
Abstract
Background:
Aberrant activation of the canonical WNT or WNT/β-catenin signaling pathway plays a pivotal role in multiple types of cancers. WNT5A, a nontransforming WNT protein suppressing the Wnt/β-catenin signaling pathway, is frequently detected to be hypermethylated in colorectal cancer (CRC). In this study, we investigated the prognostic value of WNT5A methylation in human patients and its potential underlying molecular mechanisms in cultured human CRC cells.
Methods:
We measured WNT5A mRNA level using qRT-PCR and DNA methylation using methylation-specific PCR (MSP) in HCT116, HT29, SW620, HCT8, LoVo, SW480, and Rko CRC cells. 5-FU-mediated tumor suppression was determined by measuring cell viability using MTT assay in cultured CRC cells. We also determined whether WNT5A methylation was associated with drug response and progression-free survival in CRC patients (n = 126) treated with 5-FU-based chemotherapy as first-line treatment.
Results:
WNT5A expression inversely correlated with methylation status of CRC cells. Moreover, WNT5A expression was restored upon demethylation in hypermethylated cells. 5-FU-induced tumor suppression (reduced cell viability) was reduced by WNT5A overexpression in hypermethylated HCT116 and SW620 cells and enhanced by WNT5A downregulation in unmethylated LoVo and SW480 cells. In 5-FU-treated CRC patients, WNT5A methylation status is associated with better drug response and longer progression-free survival, suggesting that 5-FU is more effective in CRC with WNT5A hypermethylation.
Conclusion:
WNT5A methylation status is prognostic and is useful as a potential drug selection biomarker in CRC.
Introduction
WNT
WNT signaling pathway is often found to be dysregulated in multiple types of cancers (Klaus and Birchmeier, 2008). It is particularly important in colorectal cancer (CRC), the second leading cause of cancer deaths among adults, as the majority of the CRC cases are characterized with mutations in the adenomatous polyposis coli (APC) gene, a negative regulator of WNT signaling, resulting in aberrant activation of WNT canonical signaling (Novellasdemunt et al., 2015). Interestingly, noncanonical and antagonistic WNT member WNT5A is frequently found to be aberrantly methylated in its promoter region in the CRC, suggesting that WNT5A methylation or gene silencing has a potential role in CRC tumorigenesis (Ying et al., 2008; Hibi et al., 2009).
It is generally believed that aberrant DNA methylation is an important mechanism of gene inactivation in patients with CRC (Issa, 2004; Kondo and Issa, 2004). WNT signaling pathway is one of the key signaling pathways that are altered through epigenetic action in cancer (Serman et al., 2014). Limited studies suggested that WNT5A gene methylation is associated with clinicopathological characteristics, including microsatellite instability and BRAF V600E mutation in CRC (Rawson et al., 2011). It has also been shown that WNT5A methylation is more frequently found at earlier stages (Hibi et al., 2009). Additional studies have found that WNT5A can interfere with the effect of chemotherapies involving 5-FU in pancreatic cancer (Wei et al., 2014). However, it remains ambiguous whether WNT5A methylation affects chemotherapies in CRC (Sherwood, 2015). In this study, we will determine the role of WNT5A methylation in 5-FU-based chemotherapy using human CRC cell lines and its prognostic value in CRC patients receiving first-line 5-FU treatment.
Materials and Methods
Human patients and tissue samples
A total of 126 primary sporadic adenocarcinomas were collected between 2000 and 2001 from the First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, China. All the specimens were obtained with full ethical approval from the institution committee. The mean age was 58.3 years (range, 28-67 years). None of the patients in this study had undergone preoperative chemotherapy, radiotherapy, or other therapies. Only patients who received first-line 5-FU-based adjuvant chemotherapy plus leucovorin intravenous injection weekly for 6 weeks were included. All patients were followed up for 5 years or until death.
Cell culture and transfection
The human CRC cell lines HCT116, HT29, SW620, HCT8, LoVo, SW480, and Rko cells were grown at 37°C with 5% CO2 and maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 50 mg/mL streptomycin, and 50 mg/mL penicillin. Cells were transfected with pcDNA3.1-WNT5A or pcDNA3.1 empty vector as control, using Effectene reagent (Qiagen) according to the manufacturer's instruction. The stable cell lines were established following selection with 400 mg/mL of G418 (Invitrogen) and maintained with 200 mg/mL G418. For downregulation experiments, cells were transfected with siRNA targeting WNT5A (SMARTpool 10 nM; Dharmacon) or negative control siRNA (siNTC 10 nM; Dharmacon) using DharmaFECT transfection reagent (Dharmacon) as per vendor's instruction. To determine cell viability, cells were treated with 5-FU at 4 μM.
5-Aza-2′-deoxycytidine treatment
HCT116 and SW620 cells were treated with 5 μM of 5-Aza-2′-deoxycytidine (Sigma) as described in details elsewhere (Qin et al., 2008). Cells were collected 0 to 5 days after treatment, and total RNA was extracted for qRT-PCR.
RNA isolation and quantitative assay
Total RNAs were extracted from human tissue in FFPE blocks with RNeasy FFPE Kit or cultured cells with RNeasy Kit (Qiagen) as per vendor's instruction. RNAs were checked with quality control. Fluorescent real-time RT-PCR was performed on the 7500 Real-Time PCR System (Applied Biosystems). The primers used included: WNT5A forward 5′-caggcttaacccggtcgc- 3′ and WNT5A reverse 5′-ctggcattctttgatgcctg-3′ (Ying et al.
Western blotting
The following antibodies were used in the western blot experiments: WNT5A, rabbit polyclonal (ab72583; Abcam) and β-actin, mouse monoclonal (clone AC-74, A2228; Sigma Aldrich). Protein lysates were prepared from cultured cells in ice-cold Tris buffer (20 mM; pH 7.5) containing 137 mM of NaCl, 2 mM of EDTA, 1% Triton X, 10% glycerol, 50 mM of NaF, 1 mM of DTT, and a protease inhibitor cocktail (Sigma). The protein lysates were then subjected for electrophoresis (SDS-PAGE) and transferred onto PVDF membranes (Amersham). The membranes were blocked with 5% nonfat milk and 0.1% Tween 20 in TBS, followed by incubation with primary antibody (RT 1 h) and HRP-tagged goat anti-mouse IgG secondary antibody. The blots were visualized and quantitated using Amersham ECL Western Blotting Detection Kit (Amersham).
Methylation-specific PCR
Genomic DNA was extracted from cultured cells using DNeasy Tissue Kit (Qiagen). Methylation status of the CpG islands in the WNT5A gene promoter was carried out using methylation-specific PCR (MSP) as described in detail elsewhere (Ying et al., 2008). In brief, 2 μg of genomic DNA was treated with sodium bisulfite using the EZ Methylation-Gold Kit (Zymo Research). Five percent of treated DNA was used for each MSP reaction (50 μL), including 25 μL 2× GoTaq Master Mix (Promega) and 0.25 μM of each primer. Analysis of the WNT5A (GenBank: NC_000003) has indicated a 229 bp CpG island with C + G content >60% located between nt47 and nt225 of WNT5A promoter region. The following methylated (M) and unmethylated (U) primers were used: M-primers (−448F/−326R): sense 5′-GTATTTTTCGGAGAAAAAGTTATGC-3′ and antisense 5′-AACCGCGAATTAATATAAACGTC-3′; for U-primers (−449F/− 325R): sense 5′-GGTATTTTTTGGAGAAAAAGTTATGTG-3′ and antisense 5′-CAACCACAAATTAATATAAACATC-3′ (All positions were labeled according to the transcription start site being +1). PCR was conducted on a Thermal Cycler 2720 with the following PCR conditions: 95°C for 10 min, 35 cycles of reaction (95°C for 30 s, 60°C for 30 s, and 72°C for 1 min), and 72°C for 7 min. After bisulfite conversion, the “C” of a methylated “CpG” would remain “C”, and the “C” of an unmethylated “CpG” would be converted to “U” and changed to “T” after PCR reaction. Results were summarized from 4 independent experiments.
MTT assay
In cultured cells, 10 μL of 5 mg/mL MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide] in PBS was added (final concentration 0.5 mg/mL) and incubated for 4 h. The MTT solution and medium were removed, and 100 μL DMSO was added to each well. Absorbance was measured at 570 nm using the ELISA microplate reader.
Statistical analyses
Statistical analyses were conducted using GraphPad Prism 6 (GraphPad Software, Inc.). Data were presented as mean ± SEM. Statistical significance was determined by Student's t-test for pairwise comparisons or by one-way ANOVA with Tukey's multiple comparison tests or two-way ANOVA with Bonferroni's multiple comparison tests for multiple comparisons. Survival curve was created using log-rank (Mantel-Cox) test. p < 0.05 was considered statistically significant.
Results
WNT5A expression is silenced by methylation in selected CRC cell lines
The expression level of WNT5A was examined by RT-PCR in a panel of CRC cell lines. Loss of WNT5A expression was observed in HCT116, HT29, SW620, and HCT8 cells, but not in LoVo, SW480, and Rko cells (p < 0.01 by one-way ANOVA with Tukey's multiple comparison tests, Figure 1A). MSP showed that WNT5A was completely or partially methylated in the silenced cell lines, but unmethylated in WNT5A-expressed cells (Fig. 1B). Moreover, WNT5A expression was restored in a time-dependent manner in HCT116 and SW620 cells treated with 5-Aza-2′-deoxycytidine (DAC, 5μM) (two-way ANOVA with Bonferroni's multiple comparison tests, Fig. 1C, D), a DNA methylation transferase inhibitor that induces re-expression of methylated genes through demethylation (Qin et al., 2009). Taken together, these results suggested that WNT5A expression is epigenetically regulated by promoter region methylation in selected CRC cells.
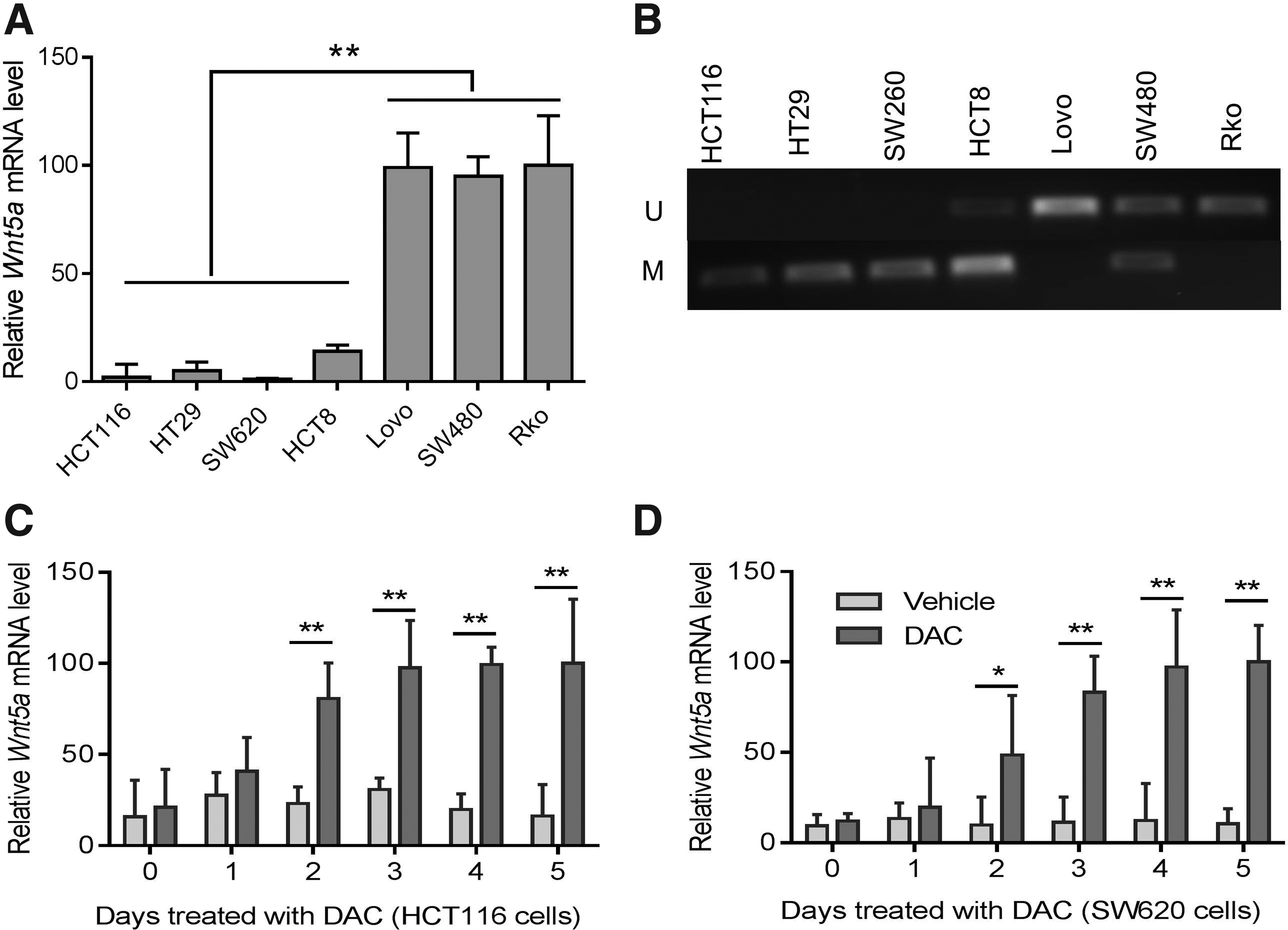
WNT5A expression is silenced by methylation in selected CRC cell lines.
WNT5A expression inversely mediates 5-FU-induced tumor suppression in CRC cells
We then investigated whether WNT5A expression was related with the sensitivity of CRC cells to 5-FU. To do so, we first overexpressed WNT5A in methylated HCT116 and SW620 cells to determine whether WNT5A restoration reduced 5-FU-mediated tumor suppression by MTT assays. The cells that stably express WNT5A or empty control vector were selected by G418, and the expression of WNT5A in these cells was confirmed by western blotting (Fig. 2A, B). MTT assays indicated that the cell viability was significantly higher (indicating reduced tumor suppression) in 5-FU-treated HCT116 and SW620 cells stably expressing WNT5A than in those expressing control vector (p < 0.01 and 0.05, respectively, by two-way ANOVA with Bonferroni's multiple comparison tests, Fig. 2C). We next knocked down WNT5A using siRNA in unmethylated LoVo and SW480 cells, and the efficiency of WNT5A siRNA was demonstrated by western blot experiments showing that WNT5A protein level was significantly lower in both LoVo and SW480 cells treated with WNT5A siRNA (32.1% and 42.4% of control, respectively) than in those with negative control siRNA (p < 0.05 by two-way ANOVA, Fig. 2D, E). MTT assays revealed that WNT5A knockdown significantly improved 5-FU-mediated tumor suppression as demonstrated by decreased cell viability (p < 0.05, two-way ANOVA, Fig. 2F). These results suggested that WNT5A methylated cells that expressed lower or no WNT5A were more sensitive to 5-FU-treatment in CRC.
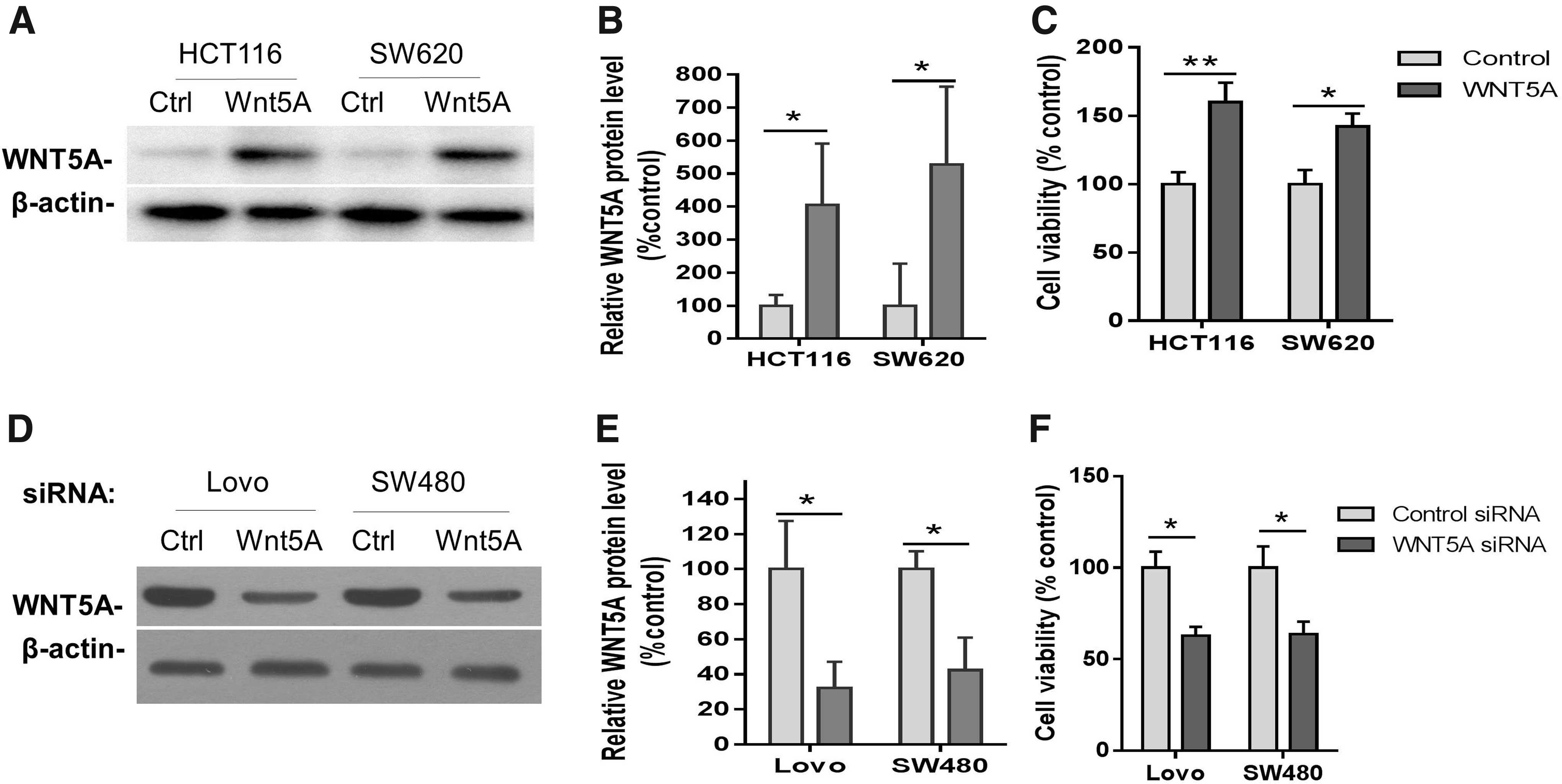
WNT5A regulates tumor cell viability in hypermethylated or unmethylated CRC cells treated with 5-FU.
WNT5A methylation is associated with positive response to first-line 5-FU treatment in CRC patients
To investigate the potential prognostic value of methylation of WNT5A in CRC, the survival rate was analyzed from the patients who received first-line 5-FU-based adjuvant chemotherapy. There was no significant association between WNT5A methylation and tumor stage, tumor grade, gender, or age (Table 1). The methylation level of WNT5A was significantly higher in the responding group (n = 39) than in the nonresponding group (n = 87) (p < 0.001, Student's t-test) (Fig. 3A and Table 1). Meanwhile, the patients with WNT5A methylation were associated with longer progression-free survival (p < 0.005, Log-rank Mantel-Cox test) (Fig. 3B). These results suggested that response to 5-FU treatment is associated with WNT5A methylation. Therefore, WNT5A methylation can be explored as a valuable marker in CRC prognosis.
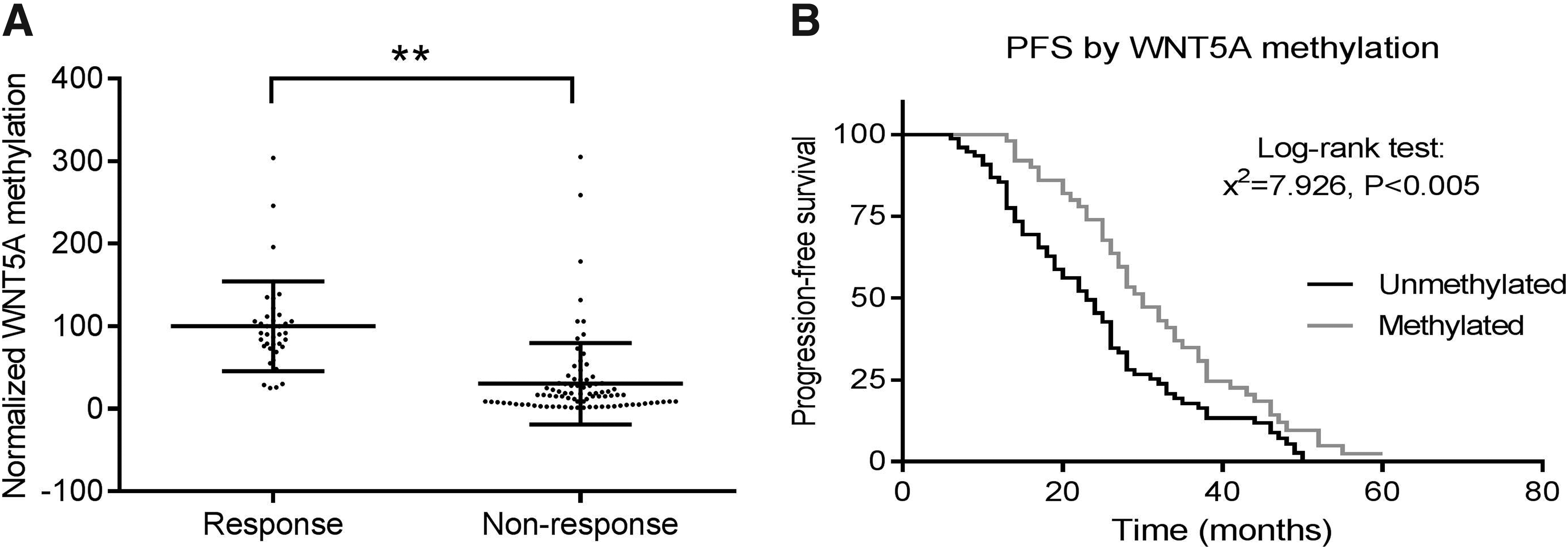
WNT5A promoter methylation was associated with 5-FU response and longer PFS in CRC patients.
χ2 test.
M, methylated; U, unmethylated.
Discussion
The WNT signaling pathway is important in the regulation of embryogenesis and carcinogenesis under physiological and pathological conditions, respectively (Clevers, 2006; Klaus and Birchmeier, 2008). In CRC, tumorigenesis is often associated with WNT signaling activation, resulted from mutations in either the β-catenin gene or the APC gene, which encodes a β-catenin inhibitor (Clevers, 2006; Klaus and Birchmeier, 2008). In addition, WNT5A, another canonical WNT inhibitor as a noncanonical WNT member, was frequently found to be methylated in the promoter region (Wang et al., 2007; Ying et al., 2008; Hibi et al., 2009; Li et al., 2010; Rawson et al., 2011). Consequently, WNT5A gene expression was silenced or downregulated, losing antagonistic action against or aggravating the aberrant activation of canonical WNT signaling. Consistent with this idea, our results in the current study have shown that WNT5A mRNA level was significantly downregulated in methylated CRC cell lines. The expression of WNT5A protein can be restored in these cell lines when they were treated with methylation inhibitor. WNT5A methylation was also found in other type of cancers, including prostate cancer (Wang et al., 2007) and esophageal squamous cell carcinomas (Li et al., 2010).
We next asked whether WNT5A upregulation has any clinical implication. Our in vitro results indicated that WNT5A overexpression desensitizes 5-FU treatment-induced tumor suppression in WNT5A methylated HCT116 and SW620 CRC cells. In contrast, downregulation of WNT5A expression promoted 5-FU treatment-induced tumor suppression in unmethylated LoVo and SW480 CRC cells. Thus, WNT5A methylation status is likely a valuable biomarker for treatment selection in CRC. These results are consistent with the notion that WNT5A functioned as an antagonist of the canonical Wnt signaling pathways, which were aberrantly activated to mediate tumor cell proliferation (Sherwood, 2015). It is possible that WNT5A methylation is an important intermediate step in CRC formation and progression, by contributing at least, in part, to the overactivation of the WNT/β-catenin signaling and subsequently cancer abnormal cell proliferation. Thus, WNT5A hypermethylation can be a surrogate for overactivation of the canonical Wnt signaling, which promotes tumor cell proliferation. Meanwhile, 5-FU is a drug known as a pyrimidine analog and commonly used in CRC to treat cancer through irreversible inhibition of thymidylate synthase, which in turn inhibits cell proliferation (Patel, 2011; Wilson et al., 2014). Therefore, through interfering with DNA replication to prevent abnormal tumor cell proliferation, 5-FU is likely more effective in CRC cells with WNT5A hypermethylation.
Indeed, our study has shown that WNT5A methylation is associated with a better drug response in CRC patients that received 5-FU as first-line treatment. This result is also consistent with a previous study showing that WNT5A overexpression is associated with poor clinical outcome in patients with ovarian cancer or ovarian cancer cell lines receiving chemotherapeutic drugs, including 5-FU (Peng et al., 2011). Thus, it is likely that WNT5A methylation is involved in mediating chemoresistance in cancers, including CRC and ovarian cancer. In addition to WNT5A, 5-FU also showed a better drug response in patients with methylation of other genes, such as HYLA2 (Pfutze et al., 2015), suggesting that epigenetic factors using different molecular mechanisms have a much broader range of impact on 5-FU-based chemotherapies, as well as on other types of treatments in CRC.
It remains elusive how WNT5A and its methylation status play a role in tumorigenesis and/or tumor suppression in CRC, as well as in other type of cancers. A previous study has shown that WNT5A overexpression suppressed the expression of CCND1, likely through promoting β-catenin degradation as CCND1 is a β-catenin target gene (Ying et al., 2008). A recent study using Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes analysis for the CRC genes has identified CCND1 as one of the hub genes in a protein-protein interaction network, further supporting the importance of Cyclin D1 in CRC (Liu et al., 2015). Interestingly, WNT5A was found to promote CCND1 expression, which is associated with gemcitabine chemoresistance, in pancreatic cancer (Wei et al., 2014). The opposing role of WNT5A among different types of cancer indicated that this effect is context dependent. Moreover, WNT5A is hypomethylated (with upregulated gene expression) and mediates multidrug resistance through ABCB1 in breast cancer (Hung et al., 2014), further supporting the molecular heterogeneity in the role of WNT5A among different types of cancer.
In conclusion, we have demonstrated that WNT5A methylation is associated with better therapeutic response in CRC patients who received 5-FU as first-line treatment. Consistent with this result, our in vitro experiments have shown that WNT5A gene expression can be restored through demethylation in hypermethylated CRC cells, and WNT5A expression level is inversely associated with 5-FU-mediated tumor suppression. Taken together, our study has suggested that WNT5A methylation can be further investigated as a biomarker for its prognostic value and for treatment selection in CRC.
Footnotes
Acknowledgment
This work was supported by grant (NSFC No. 81272371) from the Fund by National Nature Science Foundation of China.
Author Disclosure Statement
No competing financial interests exist.