
Abstract
Purpose: To determine the allele frequency of three codon-changing variants (CYP2C8*2, CYP2C8*3, and CYP2C8*4) in the Han, Uighur, Hui, and Mongolian Chinese populations, and compare genetic polymorphism differences between the Han and minority Chinese ethnicities. Methods: Five hundred seventy four healthy unrelated volunteers from four major nationalities (Han: 136; Uighur: 153; Hui: 158; Mongolian: 127) in China were recruited. The study was approved by the local research ethics committee. DNA was extracted from peripheral leukocytes using a standard protocol. A polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) method was developed to detect three alleles of CYP2C8. PCR results were confirmed by subsequent direct DNA sequencing. Results: The three minorities showed the CYP2C8 polymorphism with an allele frequency as follows: the allele frequencies of CYP2C8*2, CYP2C8*3, and CYP2C8*4 were 0.33%, 2.94%, and 2.29% in Uighur; 0.39%, 1.57%, and 1.18% in Mongolian, and 0%, 1.58%, and 0.63% in Hui, respectively. For Han, CYP2C8*2, CYP2C8*3, and CYP2C8*4 were absent. Conclusion: To the best of our knowledge, the current study described polymorphisms of CYP2C8 in Chinese minorities for the first time. The results showed that there were significant ethnic differences in the distribution of CYP2C8 in the Han and three minorities, that is, the Uighur, Hui, and Mongolian Chinese populations.
Introduction
G
CYP2C8 is one of the important CYPs, accounting for 7% of the total hepatic CYP content in human. It is responsible for the oxidative metabolism of more than 60 clinically available drugs, and typical substrates for that CYP2C8 include thiazolidinediones, meglitinides, Non-Steroid Anti-Inflammatory Drugs, antimalarials, and chemotherapeutic taxanes, etc. CYP2C8 also contributes to the metabolism of endogenous substrates, such as arachidonic acid, steroids, and retinoids (Daily and Aquilante, 2009). The human CYP2C8 gene has been mapped to chromosome 10q24 and is composed of nine exons (Klose et al., 1999). Polymorphisms in CYP2C8 were first determined in 2001 (Dai et al., 2001), and so far, in addition to the wild-type allele, more than 20 single-nucleotide polmorphisms (SNPs), including CYP2C8*2 through CYP2C8*14 and several that have not yet been assigned a star designation have been identified, mainly in exons 3, 5, and 8 (http://cypalleles.ki.se/CYP2C8.htm). CYP2C8 genetic polymorphisms have been implicated in the variability of CYP2C8 activity with different phenotypes. Of these, CYP2C8*1 is the first reported wild-type CYP2C8 gene sequence with a normal enzyme activity. CYP2C8*2 carries an 805A>T mutation leading to I296F. Compared with the CYP2C8*1 enzyme, the CYP2C8*2 enzyme shows a threefold higher km and a sixfold lower intrinsic clearance for amodiaquine (Parikh et al., 2007). CYP2C8*2 has also been associated in vitro with decreased metabolism of paclitaxel (Dai et al., 2001). The CYP2C8*3 allele contains 416G>A and 1196A>G mutations causing R139K and K399R, respectively. Clinical data suggest that the CYP2C8*3 allele is associated with increased metabolism of the CYP2C8 substrates rosiglitazone, pioglitazone, and repaglinide (Aquilante et al., 2008; Tornio et al., 2008; Tomalik-Scharte et al., 2011). While some studies have showed that CYP2C8*3 is associated with reduced clearance of paclitaxel, arachidonic acid, and ibuprofen (Dai et al., 2001; Martínez et al., 2005; Bergmann et al., 2011). CYP2C8*4 has a 792C>G substitution leading to Ile264Met, showing a lower metabolic activity for paclitaxel than the wild-type enzyme, although no statistical significance has been found (Bahadur et al., 2002). In addition to drug metabolism, CYP2C8 polymorphisms also play an important role in some disease, such as cardiovascular disease and diabetes. A study showed that there was a significant association in carriers of CYP2C8*3 and CYP2C9*2 or *3 alleles and an increased risk of acute myocardial infarction in male subjects (Theken and Lee, 2007). In diabetes patients, frequency of the CYP2C8*4 allele has markedly increased compared with those of the normal population (Weise et al., 2010).
Ethnic differences in the CYP2C8 genotype frequencies have been observed in a number of populations. For instance, CYP2C8*3 has been found in 10%-20% of Caucasians, 0.3%-2% of Africans, but seems to be absent in Asians. Similar to CYP2C8*3, CYP2C8*4 occurred primarily in Caucasians with the frequency of about 6% (Bahadur et al., 2002; Cavaco et al., 2006; Pechandova et al., 2012). CYP2C8*2 is common in Africans (frequency of 6%-28%), but is rare in other races studied (Cavaco et al., 2005; Kudzi et al., 2009). In regard to less common polymorphisms, such as CYP2C8*5 through CYP2C8*14, these polymorphisms are rare (<0.5%) and appear to be race specific, occurring primarily in Asian populations (Nakajima et al., 2003; Saito et al., 2007; Yeo et al., 2011). For Chinese, CYP2C8 polymorphisms in Han have been studied, and the results show the conservative distribution of CYP2C8 revealing that three major active CYP2C8 genotypes, namely, CYP2C8*2, CYP2C8*3, and CYP2C8*4 are absent in the Han population. As known, China is a multinational country with 55 ethnic minorities besides the Han majority. All the nationalities have their own genetic origin, cultural background, diet source, habit, and geographic environment. The relatively unique genetic, cultural, dietetic, and environmental characteristics of each nationality should affect the catalytic activity of CYP450, including CYP2C8. However, there is little information concerning CYP2C8 polymorphisms in the multiracial population of Chinese thus far.
In this study, we first determined the allele frequency of three codon changing variants: CYP2C8*2, CYP2C8*3, and CYP2C8*4 in Uighur, Hui, and Mongolian nationalities of the Chinese population. To explore the variety among different nationalities, the CYP2C8 distribution in Han has also been screened, and chi-square testing was used to compare the allele and genotype frequencies in the four nationalities.
Methods
Chemical and reagents
TIANamp Whole Blood DNA Extraction Kit was obtained from Tiangen Biotech Co., Ltd. A polymerase chain reaction (PCR) Master Mix (2×) was purchased from Fermentas. Restriction enzymes were purchased from New England Biolabs for BseRI (Fermentas) for MboI and TaqI. All other reagents were of high grade and commercially available.
Subject
Five hundred seventy four subjects, including four different nationalities in China (Han [n=136], Uighur [n=153), Hui [n=158), and Mongolian [n=127]) were used in this study. All participants gave written informed consent, which was approved by the independent Ethics Committee of the General Hospital of Shenyang Military Region and followed the recommendations of the Declaration of Helsinki. The subjects ranged in age from 20 to 26 years and in body mass index from 20 to 24. All the participants were healthy, according to their medical history, physical examination, and laboratory tests.
DNA isolation
DNA was extracted from peripheral lymphocytes using the TIANamp Whole Blood DNA Extraction Kit. DNA concentrations were determined by UV spectrophotometry. The DNA purity was more than 99.6%.
Genotyping
The polymerase chain reaction-restriction fragment length polymorphism method developed by Nakajima et al. (2003) with minor modification was applied for the genetype analysis of CYP2C8. The DNA sequence for each primer was given in Table 1. All PCR reactions were generated by a GeneAmp PCR system 2700 Thermal Cycler (Applied Biosystems). The PCR condition for CYP2C8*3 was as follows: 33 cycles of 94°C for 40 s, 55°C for 40 s and 72°C for 40 s. For CYP2C8*2 and CYP2C8*4 was 30 cycles of 94°C for 30 s, 55°C for 30 s and 72°C for 30 s. The resulting PCR product was restricted with specific restriction endonuclease as follows: for CYP2C8*3, the 467-bp PCR products of CYP2C8*1 were digested by BseRI into fragments of 310 and 157 bp, while the PCR products of the homozygous Arg139Lys allele remained uncut followed by resolving on 2% agarose gels. For CYP2C8*2, the resulting 167-bp products were digested with MboI and resolved on 12% polyacrylamide. MboI cut the PCR products from the wild-type CYP2C8*1 allele into 69-, 65-, and 33-bp fragments, whereas the PCR products from the homozygous were cut into 98 and 69 bp. For CYP2C8*4, TaqI digested the wild-type CYP2C8*1 167-bp PCR products into 85-, 53-, and 31-bp fragments, while the PCR products of the homozygous Lys399Arg allele were cut into 136 and 31 bp followed by resolving on 4% agarose gels (Fig. 1).
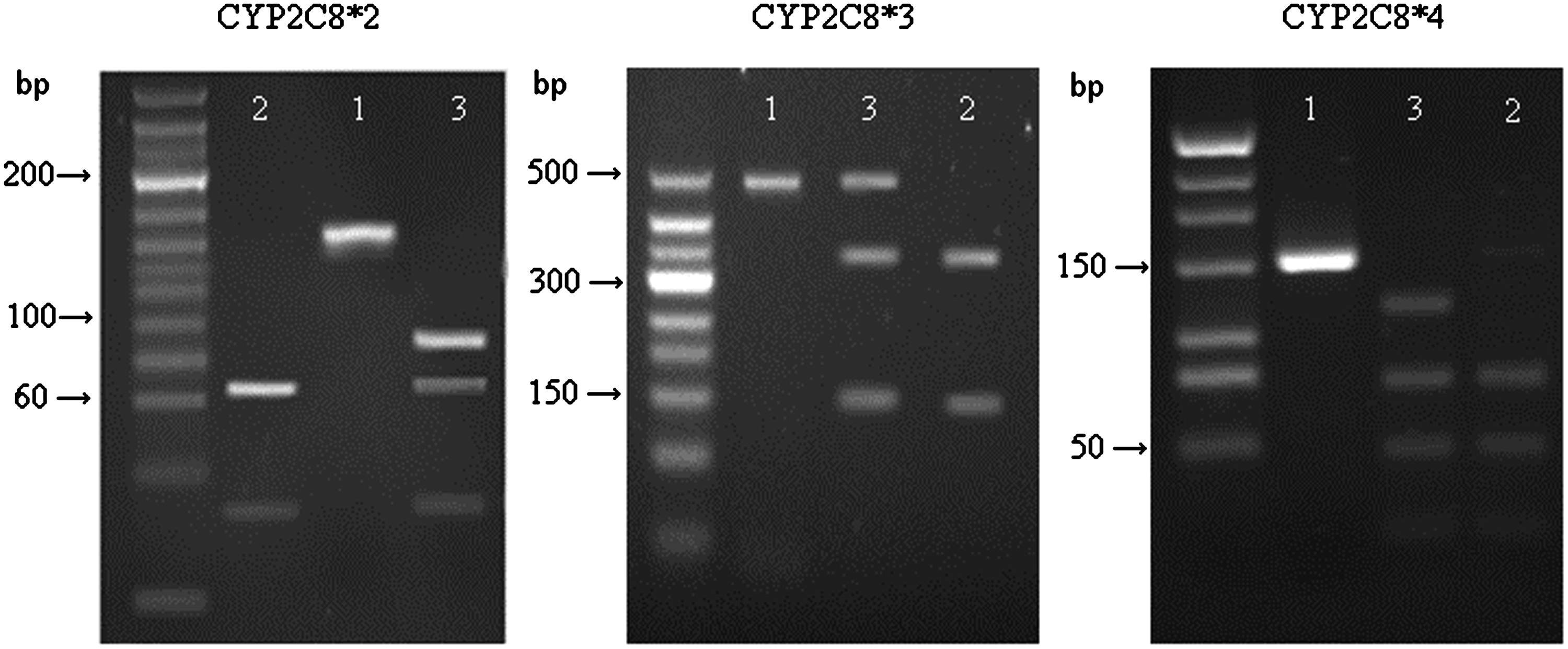
Polymerase chain reaction (PCR) products digested with restriction endonuclease of CYP2C8*2, CYP2C8*3, and CYP2C8*4. Lanes 1 showed a nondigested PCR product; line 2 showed the fragment after digestion with restriction endonuclease for wild-type; and line 3 showed the fragment after digestion with restriction endonuclease for heterozygous type. No homozygous type was detected in this study.
PCR, polymerase chain reaction.
Statistical analysis
Data were compiled according to the genotype and allele frequencies. Expected genotype frequencies were calculated using the Hardy-Weinberg equation from the allele frequencies (p2+2pq+q2=1). Chi-square testing was used to compare allele frequencies between different nationalities population. A p value of 0.05 or less was regarded as significant. All statistics were performed using SPSS 17.0 (SPSS, Inc.).
Result
DNAs from the 574 healthy Chinese volunteers were successfully amplified. Using this method, the CYP2C8 variant was detected in all the four minorities. The genotype frequencies of CYP2C8*1, CYP2C8*2, CYP2C8*3, and CYP2C8*4 in the Han, Uighur, Hui, and Mongolian Chinese populations were summarized in Table 2, and allele frequencies of CYP2C8*1, CYP2C8*2, CYP2C8*3, and CYP2C8*4 in the Han, Uighur, Hui, and Mongolian Chinese populations were summarized in Table 3. The deviations from Hardy-Weinberg equilibrium genotype frequencies for the various SNPs were assessed using the chi-squared test, and the genotypes were found to be in equilibrium.
Predicted frequencies calculated according to the Hardy-Weinberg equation.
For Hans, CYP2C8*2, CYP2C8*3, and CYP2C8*4 were absent, which was consistent with those of previous studies. In addition of Han, CYP2C8*2 was also absent in Hui. CYP2C8*3 and CYP2C8*4 were detected in all the three minorities. The chi-squared test was performed to determine the difference of the allele distribution of CYP2C8 among the four nationalities, which indicated the significant ethnical differences in the distribution of CYP2C8*2, CYP2C8*3, and CYP2C8*4 genotypes in three minorities, namely, Uighur, Hui, and Mongolian compared with the Han Chinese population. However, except for CYP2C8*2, no significant differences were observed among the Uighur, Hui, and Mongolian populations for the distribution of CYP2C8*3 and CYP2C8*4. For CYP2C8*2, the results from Uighurs were similar to those of Mongolian's. No homomutant type was detected in all the four nationalities.
Discussion
Ethnicity was an important variable contributing to interindividual variability in drug metabolism, response, and toxicity. Many SNPs in important CYP genes, including CYP3A4, CYP2C9, and CYP2B6, account for interethnic variations in response to a large number of therapeutic agents with marked ethnic differences. More recently, a study concerning the polymorphism distribution of CYP450 among different minority populations has been completed in our laboratory, which showed the unequal distribution of CYP3A4, CYP2C9, CYP2C19, and CYP2D6 in the Han, Uighur, Hui, and Mongolian Chinese populations (Zuo et al., 2012). As a continuation of these studies, we have studied the distribution of CYP2C8 in the four Chinese populations mentioned above in this article, and this is the first to estimate the distribution of CYP2C8 in the Chinese minorities.
Our data suggested that the genetic polymorphism of CYP2C8 was present in the Uighur, Hui, and Mongolian populations, but not in Han Chinese. Han Chinese is the largest population in China, accounting for ∼98% of the total Chinese population with a total of 1.2 billion. Results from this study showed that Han lack CYP2C8*2, CYP2C8*3, and CYP2C8*4 alleles; this result was similar to those of other Asian populations, including Japanese and Korean (Nakajima et al., 2003; Saito et al., 2007; Yeo et al., 2011) and also confirmed the results of a previous study. Different from Han, CYP2C8*2, CYP2C8*3, and CYP2C8*4 were detected in the Uighur and Mongolian populations. While for Hui, CYP2C8*3 and CYP2C8*4 were found. In view of heredity, Uighur have their roots in Caucasians and, thus, show a genetic makeup that is more akin to the Caucasians. However, interestingly, different results were reported in this study. CYP2C8*2 has an allelic frequency of 0.3% in Uighur, which was similar to those of other Caucasian populations. CYP2C8*3, the most studied allele gene of CYP2C8 mainly occurred in Caucasians. For Uighur, the allelic frequency was 2.9%, which was significant lower than Caucasians, such as American (13%) (Dai et al., 2001), Portugal (19.8%) (Cavaco et al., 2006), Spanish (17.4%) (Dorado et al., 2008), and Czechs (10.9%) (Pechandova et al., 2012). Similar to CYP2C8*3, the CYP2C8*4 mutant in Uighurs had an allelic frequency of 2.3%, lower than Caucasians (about 6%). The reason for the relative lower frequency of CYP2C8*3 and CYP2C8*4 among Uighurs is unknown, but might be due to geographical and environmental differences. Mongolian is one of the major nationalities living in Northeast Asia. Beside the Republic of Mongolia, the Mongolian population mainly distributes in north China and Russia, and the total population is more than 10 million. In view of genetic distance, Mongolian is close to Han (Du et al., 1998); however, results from this study showed that the allele gene distribution of CYP2C8 in Mongolians was significantly different from Hans, but similar to Uighers and Huis. All the three mutants were detected in the Mongolian population with the allelic frequency of 0.39%, 1.57%, and 1.18% for CYP2C8*2, CYP2C8*3, and CYP2C8*4, respectively. The allelic frequency of CYP2C8*3 and CYP2C8*4 in Mongolians was slight lower than those of Uighers, but no statistical significance was found (p>0.05). Huis, derived from Persians and Arabs, is one of the major minorities of China. In the long-term historical process, Huis have combined the living customs of the Han, Mongolian, and Uigher. In this study, the results showed that the feature of CYP2C8 distribution in Hui was between Han and Mongolian. Similar to Han, CYP2C8*2 was absent in the Hui. The allelic frequency of CYP2C8*3 mutant was closer to Mongolian (1.58% vs. 1.57%). CYP2C8*4 has an allelic frequency of 0.63%, lower than Mongolian (1.18%), but no statistical significance was found.
From the results mentioned above, it could be seen that the CYP2C8 polymorphism is very well conserved among all four nationalities in China, and even no homomutant type was detected. The conservative distribution of CYP2C8 may be interpreted from the following two aspects. One is the fact that CYP2C8 polymorphic sites could be situated at positions, which were very well conserved. As the major enzyme in the liver and kidney, CYP2C8 plays an important role in the metabolism of arachidonic acid to biologically active epoxyeicosatrienoic acids (EETs). EETs contributed to a series of physiological processes, including hepatic glycogenolysis, peptide hormone secretion in the pancreas and pituitary, Na+ transport and water reabsorption in the kidney, platelet aggregation, vascular inflammation, and vascular smooth muscle tone. Polymorphism in CYP2C8 could therefore potentially produce dramatic physiological and pathological changes in humans. Thus, to be protected from potential myocardial infarction and hypertension, the CYP2C8 gene must be very well conserved in the surviving individuals. This may be the other cause of the conservative distribution of CYP2C8 (Muthiah et al., 2005).
In summary, we determined allele and genotype frequencies of CYP2C8*2, CYP2C8*3, and CYP2C8*4 in the Han, Uighur, Hui, and Mongolian Chinese populations. Our findings confirm the existence of interethnic differences in the CYP2C8 allele and genotype frequencies in the Han, Uighur, Hui, and Mongolian Chinese populations. Our study provides the basic genetic information for further pharmacogenomic investigations in the Chinese population. Further studies are required to explore the clinical significance of those differences for treatment outcome and optimal dosage of drugs metabolized by CYP2C8 enzymes. It is also of potential clinical importance to be able to identify individuals who have altered pharmacokinetics for CYP2C8 substrates so that appropriate dosage strategies for these drugs can be adopted and adverse drug reactions can be avoided. In addition, this study seems to be of some significance to the study of some diseases.
Footnotes
Acknowledgments
The authors sincerely thank all the subjects for their participation in this study and all the medical staff who assisted in specimen collection.
Author Disclosure Statement
No competing financial interests exist.