
Abstract
Aims: To assess the relationship between mutations of the myocyte enhancer factor 2A (MEF2A) and premature coronary artery disease (PCAD) in a Chinese population. Methods and Results: The mutations in the exons 8 and 12 of the MEF2A gene were analyzed in both PCAD families and sporadic cases using direct sequencing of polymerase chain reaction products. In one PCAD family, seven members of the third generation were all diagnosed with CAD, and five of them had PCAD. All five members with PCAD displayed a mutation of the TT genotype in the site of 1353 G/T. Moreover, three of them (3/5) had a mutation of the DW genotype in the site of 1291-1293 CCG W/D. In sporadic cases, we also found that the haplotype of 1291-1293 CCG D+1305 G+1353 T was significantly associated with PCAD. Conclusions: The mutations of MEF2A exon 12 are implicated in PCAD, suggesting a strong genetic component in the pathogenesis of PCAD in the Chinese population.
Introduction
Methods
Selection of PCAD family and sporadic cases
All study subjects were chosen from the patients treated in the Peking Union Medical College Hospital (PUMCH), including 55 sporadic PCAD patients, 56 CAD (non-PCAD) patients, and 65 non-CAD subjects. CAD was diagnosed based on the angiographic criteria as stenosis ≥50% of the lumen diameter in a major coronary artery or its major branches. The selection criterion of PCAD was defined as CAD diagnosed before 55 years of age in men and before 65 years of age in women. The exclusion criteria included chest pain caused by inflammation, infection, heart failure, tumor, and other nonischemic reasons. The selection criteria of the PCAD family for the study were defined as more than 10 members in the family and more than five members with PCAD. A typical pedigree of the disease penetration in one family with PCAD is illustrated in Figure 1, and the clinical condition of each family member is summarized in Table 1.
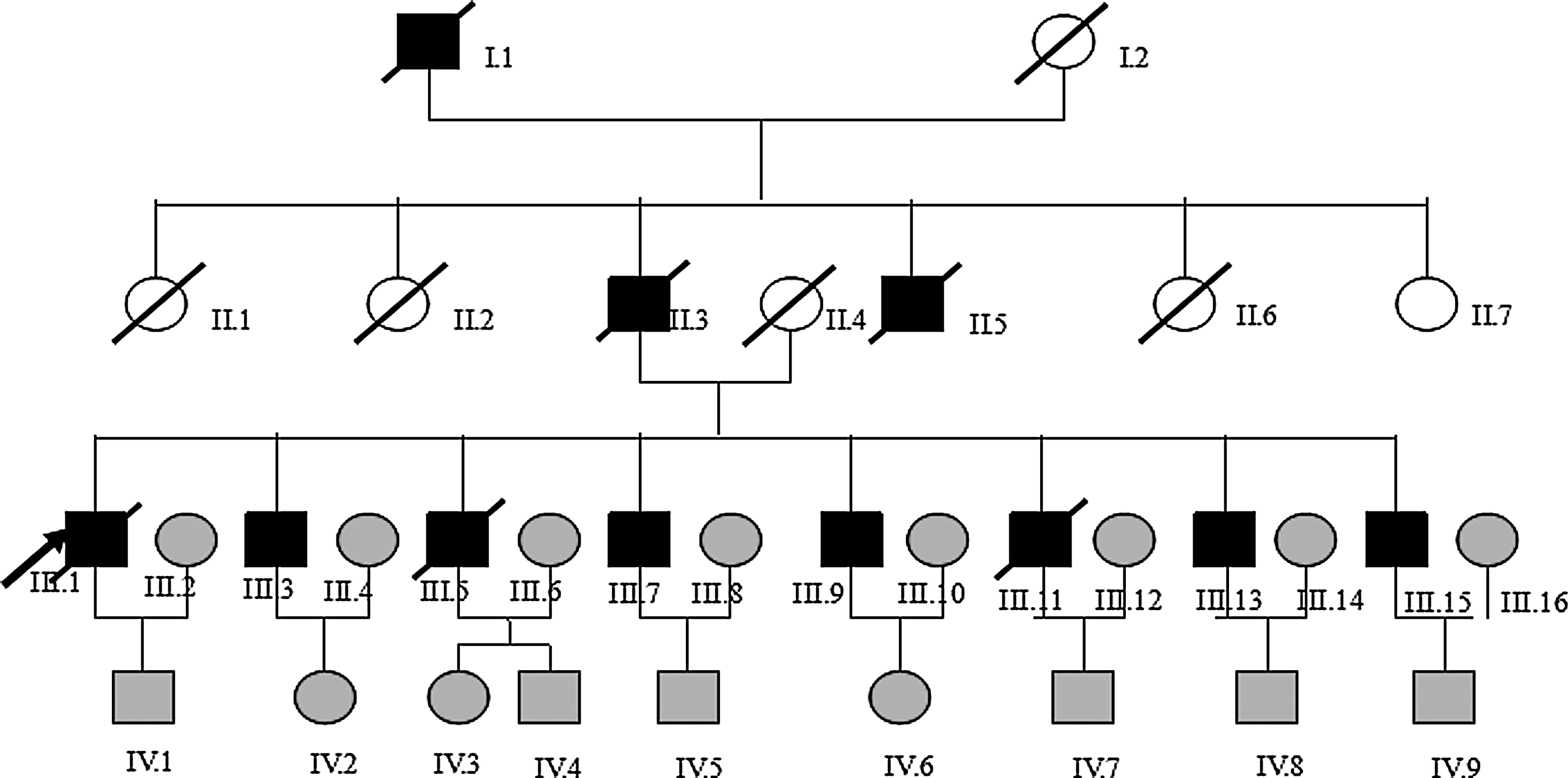
Pedigree structure and genotypic analysis of a family with premature coronary artery disease. Individuals with coronary artery disease were indicated by solid squares (men) or solid circles (women). Unaffected individuals were indicated by open symbols. Men under the age of 55 or women under 65 years of age without symptoms were shown in light gray as an uncertain phenotype. Deceased individuals were indicated by a slash (/). The proband was indicated by an arrow.
The members of uncertain phenotype in the third and fourth generations are not shown in the table.
MI, myocardial infarction; PCI, percutaneous coronary intervention; CABG, coronary artery bypass graft; PCAD, premature coronary artery disease.
All subjects enrolled in this study were Chinese with Han ethnicity background. They were all appropriately consented for their participation in the study, and the study was approved by the Committee of Ethics in PUMCH.
Sequencing analysis of MEF2A exon 8 and 12
DNA was obtained from all subjects following the phenol-chloroform isolation method. The exons 8 and 12 of the MEF2A gene were amplified by polymerase chain reaction (PCR) with specific primers (Exon 8 sense: GTT TGT GCC AAA GTA TTT TAA; exon 8 antisense: AAG AAT GAAGTT GAA GAA AGG; exon 12 sense: TGC AGA GGT ACT TGC AAG CCA T; exon 12 antisense: AGA TAT GTA GGG CAG GTC ACT). Each reaction consisted of 80 ng genomic DNA, 10 pmol primers, 2.5 mM MgCl2, and 2 mM of each dNTP in a final volume of 50 μL. The PCR products were amplified for 30 cycles under the following condition: denaturation at 95°C for 30 s, annealing at 58°C (exon 8) or 64°C (exon 12) for 30 s, extension at 72°C for 30 s with final extension time of 7 min at 72°C. The initial denaturation stage was carried out at 95°C for 5 min. Then, the products of PCR were purified and sequenced with an automated ABI 3730 system.
Statistical analysis
Statistical analysis was carried out using the SPSS v13.0 for Windows statistical package (SPSS, Chicago, IL). A p-value of 0.05 or less was considered significant. The Hardy-Weinberg equilibrium for genotype distribution was estimated by the chi-square test. The linkage disequilibrium analysis for alleles between different groups and the risk estimation of different haplotypes were performed by the UNPHASED software after a pedigree database had been set up.
Results
There was no polymorphism detected in MEF2A exon 8. However, five polymorphism sites were identified in MEF2A exon 12 of sporadic PCAD patients and their family members. One CAD patient displayed a novel 1303 C/T mutation. Further study is needed to define its relevance and significance in CAD.
In the PCAD family (Table 2), all of the third generation who survived from PCAD had a genotype of TT at the 1353 G/T polymorphism site. In addition, three of them had the genotype of DW at the 1291-1293 CCG/− site. Therefore, we postulate that the combined mutations of 1353 G/T and 1291-1293 CCG/− sites may be closely related to the onsets of PCAD in many members in this family.
The five members in the third generation are shown in bold.
In sporadic cases, the 1291-1293 CCG/−, 1305 G/A, and 1353 G/T polymorphisms in the PCAD, non-PCAD, and non-CAD groups were in the Hardy-Weinberg equilibrium. Using a case-control study, we found that the CAG repeat number at site 1 and the 1305 G/A polymorphism at site 4 (Table 3) were not related to PCAD or CAD susceptibility. However, the DD/DW genotype and the D allele at the site of 1291-1293 CCG/− and the TT genotype and the T allele at the site of 1353 G/T were strongly associated with CAD and PCAD in particular. There was a strong linkage disequilibrium in the distribution of polymorphisms 1291-1293 CCG/−, 1305 G/A, and 1353 G/T among different groups.
A novel mutation of 1303 C→T was found in only one case with CAD in all subjects.
STR, short tandem repeat; SNP, single-nucleotide polymorphism; MEF2A, myocyte enhancer factor 2A.
The haplotype frequencies defined by three polymorphism sites (including 1291-1293 CCG/−, 1305 G/A, and 1353 G/T) are shown in Table 4. Compared with the non-PCAD and non-CAD groups, the frequency of haplotype 1291-1293 CCG D and 1305 G and 1353 T was markedly higher in PCAD.
The shaded box marked the haplotype with statistically significant difference compared with the wild haplotype (p<0.05). Haplotypes were only shown with frequency ≥3%.
Discussions
MEF2A belongs to a family of four closely related transcription factors (MEF2A to MEF2D). The MEF2 factors act as a homo- or heterodimer, and each of these proteins binds to the MEF2 target DNA sequence (the A-T-rich sequence) present in the regulatory regions of many downstream genes related to vascular development and controls transcription. MEF2A is expressed in endothelial and smooth muscle cells of coronary arteries, and MEF2A mutations could disrupt the growth or differentiation of these cells, increasing the risk of developing coronary artery disease among mutation carriers.
In 2003, Wang et al. first reported a MEF2A mutation associated with CAD/MI in a large family with several affected members. The mutation in MEF2A exon 12 resulted in a seven-amino acid deletion of the protein that was important for nuclear localization. However, the exact role of MEF2A mutations and polymorphisms in CAD remains to be fully studied (Wang et al., 2005a; Weng et al., 2005b). In addition, the mutation frequency of MEF2A may vary greatly in different races, regions, and populations. In our study, we found that the frequency of combined mutations of 1291-1293 CCG/− and 1353 G/T was higher in a family with apparent inheritable PCAD (Figure 2). This is consistent with the findings of a subsequent study in sporadic cases. Therefore, this study suggests that the haplotype of 1291-1293 CCG D and 1305 G and 1353 T is a genetic risk factor of PCAD in the Chinese Han population.
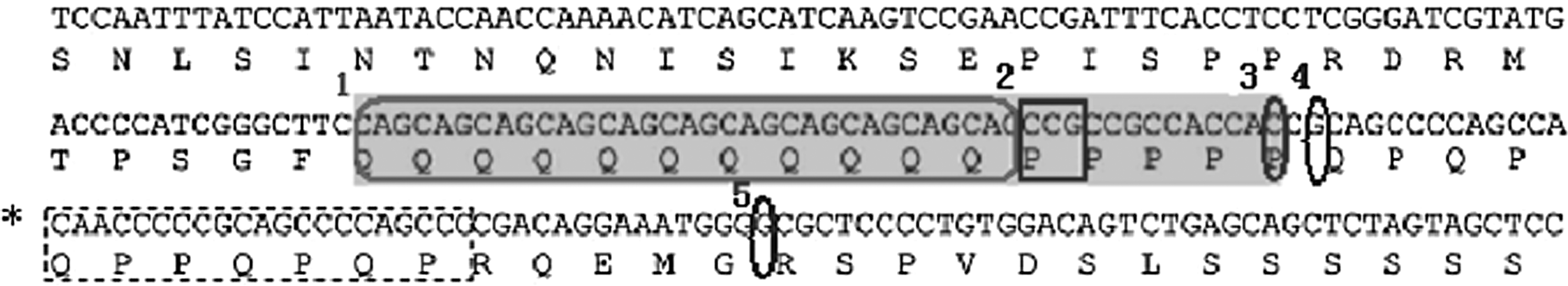
MEF2A contains a MADS domain and a MEF2-specific domain at the N terminus. The MADS domain mediates protein dimerization and binding to A-T-rich DNA sequences, while the adjacent MEF2 domain is required for high-affinity DNA binding, dimerization, and interacting with cofactors. The C terminus plays an important role in transcription activation and nuclear localization (Black et al., 1998; McKinsey et al., 2002). Therefore, the effects of the mutations discussed above in this study lying in the C terminus of MEF2A were probably to change the transcription activity of many downstream target genes and influence the normal function of vascular endothelium cells. This might be a potential molecular mechanism that would explain the pathogenesis of PCAD, either familial or sporadic.
In conclusion, CAD is a multifactorial disease caused by environmental and genetic factors. Many genes and mutations might be involved in the genesis of the disease. So, the analysis based on haplotypes can be advantageous over an analysis based on individual mutations in the presence of multiple susceptibility alleles and provides more information. The statistical model based on haplotypes analysis in this study can be further used to identify and locate complex gene mutations in PCAD.
Footnotes
Acknowledgments
This study was supported by the Science Research Foundation of the Peking Union Medical College Hospital.
Author Disclosure Statement
No competing financial interests exist.