
Abstract
Down syndrome (DS) is the most common cause of mental retardation. Recent reports have investigated possible genetic factors that may increase maternal risk for DS. Methionine synthase reductase (5-methyltetrahydrofolate-homocysteine methyltransferase reductase MTRR) plays an important role in folic acid pathway and a common polymorphism (c.66A>G) has been associated with DS but results were controversial. This meta-analysis summarizes the available data concerning this association. Online major databases were searched to identify case-control studies regarding MTRR 66A>G polymorphism and DS. Crude odds ratios (OR) and 95% confidence intervals (CI) were calculated for maternal risk to have a DS child both using fixed and random effects (RE) models. Eleven articles from six populations were identified, including 1226 DS mothers and 1533 control mothers. Heterogeneity among studies was significant (Q=29.7, DF=10, p=0.001; I2=66.3%). The pooled OR in a RE model showed an increase in the risk of having a DS child associated with the G allele (OR 1.23, 95% CI 1.02-1.49). The fixed effect pooled OR was 1.19 (95% CI 1.08-1.31). This meta-analysis indicates that maternal MTRR 66A>G polymorphism is associated with an increased risk of having a DS child.
Introduction
D
It has been proposed that an altered maternal folate metabolism could be associated to centromeric DNA hypomethylation and chromosomal nondisjunction (James et al., 1999). Abnormal folate metabolism could also counteract the overexpression of the three copies of the cystathionine beta synthase gene (CBS) in the trisomy 21 fetus, assuring folate availability for both DNA synthesis and methylation (Hobbs et al., 2000).
From the original paper of James et al. (1999) to date, several reports have investigated the influence of folate pathway polymorphisms on maternal risk for DS and the most studied genes are methylenetetrahydrofolate reductase (MTHFR) and 5-methyltetrahydrofolate-homocysteine methyltransferase reductase, also known as methionine synthase reductase (MTRR). Results of the association between single nucleotide polymorphisms within these genes and the risk of DS are controversial and discrepancies among reports have been explained mostly by the nutritional environment and genetic characteristics of the populations (Guéant et al., 2003).
The MTRR gene was mapped to chromosome 5p15.2-15.3 (Leclerc et al., 1998) and encodes an enzyme responsible for restoration of methionine synthase (MTR) activity by reductive methylation of cobalamin, leading to methionine synthesis and production of S-adenosylmethionine, which is the main cellular methyl donor for transmethylation reactions (Wolthers and Scrutton, 2007). A common polymorphism in human MTRR is an adenine to guanine transition at position 66 (c.66A>G), which results in replacement of isoleucine with methionine at residue 22 (p.I22M) (Wilson et al., 1999). This substitution decreases the ability of MTRR to restore MTR activity in vivo (Olteanu et al., 2002).
In the last decade several reports have evaluated the association between MTRR 66A>G polymorphism and an increased risk of having a DS child, and the results are controversial. This meta-analysis summarizes published data concerning this association.
Methods and Analyses
Eligible studies were identified by searches in major literature databases (PubMed at www.ncbi.nlm.nih.gov/pubmed and Web of Science at www.isiknowledge.com), using the following keywords, alone and combined: “A66G,” “66A>G,” “MTRR,” “methionine synthase reductase,” “5-methyltetrahydrofolate-homocysteine methyltransferase reductase” and “Down syndrome” or “trisomy 21.” SCIELO (at www.scielo.org) was also screened using the same keywords in English, Portuguese and Spanish in order to assure the recovery of papers published in Latin American journals not indexed in the other databases. We included studies that met the following criteria: (1) case-control studies; (2) use of validated genotyping methods to identify the polymorphism; (3) availability of MTRR genotypes for Down syndrome mothers (DSM) and control mothers (CM), and (4) written in English, Portuguese or Spanish. Both authors have reviewed independently each article. Crude odds ratios (OR) and 95% confidence intervals (CI) for having a DS child were calculated comparing GG and/or AG with AA and comparing G allele with A allele. Heterogeneity among studies was tested considering that when there is heterogeneity among studies, the pooled OR is preferably estimated using the random effects (RE) model instead of fixed effects (FE) model. Q-statistics and I2 metrics were calculated as described (Cochran, 1954; Higgins and Thompson, 2002). Statistical analyses were performed using Meta-Disk (version 1.4). The frequencies were evaluated for accordance to Hardy-Weinberg equilibrium (HWE) using the χ2 test.
Results
We screened major bibliographic databases searching for articles focusing on the association of maternal MTRR 66A>G polymorphism and DS. We found 11 reports that met the inclusion criteria conducted in populations of different ethnic backgrounds from seven countries: United States and Canada (Hobbs et al., 2000), Ireland (O'Leary et al., 2002), France (Chango et al., 2005), China (Wang et al., 2008), Italy (Scala et al., 2006; Coppedè et al., 2009; Pozzi et al., 2009) and Brazil (da Silva et al., 2005; Santos-Rebouças et al., 2008; Brandalize et al., 2010; Zampieri et al., 2012). The reports included 1233 DS and 1533 CM and details of the studies are presented in Table 1.
p-Value for Hardy-Weinberg equilibrium in CM group.
DSM, Down syndrome mother; CM, control mother.
The observed genotype frequencies were evaluated for accordance to HWE. The distribution of genotypes were in agreement with HWE in all studies but one (Chango et al., 2005) (χ2=12.19, p=0.002).
The frequency of heterozygous genotype was the highest in both DSM and CM, ranging from 42.7% to 60.0% in DSM and 45.4% to 56.3% in CM. The genotype and allele frequencies are shown in Table 2.
Overall analysis of the association between the mutant allele and an increase in risk of DS has revealed significant heterogeneity among studies (Q=29.7; DF=10; I2=66.3%; p=0.001), and the RE pooled OR was significant, indicating a 20% increase (OR 1.23; 95% CI 1.02, 1.49) in the risk of having a DS child (Fig. 1).
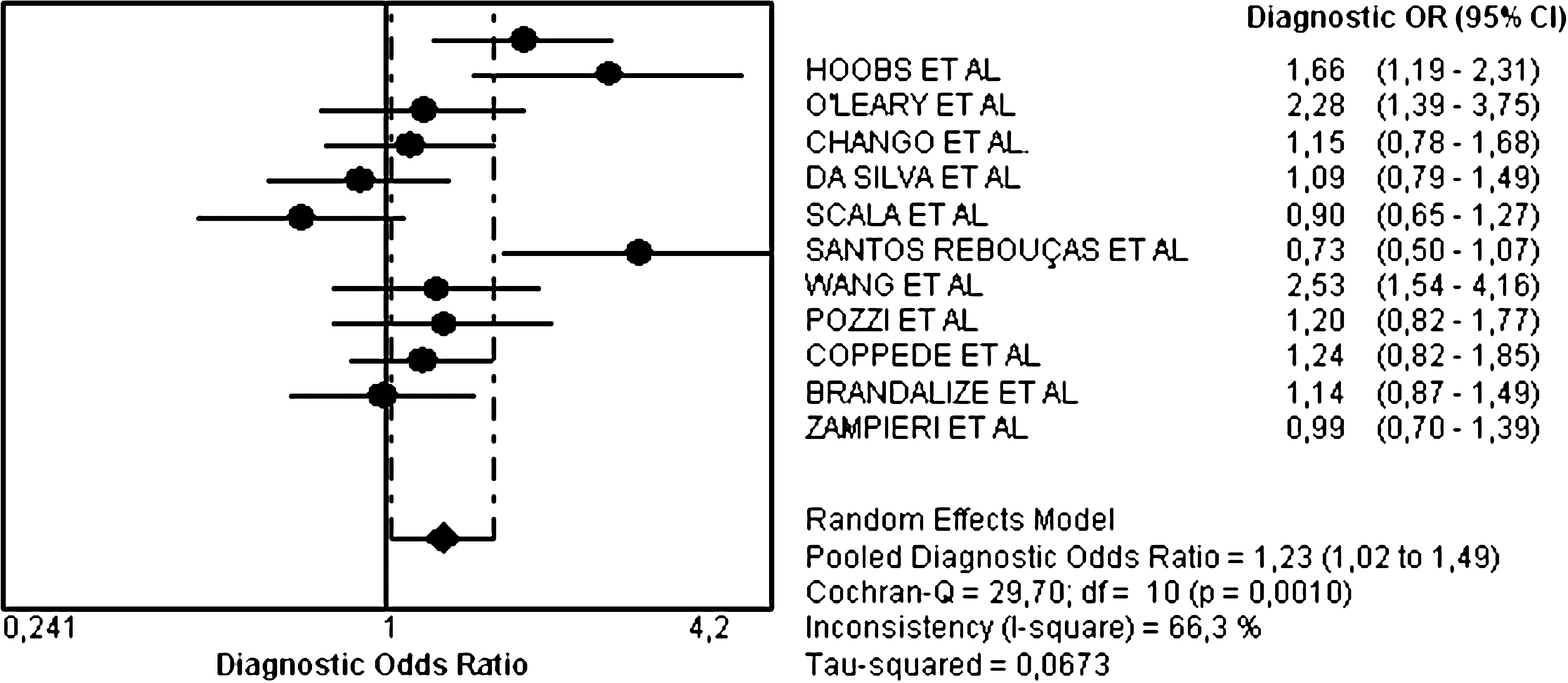
Random effects pooled odds ratio (OR) and 95% confidence intervals forest plot for the association between methionine synthase reductase 66A>G and Down syndrome in case-control studies. The OR estimates are represented by solid black circles and the size of the symbol indicates the weight of the respective study in the meta-analysis. The pooled OR is represented by a solid black diamond.
We also evaluated the association using different interaction models; recessive (GG vs. AG+AA), dominant (GG+AG vs. AA) and codominant (both GG vs. AA and AG vs. AA). The heterogeneity between-studies was significant using the recessive (Q=25.8, DF=10, p=0.004, I2=61.2%) and dominant (Q=19.1, DF=10; p=0.04, I2=47.6) models. No heterogeneity was observed when AG genotype was compared to AA (Q=12.2, DF=10, p=0.27, I2=17.8%) and the FE pooled OR was significant (OR 1.21, 95% CI 1.02-1.44) (Table 3).
CI, confidence interval; OR, odds ratio.
Discussion
DS is an important public health issue and most DS individuals need special services (education, medical, and social) for the duration of their lives. Since the first report of a possible association between genetic polymorphisms in folate-metabolizing encoding genes and an increased DS risk (James et al., 1999), several studies have investigated this hypothesis.
The distributions of MTRR 66A>G genotypes vary greatly according to genetic background. The higher frequencies of MTRR GG genotype were described in Caucasians and Indians ranging from 28% to 35%, and the lowest (less than 7.5%) were observed in Latin descent populations (Rady et al., 2002; Yang et al., 2008; Rai et al., 2011). We also observed differences in MTRR GG genotype frequencies among groups of different ethnicities, ranging from 14.7% in the Chinese population to 40.3% in French population.
Hobbs et al. (2000) reported the association of maternal MTRR 66A>G polymorphism with DS risk in North American population (including samples from 16 United States and Canada). The comparison of genotypic distribution between DSM and CM indicated an increase of 2.57-fold (95% CI 1.33-4.99) for women with GG genotype. The risk was increased to 4.08-fold (95% CI 1.94-8.56) when the homozygous GG genotype was combined to MTHFR TT genotype suggesting a multiplicative effect. This pattern of association was observed in the Chinese population, where maternal GG genotype increased 5.2-fold (95% CI 2.06-17.50) the risk. The multiplicative effect of MTHFR TT and MTRR GG genotypes were also observed, leading to a sixfold (95% CI 2.06-17.50) increased risk (Wang et al., 2008). An impressive 15-fold (95% CI 1.94-116.0) increased risk associated with homozygous GG genotype was observed in the Irish population (O'Leary et al., 2002).
In France, there was no significant difference between DSM and CM MTRR G allele frequencies (Chango et al., 2005). The authors emphasized the importance of correlating homocysteine (Hcy) and folate levels to increase the sensitivity to detect a correlation between genotype and DS risk.
Four studies were conducted in Brazil. da Silva et al. (2005) have evaluated five folate metabolizing pathway polymorphisms (MTHFR 677C>T, MTHFR 1298A>C, MTRR 66A>G, MTR 2756A>G and CBS 844ins68) and observed higher Hcy levels among DSM when compared to CM. However, the statistical difference was associated only with a MTHFR TT genotype. For MTRR 66A>G, no difference was observed for both genotype and allele distributions between the case and control groups, indicating that this polymorphism did not act as an independent risk factor for DS. Santos-Rebouças et al. (2008) evaluated folate pathway polymorphisms combined to nutritional deficiency as maternal risk factor for DS and did not found an independent or combined association of maternal genotypes to an increase risk of DS birth. In a sample of mothers of European descent living in the South region of Brazil, Brandalize et al. (2010) have evaluated four folate metabolizing polymorphisms (MTR 2756 A>G, MTRR 66A>G, CBS 844ins68 and RFC 80 G>A) and found that individual polymorphisms are not associated with DS. Zampieri et al. (2012) have studied 12 polymorphisms in folate pathway and their influence on serum folate and plasma methylmalonic acid (MMA) concentrations as an indicator of high Hcy levels. Although the authors have found that polymorphisms in folate metabolism genes modulate the maternal risk for bearing a child with DS, no independent association between MTRR 66A>G polymorphism and DS risk was observed. However, this polymorphism was the only one to affect MMA concentration, validating the proposed association between the presence of the mutant 66 G allele both in heterozygous and homozygous status with higher Hcy concentrations.
Three reports concerning MTRR 66A>G polymorphism and an increased risk of having a DS child were conducted in the Italian population (Scala et al., 2006; Coppedè et al., 2009; Pozzi et al., 2009). While Scala et al. (2006) and Coppedè et al. (2009) did not found association between MTRR 66A>G polymorphism and DS, Pozzi et al. (2009) have found that the presence of the mutated G allele increases two-fold the risk (95% CI 1.11-4.40) for a DS offspring after parity adjustment, with similar trends when analyses focuses on woman aged less than 35 years old. On the other hand, the association was not observed by Coppedè et al. (2009) when a sample of Italian woman <35 years at conception was evaluated.
Crude data analysis has showed association of MTRR 66A>G and DS in USA and Canada, Ireland and China populations (Hobbs et al., 2000; O'Leary et al., 2002; Wang et al., 2008) while in the remaining studies, all conducted in Latin European descent populations (Chango et al., 2005; da Silva et al., 2005; Scala et al., 2006; Santos-Rebouças et al., 2008; Coppedè et al., 2009; Pozzi et al., 2009; Brandalize et al., 2010; Zampieri et al., 2012), only one study reported an association of MTRR 66A>G and DS, after parity adjustment (Pozzi et al., 2009). However, a previous report on the Italian population in a sample of greater size has denied this association (Coppedè et al., 2009).
Controversial results were also observed in meta-analyses published to date. Zintzaras (2007) condensed the results of five reports and included 559 DSM and 866 CM and did not find any evidence for the association between MTRR 66A>G and DS. On the other hand, a recent report by Rai (2011), using data extracted from six reports with a total of 623 DSM and 936 CM describe a 1.42-fold increase in DS offspring when mothers carry G allele. We found a result similar to that described by Rai (2011), with a significant increase of 1.23-fold (95% CI 1.02-1.47) of having a DS child for woman carrying MTRR 66G allele.
It was previously proposed that the association of MTHFR 677C>T polymorphism with neural tube defects could be specific for non-Latin European descent populations (Amorim et al., 2007). We can imagine a similar scenario for the association between MTRR 66A>G and DS. When non Latin European descent reports were excluded (three studies), the RE pooled OR decreases and looses significance (data now shown). The occurrence or absence of association between a polymorphism in a gene involved in folate metabolism with DS could be explained by difference in maternal folate intake, variation in mutant allele frequency in the population, genetic heterogeneity of studied population and different gene-nutrient interaction among populations (Amorim et al., 2007). Correlation of maternal red blood cell folate with polymorphic genotypes will help to identify the role of these genetic variants as risk factors for DS, and only few reports perform this correlation.
The large between-studies heterogeneity we have observed may be due to discrepancies in study design but it could reflect genuine differences among the studied populations (Zintzaras, 2007). Our meta-analysis indicates an independent association between MTRR G allele and an increased risk of DS, and the populations without Latin European descent could be at greater risk.
Footnotes
Acknowledgments
This study was supported by Fundação de Amparo a Pesquisa do Estado do Rio de Janeiro—FAPERJ, Brazil (No. E-26/110.427/2011).
Author Disclosure Statement
None declared.