
Abstract
Background: Genetic factors are involved in the etiology of Mycobacterium tuberculosis infection. Recently, ALOX5 has been identified as a candidate gene for tuberculosis (TB) susceptibility. We investigated whether an association between ALOX5 and TB exists in a Chinese pediatric population from northern China. Methods: We conducted a case-control study comprising 488 individuals aged 2 months to 17 years by genotyping 18 tag-single-nucleotide polymorphisms (SNPs) from the ALOX5 gene. The tag-SNPs were selected from the international HapMap project. An Illumina BeadXpress Scanner was utilized for genotyping, supported by the high-density BeadArray technology in combination with an allele-specific extension, adapter ligation, and amplification assay. Statistical analyses were performed to determine correlations between genetic variation and disease. Results: Our study is the first to show that ALOX5 is associated with susceptibility to pediatric TB in a subset of children in northern China. The rs2115819 T allele of ALOX5 presents a risk factor for childhood TB disease.
Introduction
The enzyme 5-lipoxygenase (5-LO, encoded by the ALOX5 gene) contains three nuclear localization sequences (NLSs) and a phosphorylation site (ser271) involved in nuclear localization (Matsumoto et al., 1988). In situ hybridization studies with rat lung tissue show that the protein only localizes to macrophages and that it is a key enzyme catalyzing reactions involved in the formation of leukotrienes (LTs) and lipoxins (LXs) (Voelkel et al., 1996; Luo et al., 2003). ALOX5 is considered to play a critical role in inflammatory and immune disease pathology, something LTs and LXs participate in. It is possible that mutation of the ALOX5 gene could reduce synthesis of LTs and LXs. For example, mutation of either NLS1 or ser271 was found to reduce nuclear localization of 5-LO, while decreasing leukotriene B4 (LTB4) synthesis in transfected mouse fibroblasts (Matsumoto et al., 1988). After M. tuberculosis infection, LTB4 and lipoxin A4 (LXA4) levels increased in murine sera, but not in 5-LO-deficient mice. Importantly, the 5-LO-deficient mice displayed an increased resistance to M. tuberculosis infection (Bafica et al., 2005).
The ALOX5 gene spans approximately 82 kb of the chromosome 10 10q11.2 locus and has 14 exons and 13 introns. ALOX5 polymorphisms are associated with TB in Ghana (Herb et al., 2008). Although 5-LO-dependent LT and LX synthesis could play a major role in the immune modulation of M. tuberculosis infection, and ALOX5 is believed to contribute to the TB onset, no research data for the Chinese population currently exist. In addition, because the previous ALOX5 TB association study was conducted on adults, research data are lacking for children. Hence, our study aimed to investigate if an association between the ALOX5 polymorphism and susceptibility to TB could be found in a subset of the pediatric population of northern China.
Materials and Methods
Ethics statement
The clinical investigation has been conducted according to the principles expressed in the Declaration of Helsinki. For research involving human participants, written informed consent has been written by the patients or the guardian of the patients. The research has been approved by the Ethics Committee of the Beijing Children's Hospital.
Study population
A total of 488 blood samples from 208 unrelated TB patients, 102 latent TB infection (LTBI) patients without active TB, and 178 healthy controls were obtained in this study. All study individuals were of Han Chinese ethnicity originating from the Beijing city and surrounding provinces in northern China. Study participants were admitted to the Beijing Children's Hospital (affiliated with the Capital Medical University, Beijing, China) between February 2005 and August 2010. All participants had been vaccinated with BCG at birth, as confirmed by the presence of a characteristic scar on the left shoulder and vaccination records.
All pediatric TB cases were diagnosed according to the diagnostic criteria of the Pediatric Tuberculosis Clinical Diagnosis Standard for China (Camila et al., 2007). The diagnostic criteria mainly includes positive clinical manifestations, imaging, a purified protein derivative (PPD) skin test, Acid-Fast Bacilli Stain, bacterial culture, fiberoptic bronchoscopy observation, cerebrospinal fluid findings, etc. In this study, primary immunodeficiency was ruled out according to the case history and clinical manifestations. None of the patients had histories of HIV infection, malnutrition, or conditions affecting the immune function, or were receiving immunosuppressive therapy or had any other type of lung disease. In addition, all TB cases were new; none had a previous history of TB or had received previous treatment for the disease.
In the LTBI group, all children with positive PPD skin test results ≥10 mm without the onset of active disease were diagnosed according to the diagnostic standards of the Pediatric Tuberculosis Clinical Diagnosis Standard for China (Hu et al., 2002); these cases were confirmed using the TSPOT test (an interferon-gamma release assay). Follow-up visits 1 year later showed that all of the LTBI patients that participated in our study did not develop TB disease.
The control group had normal radiographic examination findings, PPD skin test results <5 mm and did not have a history of TB or close contact with known active TB cases.
Laboratory methods, selection of tag-single-nucleotide polymorphisms, and genotyping
Following data release from Phase II of the International HapMap project (Frazer et al., 2007), the selection of tag-single-nucleotide polymorphism (SNPs) based on publicly available genotypes became a cost-effective option for researchers. Sample-based genotypes (data release 24) were downloaded for all variants in the gene regions and the 5000-bp promoter regions surrounding the ALOX5 gene (chr10:45189635-45261569).
First, since the study populations under investigation were from the Chinese population, downloaded genotypes were restricted to those for the Han Chinese in the Beijing, China (CHB) population. Tag-SNPs were selected using a pairwise tagging algorithm, and r2 exceeded 0.8 for all downloaded SNPs with a minor allele frequency >5% (de Bakker et al., 2005).
Second, we used an Illumina assay design score cutoff of 0.6 (scores are determined by the Illumina Assay Design Tool, and minimum recommendations are based on Illumina internal testing). SNPs were separated by more than 60 bp as recommended by the GoldenGate assay (Illumina, San Diego, CA).
Because the tag-SNP probabilities were discrete, accordingly, functional ranking of tag-SNPs with the same probability was used. Eighteen ALOX5 tag-SNPs were finally selected for genotyping (shown in Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/gtmb).
Blood samples from all the participants were collected and stored at −20°C. Genomic DNA was extracted from peripheral blood leukocytes according to standard methods. The present study used an Illumina BeadXpress Scanner for genotyping. This system uses the high-density BeadArray technology in combination with an allele-specific extension, adapter ligation, and amplification assay protocol. Briefly, biotinylated DNA was immobilized on paramagnetic beads and the pooled SNP-specific oligonucleotides were annealed onto the DNA. Oligonucleotides that hybridized were extended and ligated to generate DNA templates that were amplified using universal fluorescently labeled primers. Single-stranded PCR products were hybridized to a SentrixH Array Matrix. Genotypic analysis was performed using the GoldenGate Assay Kit. Oligonucleotides were amplified by allele-specific primer extension. After Sentrix Array Matrix hybridization, signal intensities were read by a BeadArray Reader (Illumina). BeadStudio III software (Illumina) was used for clustering the genotype data. Twenty random duplicated DNA samples were genotyped twice to provide replicate data to review and refine the clustering. For quality control, only the samples and marker call rates >95% were used. BeadStudio genotype data were transferred electronically to Statistical Package for Social Sciences version 13.0 (SPSS) for further analysis.
Statistical analysis
Statistical analyses were carried out using SPSS 13.0. Differences between genotype distributions and allele frequencies were tested by chi-square analysis. Significant differences between or among groups were indicated by values of pb<2.8*10−3. Haploview (Barrett et al., 2005) was employed to visualize linkage disequilibrium (the D’ statistic) and haplotype blocks between these variants.
Results
Patients and healthy controls (noninfected controls)
The study subjects included 208 patients with TB, 102 latent TB infection (LTBI) subjects, and 178 ethnically matched noninfected controls representing a subset of the Han Chinese pediatric population of northern China. The study participants were matched for sex and age. The mean age was 5.8 years (SD 4.7; range, 2 months-16 years) for TB patients, 7.1 years (SD 4.7; range, 3 months-15 years) for LTBI subjects, and 5.5 years (SD 4.2; range, 3 months-15 years) for the control subjects. The proportion of male subjects was 129/208 (62.0%) in the TB patient group, 58/102(56.9%) in the LTBI group, and 101/178 (56.7%) in the control group. Detailed clinical information is listed in Table 1.
TB, tuberculosis; LIBI, latent TB infection; CSF, cerebrospinal fluid; FOB, fiberoptic bronchoscopy observation; PPD, purified protein derivative.
Tag-SNPs, tagged-haplotype, and linkage disequilibrium analysis
Eighteen tag-SNPs within ALOX5 were selected using the above criteria for picking tag-SNPs (Methods); 17 of them were located within the intron region, while rs2228064 existed at exon 2 to be a synonymous (p. T90T) SNP (Fig. 1). All of the tag-SNPs could be multiplexed in a single well for genotyping.
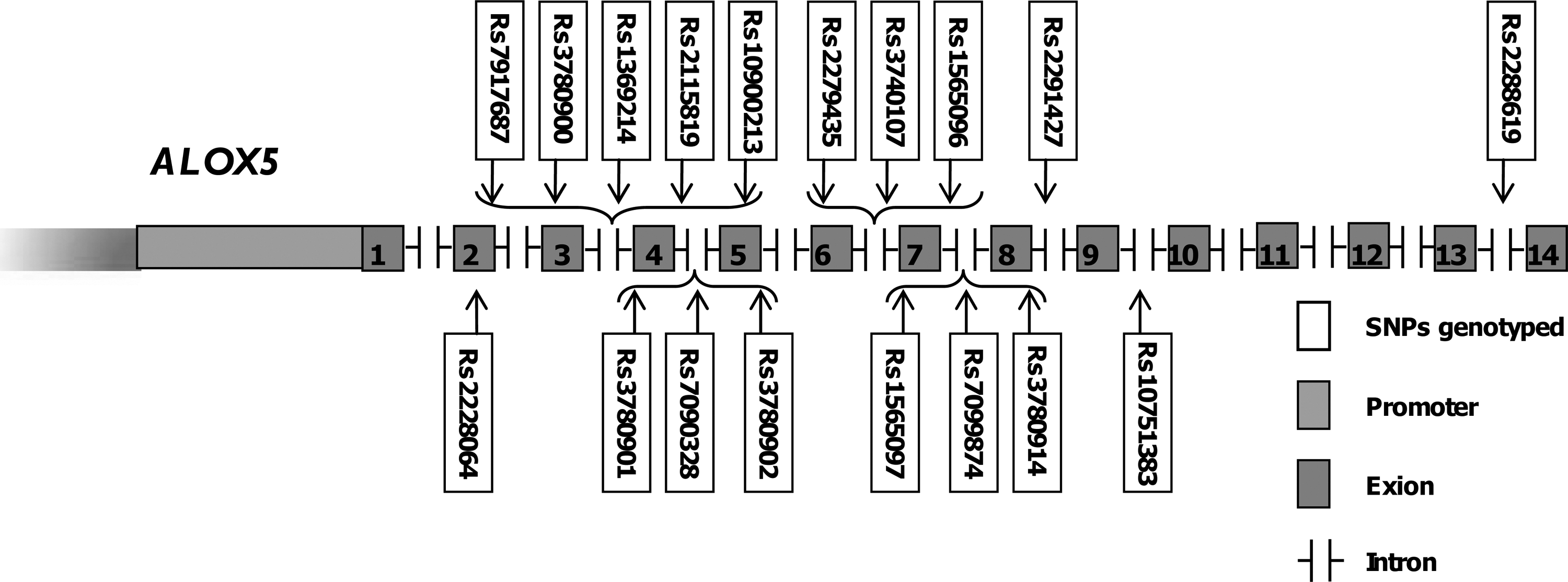
The gene structure of ALOX5 and genetic positions of the 18 tag-single-nucleotide polymorphisms genotyped in ALOX5.
According to the results, the average call rate for the tag-SNPs was 99.2% overall and 99.3% among the study participants. Four tag-SNPs (rs7917687, rs2279435, rs1565096, and rs2291427) were not in the Hardy-Weinberg equilibrium (HWE) in the control group, so were excluded from further analysis. The same linkage disequilibrium map was detected in the control group when compared with the CHB population in the HapMap database (Supplementary Fig. S1).
Allele and genotype analysis
The allele frequencies and genotypic distributions of the tag-SNPs are shown in Supplementary Table S2. Considering multiple-site detection was carried out in this experiment, a Bonferroni-corrected p-value for each site was used in our analysis. Out of the 14 tag-SNPs in HWE in the control group, one tag-SNP showed obvious differences between the TB and control groups, and also between the TB and LTBI groups (p<0.05, listed in Table 2). The frequency of the rs2115819 T allele is 51.0% in the TB group, 31.6% in the control group, and 27.2% in the LTBI group. The rs2115819 T allele appeared to be one TB risk factor in that the frequency was significantly higher in the TB group compared to both control groups (Corrected p=1.2e-06, odds ratio [OR]=2.22, 95%confidence interval [CI]=[1.67-3.03]) and the LTBI group (Corrected p=1.56e-06, OR=2.78, 95%CI=[1.89-4.00]).
SNP, single-nucleotide polymorphism; CI, confidence interval; OR, odds ratio.
Discussion
ALOX5 mRNA is 2568 base pairs in length, coding for a 674 aa protein called 5-lipoxygenase (5-LO). 5-LO is involved in inflammatory and immune disease, it is a key enzyme that catalyzes reactions in the formation of two eicosanoids: LTs and LXs (Matsumoto et al., 1988; Voelkel et al., 1996; Luo et al., 2003). A gene mutation of ALOX5 reduces the synthesis of LTs and LXs. When wild-type mice were infected with M. tuberculosis, high levels of LTB4 and LXA4 were produced by endothelial cells and macrophages. As early as 1 week after M. tuberculosis aerosol infection (300 CFU/animal), LTB4 and LXA4 increased in the sera of the experimental mice (B6, 129S F2/J). Synthesis of LXA4 was maintained during chronic TB infection. In mice lacking ALOX5 expression (5-LO−/− mice), TB infection resulted in a fewer bacilli and a lower inflammatory infiltration than in wild-type animals, and in this study, no evaluated LTB4 or LXA4 was detected postinfection (Bafica et al., 2005). With high-dose challenge infections (but not low-dose), however, the normally resistant wild-type mouse strain succumbed more rapidly to infection than did the 5-LO−/− mice. This indicates that in the 5-LO-deficient mice (that exhibited increased resistance to M. tuberculosis infection), ALOX5 is a strong candidate gene for susceptibility to TB infection.
M. tuberculosis activates the 5-LO pathway not only to inhibit apoptosis, but also to prevent cross presentation of its antigens by dendritic cells, which could block T cell triggered immunity (Divangahi et al., 2010). In 5-LO−/− mice (compared with wild-type mice), increased expression of interleukin-12 and interferon-γ were detected (through real-time reverse transcription-polymerase chain reaction analysis) (Bafica et al., 2005). 5-LO deficiency is associated with a reduced matrix metalloproteinase-2 activity and diminished plasma macrophage inflammatory protein-1-alpha (CCL3) (Zhao et al., 2004). As these cytokine/chemokines are involved in the pathogenesis of TB (Zhu and Friedland, 2006; Aly et al., 2007; Lee and Kornfeld, 2010; Yim and Selvaraj, 2010; Green et al., 2011), 5-LO could influence the survival of M. tuberculosis through a potential cytokine/chemokine intermediary route.
In 2008, Herb et al. (2008) genotyped ALOX5 polymorphisms in 1916 sputum-positive adult patients with pulmonary TB and 2269 exposed apparently healthy controls from the Ghanaian population. After analyzing polymorphisms for a variable number of tandem repeats of the ALOX5 promoter and of the exonic nonsynonymous variant g.760G>A, an association of ALOX5 variants with susceptibility to TB was detected. Unlike the previous study by Herb, we hereby investigate tag-SNPs that represent the entire ALOX5 gene region to explain the association of ALOX5 with susceptibility to TB disease in a pediatric population in North China. Compared with other research groups, the present association study using the 18 selected tag-SNPs may reflect the relationship of ALOX5 and TB disease more comprehensively. And, besides TB cases and healthy controls, we also analyzed LTBI samples that we believed would provide more robust data for assessing the association of ALOX5 with the onset of TB disease from M. tuberculosis infection.
By our present study, the rs2115819 tag-SNP T allele seemed to be a risk factor for TB. Significant differences were found between the TB patients group and the control group and also between the TB group and the LTBI group. We noticed that the rs2115819 risk T allele was less frequent in the LTBI group than in the control group. This may be because the LTBI samples used in our experiment are more resistant to TB disease, for although having been diagnosed to be infected by M. tuberculosis, they had not progressed to the onset of TB disease even 1 year latter. This is the first time that ALOX5 has been shown to be associated with pediatric TB, not only in the TB-control study, but also in the TB-LTBI study.
Although ALOX5 has been found to be associated with adult TB in the Ghanaian population, ALOX5 association study results have not been reported for any other population. Crucially, there is a complete lack of data for pediatric groups. Children represent a unique risk group for TB, because their progression from the onset of infection to active disease is usually rapid. TB disease in young children reflects recent infection rather than secondary reactivation. The progression from TB infection to the disease onset is rapid, usually less than 1 year (Newton et al., 2008; Rigouts, 2009). Thus, pediatric TB is clinically and pathologically different from adult TB. Childhood TB is not only an important public health issue, but a matter of sensitivity for those families affected by the disease.
One weakness of our study is the lack of association data of genetic polymorphisms with ALOX5 expression and 5-LO activities. As we know genetic variations should contribute to altered expression levels. Although rs2115819 are tag-SNPs existing in intron 3, they could also potentially regulate gene expression by interacting with transcriptional regulation factors or by influencing the mRNA splicing process. Clearly, further investigations are now needed to discover whether the different ALOX5 genetic variants are associated with changes in ALOX5 expression levels.
Conclusion
Although 5-LO-dependent LT and LX synthesis could play a major role in the immune modulation of M. tuberculosis infection, and ALOX5 is believed to contribute to the TB onset, research data were lacking for the Chinese population. In addition, the previous ALOX5 association study conducted on adults provided no data for children. By investigating if polymorphisms in ALOX5 are associated with susceptibility to TB disease in a subset of the pediatric population of northern China, we have shown the rs2115819 T allele of ALOX5 to present a risk factor for childhood TB disease.
Footnotes
Acknowledgments
The authors wish to thank all of the subjects for their ongoing participation in this study. This study was supported by grants from the National Natural Science Foundation of China (No. 81071315 and No. 81201263), Beijing Natural Science Foundation (No. 7121007), a grant from the Ministry of Science and Technology (No. 2011ZX09302), and the National Science & Technology Pillar Program of China (No. 2008BAH24B05).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.