Further studies are needed to investigate the association between the ApoA5 gene polymorphism and components of MetS, such as triglyceride levels and abdominal obesity.
In conclusion, our study suggested that the ApoA5 −1131T>C polymorphism was significantly associated with risk of MetS in Asians, but not in white populations, whereas the ApoA5 c.C56G polymorphism was significantly with MetS in white populations.
Footnotes
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.