
Abstract
Objective: Several studies have examined the association between the GCK -30G>A polymorphism and the risk of gestational diabetes mellitus (GDM) and type 2 diabetes mellitus (T2DM). However, inferences from these studies are hindered by their limited statistical power and conflicting results. The aim of this meta-analysis is to provide a relatively comprehensive picture of the association of the GCK -30G>A polymorphism with GDM and T2DM risk. Methods: A literature search for eligible studies published before August 15, 2013, was conducted in PubMed, Embase, Web of Science, Cochrane Library, and CNKI (China National Knowledge Infrastructure). Pooled odds ratios with their corresponding 95% confidence intervals were used to evaluate the strength of the association under a fixed- or random-effect model according to heterogeneity test results. All analyses were performed using Stata software, version 12.0. Results: Eighteen case-control studies from 17 published reports were included in this meta-analysis with a total of 2011 patients with GDM, 11,057 with T2DM, and 26,102 healthy controls. For GDM, the combined results showed that the risk allele of the -30G>A polymorphism may be associated with an increased risk of GDM. Stratified analyses showed that the magnitude of the effect was especially significant among whites, indicating ethnicity differences for GDM susceptibility. For T2DM, the pooled ORs were not significant in the overall population, although all the ORs >1 suggested an increased risk of T2DM for carriers of the A allele. However, whites seem to be significantly more susceptible to T2DM than Asians. Conclusion: This meta-analysis indicated that the risk allele of the GCK -30G>A polymorphism may increase GDM and T2DM risk in whites, whereas additional studies are needed to confirm the effect of this polymorphism on both diseases in Asians and Africans.
Introduction
G
Because women with a history of GDM are at an increased risk of developing T2DM later in their lives and women with a family history of T2DM may be predisposed to an increased risk of GDM (Kim et al., 2002), it is plausible to hypothesize that GDM may share the same risk factors and genetic susceptibilities as T2DM. Over the past few decades, results of candidate gene studies, giving support to this hypothesis, showed that frequencies of some alleles associated with an increased risk of developing T2DM were also increased in women who had GDM (Megia et al., 2004; Shaat et al., 2005).
Glucokinase (GCK), encoded by the GCK (or MODY2) gene on chromosome 7p, is thought to be part of the glucose-sensing mechanism for the synthesis of proinsulin and the release of insulin (Froguel et al., 1992). Because of its role in the regulation of insulin secretion, the GCK gene is an attractive candidate gene for both GDM and T2DM risk. Several variants in the GCK gene have been identified, with the -30G>A (rs1799884) variant being the most extensively examined in GDM and T2DM association studies (Chiu et al., 1994; Zaidi et al., 1997; Shaat et al., 2006; Freathy et al., 2010). The variant G to A at position −30 might lead to a decrease in GCK activity and to an increase in the threshold for glucose-stimulated insulin secretion (Rissanen et al., 1998). Because this variant is located within a highly conserved region of the GCK promoter, it might alter transcriptional regulation of the gene (Rose et al., 2005). If this variant reduced the transcription of the GCK gene, the cellular activity of GCK would likely decrease and lead to impaired glucose sensing, insulin secretion of β cells and eventually diabetes (Murad et al., 2010). To date, many case-control studies have investigated the role of the GCK -30G>A polymorphism in the development of GDM and T2DM among various populations. However, results are still conflicting, and the molecular mechanism by which the -30G>A polymorphism influences an individual's susceptibility to GDM and T2DM is as yet unclear. Hence, we performed a meta-analysis to derive a relatively comprehensive picture of the relationship between the GCK -30G>A polymorphism and the risk of GDM and T2DM.
Materials and Methods
Search strategy
A literature search for eligible studies published before August 15, 2013, was conducted in the following: PubMed, Embase, Web of Science, Cochrane Library, and CNKI (China National Knowledge Infrastructure). The following combined Medical Subject Heading terms were used: (“glucokinase” or “GCK”) and (“gestational diabetes” or “GDM” or “type 2 diabetes mellitus” or “T2DM”) and (“genetic polymorphism” or “SNP”). The search was done without any limitation on language but included only studies that were conducted on humans.
Selection criteria
To be included in the analysis, candidate studies had to meet the following criteria: (1) case-control study focused on the relationship between the GCK -30G>A polymorphism and GDM or T2DM risk, (2) all patients meeting the diagnostic criteria for GDM or T2DM, and (3) sufficient original data for calculating odds ratios (ORs) with corresponding 95% confidence intervals (CIs). The major reasons for excluding studies were design other than a case-control study, duplicate publications, and no available data reported.
Data extraction
Two independent reviewers checked each full-text report for eligibility, then extracted and tabulated the following data from eligible studies: surname of first author, year of publication, country of origin, ethnicity, definition and numbers of cases and controls, age, body mass index, diagnostic criteria, genotyping method, allele and genotype frequency, and Hardy-Weinberg equilibrium (HWE) status in controls. Disagreements were solved by discussion with all authors until consensus was reached.
Quality assessment
The Strengthening the Reporting of Genetic Association Studies (STREGA) quality score system and the Newcastle-Ottawa Scale (NOS) criteria were used to assess the qualities of all included studies (Little et al., 2009; Stang 2010). The STREGA system includes 22 assessment items related to quality appraisal with scores ranging from 0 to 22. The NOS criteria use a “star” rating system to judge methodologic quality based on three aspects of a study: selection, comparability, and exposure. Two authors assessed the quality of included studies independently. Discrepancies on the quality scores were solved by discussions and subsequent consensus.
Statistical analysis
With consideration of possible between-study heterogeneity, a statistical test for heterogeneity was conducted by using Cochran Q statistic, with p<0.05 indicating significant between-study heterogeneity (Jackson et al., 2012). Heterogeneity was also assessed through the I2 test, with I2>50% indicating significant heterogeneity (Higgins and Thompson, 2002). For p<0.05 or I2>50%, a fixed-effect model was used to estimate the pooled ORs with their corresponding 95% CIs. Otherwise, a random-effects model was applied. The significance of the pooled ORs was determined by using the Z test. Genotype distributions in the controls were tested for HWE status with use of the chi-square test. Prespecified stratified analyses were performed to explain heterogeneity and to investigate whether the reported association was present in subgroups based on ethnicity, diagnostic criteria, and genotyping method where applicable. Sensitivity analyses were performed to assess the stability of the pooled results. The Begg's funnel plot and Egger linear regression test were used to assess the potential for publication bias (Peters et al., 2006). All two-tailed p values<0.05 were considered to represent statistically significant differences. All analyses were performed by using Stata software, version 12.0 (Stata Corp, College Station, TX).
Results
Baseline characteristics of included studies
A total of 212 relevant papers were identified by using the prespecified search strategy. In accordance with the inclusion criteria, 18 case-control studies from 17 published reports met our inclusion criteria (Chiu et al., 1994; Shimokawa et al., 1994; Stone et al., 1996; Lotfi et al., 1997; Yamada et al., 1997; Zaidi et al., 1997; Rissanen et al., 1998; Rose et al., 2005; Shaat et al., 2006; Cauchi et al., 2008; Ezzidi et al., 2009; Ma and Wang, 2009; Freathy et al., 2010; Hu et al., 2010; Murad et al., 2010; Santos et al., 2010; Tam et al., 2010), with 6 focusing on GDM and 12 on T2DM. A flow chart of the study selection process and the specific reasons for exclusion from this meta-analysis are shown in Figure 1. A total of 39,170 patients were involved in this meta-analysis, including 2011 with GDM, 11,057 with T2DM, and 26,102 healthy controls. Publication years of the included studies ranged from 1994 to 2010. The distribution of genotypes in the controls was consistent with HWE in all studies (all p>0.05). The quality scores of all the included studies were moderate-high, with STREGA scores>13 and NOS stars>5. The characteristics and methodologic quality of the included studies are summarized in Table 1.
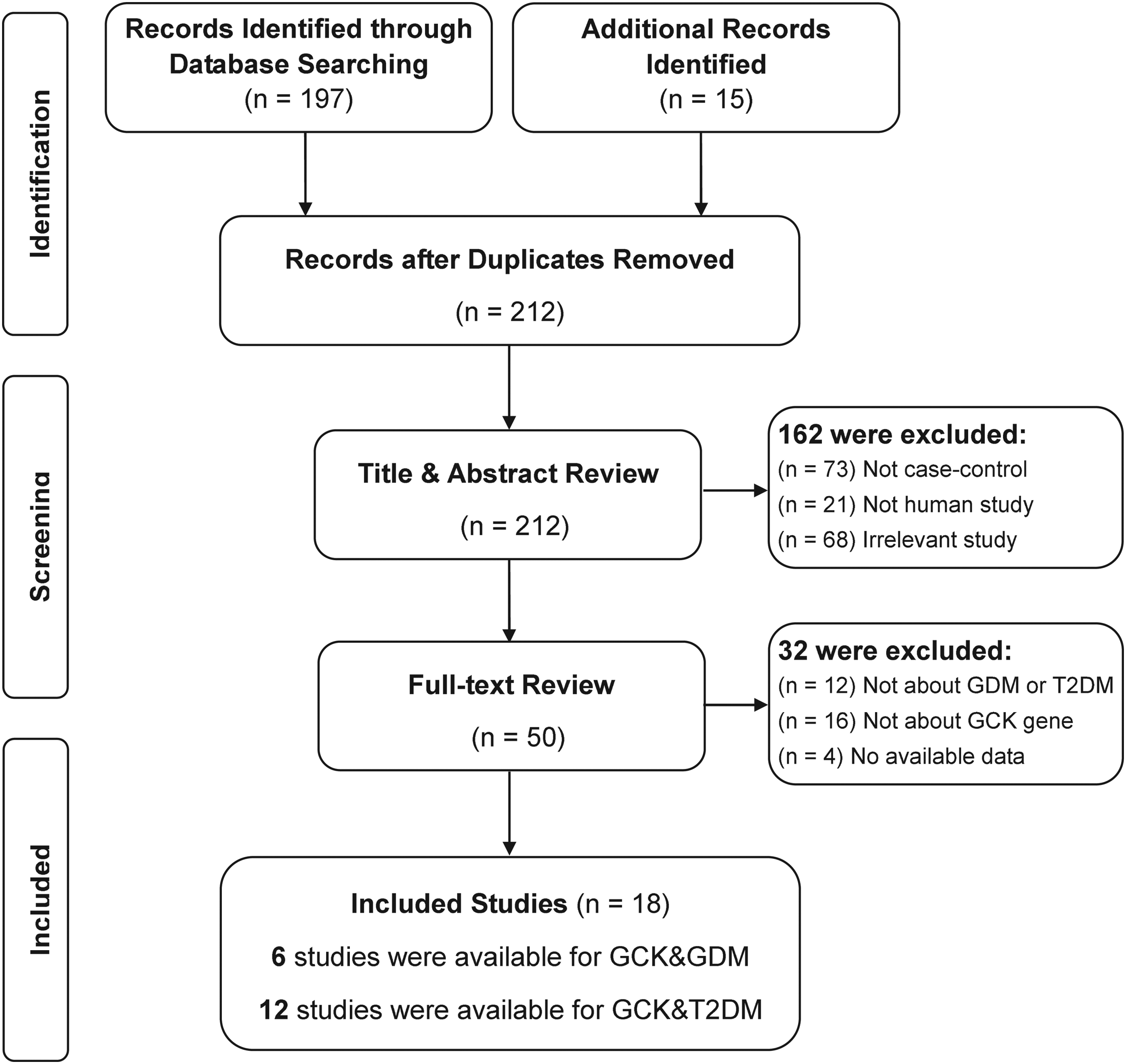
Flow diagram of the selection of studies and specific reasons for exclusion from the present meta-analysis. GDM, gestational diabetes mellitus; T2DM, type 2 diabetes mellitus.
ADA, American Diabetes Association; BMI, body mass index; DPSG-EASD, Diabetic Pregnancy Study Group of the European Association for the Study of Diabetes; GDM, gestational diabetes mellitus; IADPSG, new International Association of Diabetes and Pregnancy Study Groups; NA, not available; NOS, Newcastle-Ottawa Scale; PCR-RFLP, polymerase chain reaction restriction fragment-length polymorphism; PCR-SSCP, polymerase chain reaction-single strand conformation polymorphism; SD, standard deviation; STREGA, Strengthening the Reporting of Genetic Association Studies; T2DM, type 2 diabetes mellitus; UK, United Kingdom; USA, United States; WHO, World Health Organization.
Association between GCK -30G>A polymorphism and GDM risk
Table 2 evaluates the association between the GCK -30G>A polymorphism and GDM risk. Six case-control studies investigated the relationship between -30G>A and GDM risk with a total of 2011 GDM cases and 7490 healthy controls. The fixed-effects model was conducted because no heterogeneity obviously existed (all p>0.05 and I2<50% under five genetic models). The combined results indicated that the A allele of the −30 G>A polymorphism was significantly associated with increased risk of GDM under all genetic models (A versus G: OR, 1.25 [95% CI, 1.13-1.37; p<0.001]; GA+AA versus GG: OR, 1.25 [95% CI, 1.12-1.40; p<0.001]; AA versus GG+GA: OR, 1.64 [95% CI, 1.24-2.17; p=0.001]; AA versus GG: OR, 1.50 [95% CI, 1.06-2.11; p<0.001]; AA versus GA: OR, 1.44 [95% CI, 1.07-1.92; p=0.015]).
CI, confidence interval; OR, odds ratio.
Stratified analysis by ethnicity showed that individuals with the A allele had increased GDM risk compared with people with the G allele in white and Asian, but not in African, populations, indicating ethnic variation in susceptibility to GDM (Fig. 2). Although GDM risk was significantly increased in Asians under allele and dominant models (A versus G: OR, 1.31 [95% CI, 1.04-1.67; p=0.021]; GA+AA versus GG: OR, 1.39 [95% CI, 1.07-1.80]; p=0.014), the small sample size in only one included study may limit the statistical power of this result. Subsidiary analyses of the criteria used for diagnosing GDM showed that Diabetic Pregnancy Study Group of the European Association for the Study of Diabetes and International Association of the Diabetes and Pregnancy Study Groups subgroups had a significantly higher risk of GDM than the other two subgroups (Table 2). In addition, significant associations were observed for polymerase chain reaction-single-strand conformation polymorphism and Illumina subgroups based on genotype method, but not in the polymerase chain reaction-restriction fragment-length polymorphism subgroup (Table 2).
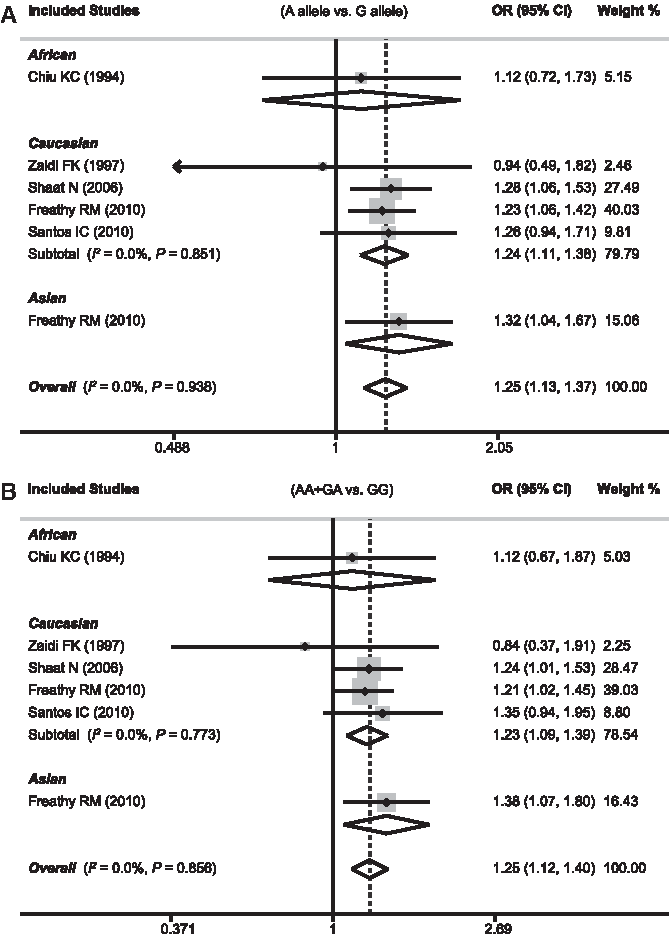
Forest plots of odds ratios for the association between the GCK -30G>A polymorphism and susceptibility to gestational diabetes mellitus in subgroup analysis based on ethnicity under allele model
Association between GCK -30G>A P polymorphism and T2DM risk
Table 3 summarizes findings on the relationship between the GCK -30G>A polymorphism and susceptibility to T2DM. Data from 12 case-control studies (compromising 11,057 T2DM cases and 18,612 healthy controls) were pooled together for analysis. Because heterogeneity obviously existed (all p<0.05 and I2>50% under five genetic models), the random-effects model was used. The pooled ORs were not significant in the overall population, although all the ORs were >1, suggesting an increased risk of T2DM for carriers of the A allele. However, stratified analysis by ethnicity suggested that whites seemed to be more susceptible to T2DM than Asians, with the ORs significantly >1 (Fig. 3). A significant association was also detected in Africans, but the statistical power of this result may be limited by a small sample size because only one study of this polymorphism has been conducted in an African population. In addition, there is an obvious contradiction in the implication of a result with all ORs under five genetic models <1, which indicated the opposite conclusion compared with the overall effect (Table 3). Furthermore, stratified analysis based on diagnosis criteria and genotyping method found no significant results for any of the six subgroups.
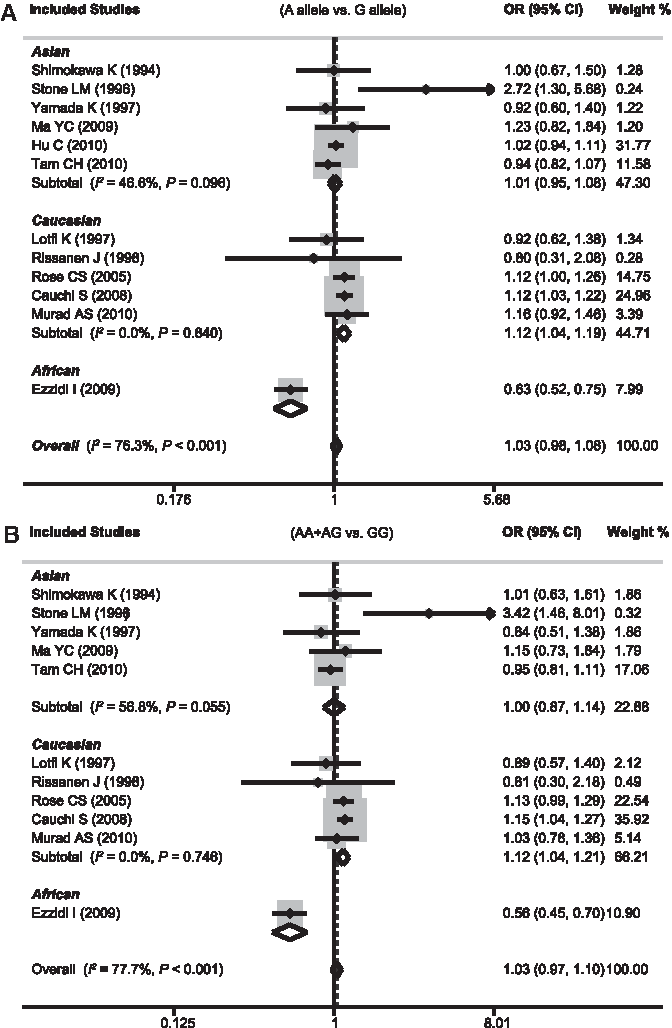
Forest plots of odds ratios for the association between the GCK -30G>A polymorphism and susceptibility to type 2 diabetes mellitus in subgroup analysis based on ethnicity under allele model
Sensitivity analyses and publication bias
Sensitivity analyses of both GDM and T2DM susceptibility indicated that no single study significantly influenced the pooled ORs (Fig. 4). The shapes of the Begg's funnel plots did not reveal any evidence of obvious asymmetry (Fig. 5). In addition, we did not find any evidence of publication bias from the Egger linear regression test (GCK -30G>A and GDM: t=(1.53, p=0.200; GCK -30G>A and T2DM: t=0.45, p=0.664). These tests indicated a promising level of robustness and accuracy for the results of this meta-analysis.
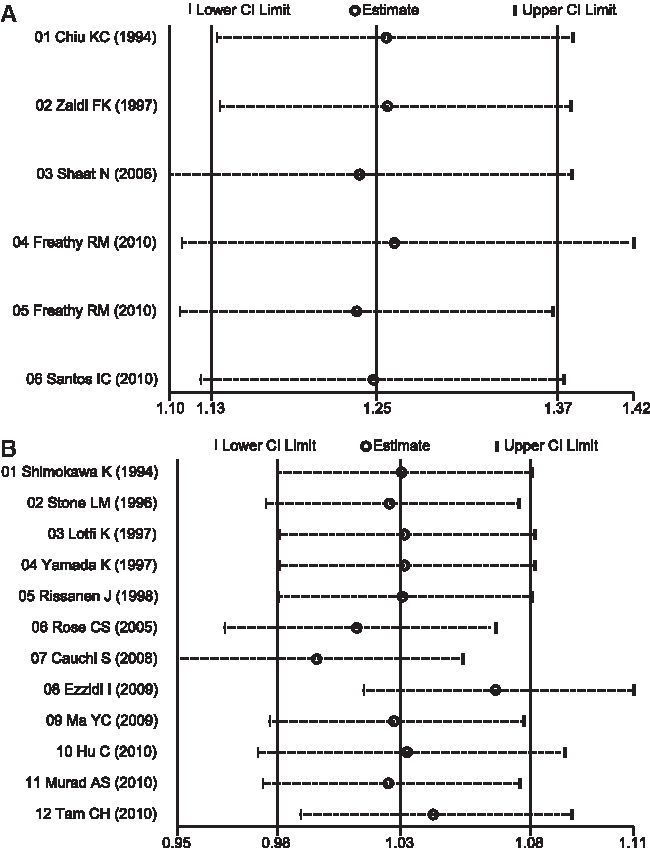
Sensitivity analysis of the association between the GCK -30G>A polymorphism and susceptibility to gestational diabetes mellitus

Begg's funnel plots of publication bias for the association between the GCK -30G>A polymorphism and susceptibility to gestational diabetes mellitus
Discussion
GDM shares many risk factors with T2DM, and up to 50% of women with GDM develop T2DM within 10 years after pregnancy (Ben-Haroush et al., 2004). However, detailed pathogenetic mechanisms of GDM and T2DM remain a matter of speculation. GCK is the key glucose phosphorylation enzyme responsible for the first rate-limiting step in the glycolysis pathway and regulates glucose-stimulated insulin secretion from pancreatic β-cells and glucose metabolism in the liver (Matschinsky 1990). Inactivating GCK mutations lead to maturity-onset diabetes of the young and neonatal diabetes (Njolstad et al., 2001), whereas activating GCK mutations cause persistent hyperinsulinemia and hypoglycemia (Christesen et al., 2002; Gloyn et al., 2003). Thus, the GCK gene seems to play a key role in glucose homeostasis and is a determinant of diabetes risk in several populations. To date, many case-control studies have investigated the role of the GCK -30G>A polymorphism in the development of GDM and T2DM among various populations. Their results, however, remain controversial and ambiguous. Regarding the effect of GCK -30G>A on T2DM risk, the discrepancies between the results of related studies were even more significant because of the low power of some of the small-scale association studies, which mainly concluded that there was a significant association in whites (Rose et al., 2005; Cauchi et al., 2008; Murad et al., 2010; Tam et al., 2010). Several previous reviews mainly focused on the evidence regarding T2DM-associated common variants and GDM susceptibility, but they did not consider the combined effect of the variants on GDM and T2DM (Mao et al., 2012; Fu et al., 2013; Wang et al., 2013; Zhang et al., 2013). This systematic review provides a more comprehensive summary of the currently available evidence on the association between the GCK -30G>A polymorphism and the risk of GDM and T2DM. In this meta-analysis, GCK -30G>A was associated with increased GDM susceptibility in the overall population, whereas the association with T2DM was significant only in white populations.
In this meta-analysis, subgroup analyses were performed to evaluate between-study heterogeneity. In the stratified analysis by ethnicity, a significant association was found in whites for the polymorphism under most genetic models. However, few significant results were detected among Asians. There are several possible reasons for such difference. First, the distribution of the A allele varies extensively between different races. Additional studies are warranted to further validate ethnic differences in the effect of this polymorphism on GDM and T2DM risk. In addition, different populations usually have different linkage disequilibrium patterns. The GCK -30G>A polymorphism may be in close linkage with different nearby causal variants in different populations. Moreover, clinical heterogeneity in factors such as age, sex ratio, body mass index, years from onset, and disease severity may also explain the discrepancy. Finally, such different results could also be explained by the study design or sample size. Because significant between-study heterogeneity was found in the Asian subgroup, the results regarding Asian population must be interpreted with caution.
In interpreting the results of this meta-analysis, some specific issues need to be addressed. First, only published studies were included, and as a result, publication bias may have occurred. However, this was not shown by statistical analysis. Second, as with other complex traits, GDM and T2DM risk may be modulated by other genetic markers besides the GCK gene. Thus, fully elucidating the pathogenesis of GDM and T2DM would demand an investigation into the association and combined interaction of many gene variants with GDM and T2DM risk. Third, the sample size for GDM in Asians and Africans, as well as T2DM in Africans, limited the statistical power of the results. Thus, further studies of a wider spectrum of patients should be carried to investigate the role of the variant in different ethnicities. Last, meta-analyses might miss true effects in the presence of even modest between-study heterogeneity because they are based on the assumption of etiologic homogeneity across studies. As a result of heterogeneity, summary estimates provided in the present study could reflect only a crude analysis. Thus, further well-designed studies need to focus on exploring sources of heterogeneity. Despite these limitations, our study is the first comprehensive meta-analysis of all eligible studies on the association between the GCK -30G>A polymorphism and GDM and T2DM risk.
In summary, the current meta-analysis provides additional evidence supporting the hypothesis that the risk allele of the GCK -30G>A polymorphism may increase GDM and T2DM risk in whites, whereas future studies are still needed to confirm the effect of this polymorphism on risk in Asians and Africans. Thus, GCK -30G>A can probably be used with other genetic markers together to identify individuals at high risk of GDM and T2DM. However, because of the limitations of this study, these results should be interpreted with caution. Future large-scale studies are needed to confirm their accuracy.
Footnotes
Author Disclosure Statement
No competing financial interests exist.