
Abstract
Aim: To investigate the association between autism spectrum disorder (ASD) and the phospholipase A2 group IVC (PLA2G4C) and phospholipase A2 group XIIA (PLA2G12A) polymorphisms in the Northeast Han Chinese population. Materials and Methods: A total of 68 family trios (children diagnosed with ASD and their unaffected parents) were enrolled. Five single-nucleotide polymorphisms (SNPs) (rs9226, rs1045376, rs251684, rs2307279, and rs156631) in PLA2G4C and four SNPs (rs6533451, rs2285714, rs2285713, and rs11728699) in PLA2G12A were selected and genotyped. The association between the SNPs and ASD was analyzed using the transmission disequilibrium test. Results: Our results showed a significant association between ASD and the rs251684 variant of PLA2G4C (transmitted/nontransmitted = 36/21, χ2 = 3.947, p = 0.047), but no association between ASD and the other eight SNPs investigated (all p > 0.05). Moreover, we found no preference in the transmission of haplotypes constructed for either PLA2G4C or PLA2G12A. Conclusion: The rs251684 polymorphism of PLA2G4C may be associated with ASD risk.
Introduction
A
The phospholipase A2 (PLA2) superfamily consists of a diverse group of enzymes that hydrolyze glycerophospholipids to produce free fatty acids and lysophospholipids, both of which serve as precursors in the production of signaling molecules (Six and Dennis, 2000). Arachidonic acid (AA), one of the fatty acids released from the hydrolyzation of membrane phospholipids via PLA2, functions as a potent mediator of inflammation signaling. The lysophospholipids, another product of PLA2, are key regulators of phospholipid remodeling and membrane perturbation. In pathological conditions, increased PLA2 activity may potentially lead to nervous system dysfunction and cognitive impairment (Farooqui et al., 2006; Ong et al., 2015). PLA2s can be classified into five main isoforms: cytosolic PLA2 (cPLA2), secretory PLA2 (sPLA2), calcium-independent PLA2 (iPLA2), plasmalogen-selective PLA2 (PlsEtn-PLA2), and lipoprotein-PLA2 (Lp-PLA2). Moreover, sPLA2, cPLA2, and iPLA2 exist in different regions of the brain, including the neurons, microglia, and astrocytes (Molloy et al., 1998; Zanassi et al., 1998; Balboa et al., 2002).
cPLA2γ, one of cPLA2 isoforms, is expressed in the brain, skeletal muscle, and heart (Underwood et al., 1998; Tucker et al., 2005). The cPLA2γ gene (PLA2G4C) is located on chromosome 19q13.3 and was first discovered using a BLAST analysis in 1998 (Pickard et al., 1999). Furthermore, the cPLA2γ activity is calcium independent because it lacks a C2 domain (Murakami et al., 2003). cPLA2γ is the paralogue of cPLA2α, which is the only PLA2 with a substantial preference for AA generation over the production of other fatty acids (Clark et al., 1991; Asai et al., 2003). AA and docosahexaenoic acid (DHA) are essential for learning, memory, neurogenesis, neuronal differentiation, neural plasticity, and signal transduction (Tamiji and Crawford, 2010). In addition, the plasma DHA/AA ratio correlates with ASD (Yui et al., 2016a). Most importantly, the cyclooxygenase (COX) pathway converts AA to prostaglandins E2 (PGE2) (Murakami et al., 2003), which alters the Wnt-dependent migration and proliferation of neuroectodermal stem cells implicated in ASD (El-Ansary and Al-Ayadhi, 2012; Wong et al., 2014).
Unlike cPLA2γ, sPLA2 has a highly conserved Ca2+-binding loop and a His-Asp dyad at the catalytic site (Lambeau and Gelb, 2008; Titsworth et al., 2008). Furthermore, sPLA2 plays an important role in central nervous system injuries and neurotrauma (Titsworth et al., 2008). sPLA2 is associated with synaptosomes and synaptic vesicle fractions (Farooqui et al., 2006). Notably, abnormalities at the synapse are found in ASD (Garber, 2007). The phospholipase A2 group XIIA (PLA2G12A) gene is located on chromosome 4q25 and encodes one of the variants of sPLA2 (sPLA2G12A) (Lambeau and Gelb, 2008). PLA2G12A is expressed in different regions of the brain, but is mainly expressed in the prefrontal cortex, which correlates with the cognitive phenotype of ASD (Ee et al., 2014; Hashemi et al., 2016).
The heritability of ASD was estimated to be 50% (Sandin et al., 2014). Accumulating twin- and family-based studies further suggest the critical role of genetic factors in ASD. For instance, babies born into families with an ASD sibling have a 10 times greater risk of ASD than babies born into other families (Hallmayer et al., 2011; Sandin et al., 2014). Due to the potential involvement of phospholipase A2 group IVC (PLA2G4C) and PLA2G12A in ASD, we designed a family-based study and used the transmission disequilibrium test (TDT) to investigate whether single-nucleotide polymorphisms (SNPs) in PLA2G4C and PLA2G12A were associated with ASD in the Northeast Han Chinese population.
Materials and Methods
Ethics statement
The study was approved by the Ethics Committee of the First Hospital of Jilin University, and written informed consents were obtained from the unaffected parents of an autistic child.
Study subjects
A total of 68 families were recruited from the First Hospital of Jilin University in Changchun, Jilin Province, China. Each family contained an autistic child, but the parents were unaffected. The 68 unrelated ASD children (54 boys, 14 girls) were diagnosed by Pediatric Neurology and Neurorehabilitation doctors based on the Diagnostic and Statistical Manual of Mental Disorders (fifth edition) (American Psychiatric Association, 2013). The mean age of the patients was 3.91 years (range 2-10). Clinical examinations, including standard karyotying, fragile-X testing, computed tomography scan, and other clinical analyses, were performed for all probands to exclude fragile X syndrome, Rett syndrome, and other known syndromes associated with ASD. All of the participants belonged to the Han population in Northeast China.
Tag SNP selection
We selected Tag SNPs for PLA2G4C and PLA2G12A using the HapMap database and Tagger implementation in the Haploview 4.2 software (Barrett et al., 2005), according to the following selections: Han Chinese in the Beijing population, r2 ≥ 0.8, tagger pairwise as the pairwise method, and minor allele frequency >10%. We selected SNPs in PLA2G4C and PLA2G12A, including rs9226, rs1045376, rs251684, rs2307279, rs156631, rs6533451, rs2285714, rs2285713, and rs11728699.
DNA extraction and SNP genotyping
Genomic DNA was extracted from peripheral blood samples using a DNA extraction kit, according to the manufacturer's instructions (ClotBlood DNA kit, CWBio, Beijing, China). Genotyping of each variant was performed using polymerase chain reaction and matrix-assisted laser desorption/ionization time of flight mass spectrometry in the MassARRAY system (Sequenom, San, Diego, CA) (Buetow et al., 2001).
Statistical analyses
The Hardy-Weinberg equilibrium of the genotype distributions was determined by the goodness of fit chi-squared (χ2) test. The TDT is a family-based method to evaluate the transmission of the associated marker allele from a heterozygous parent to an affected offspring (Spielman et al., 1993; Spielman and Ewens, 1996). Linkage disequilibrium (LD) blocks were constructed to evaluate the association between ASD and different haplotypes using the Haploview 4.2 software (Barrett et al., 2005). The statistical tests were two tailed, and statistical significance was defined as p < 0.05.
Results
We genotyped the nine tag SNPs in all 68 families. However, due to Mendelian errors in 3 of the families, we ultimately only analyzed 65 families. The remaining children included 51 boys and 14 girls, with a mean age of 4 years (range 2-10). To investigate the relationship between PLA2G4C and cPLA2γ as well as the relationship between PLA2G12A and sPLA2G12A, we mainly focused on the exons and 3′ untranslated regions of both genes. The genotype distributions of the nine SNPs were in Hardy-Weinberg equilibrium (p > 0.05) (Table 1).
HWE, Hardy-Weinberg equilibrium; MAF, minor allele frequency; PLA2G4C, phospholipase A2 group IVC; PLA2G12A, phospholipase A2 group XIIA; SNP, single-nucleotide polymorphism; UTR, untranslated regions.
We next investigated the genotype distribution and allele frequency of our study participants. The genotypic frequency of homozygous mutations in the SNPs (rs251684, rs2307279, and rs156631) of PLA2G4C and the SNPs (rs6533451, rs2285714, and rs11728699) of PLA2G12A ranged from 4.6-12.3% to 1.5-15.4% in the patients, and 2.3-11.5% to 6.9-14.6% in the parents, respectively. However, in PLA2G4C (rs9226) and PLA2G12A (rs2285713), no homozygous mutations were found in either the patients or the parents. For rs1045376 in PLA2G4C, the genotypic frequency of homozygous mutations is 0% in patients and 2.3% in parents. In addition, the mutant allele frequency in PLA2G4C and PLA2G12A ranged from 10.0-31.5% to 3.1-37.7% in the patients, and 7.7-29.2% to 2.3-39.6% in the parents, respectively (Table 2).
Data are expressed as n (%).
1 for ancestral allele; 2 for mutation allele; —, no data.
Furthermore, we used TDT to investigate the association between ASD and the five SNPs of PLA2G4C or the four SNPs of PLA2G12A. We found a significant association between ASD and the rs251684 variant of PLA2G4C (transmitted/nontransmitted = 36/21, χ2 = 3.947, p = 0.047). Our results revealed that the A allele of rs251684 exerted excessive transmission from parents to their ASD children. However, we found no association between ASD and the other SNPs of PLA2G4C and PLA2G12A (all p > 0.05) (Table 3).
NT, number of nontransmissions; T, number of transmissions.
After we obtained the ASD association results of each SNP in both genes, we further analyzed the relationship between ASD and the haplotypes. We constructed one block from the five SNPs (rs9226, rs1045376, rs251684, rs2307279, and rs156631) in PLA2G4C, and another block from the four SNPs (rs6533451, rs2285714, rs2285713, and rs11728699) in PLA2G12A. Our results showed no preference in the transmission of any haplotypes (Table 4). Moreover, we found an LD between rs251684 and rs2307279 in PLA2G4C, and a strong LD among all four SNPs in PLA2G12A (Figs. 1 and 2).
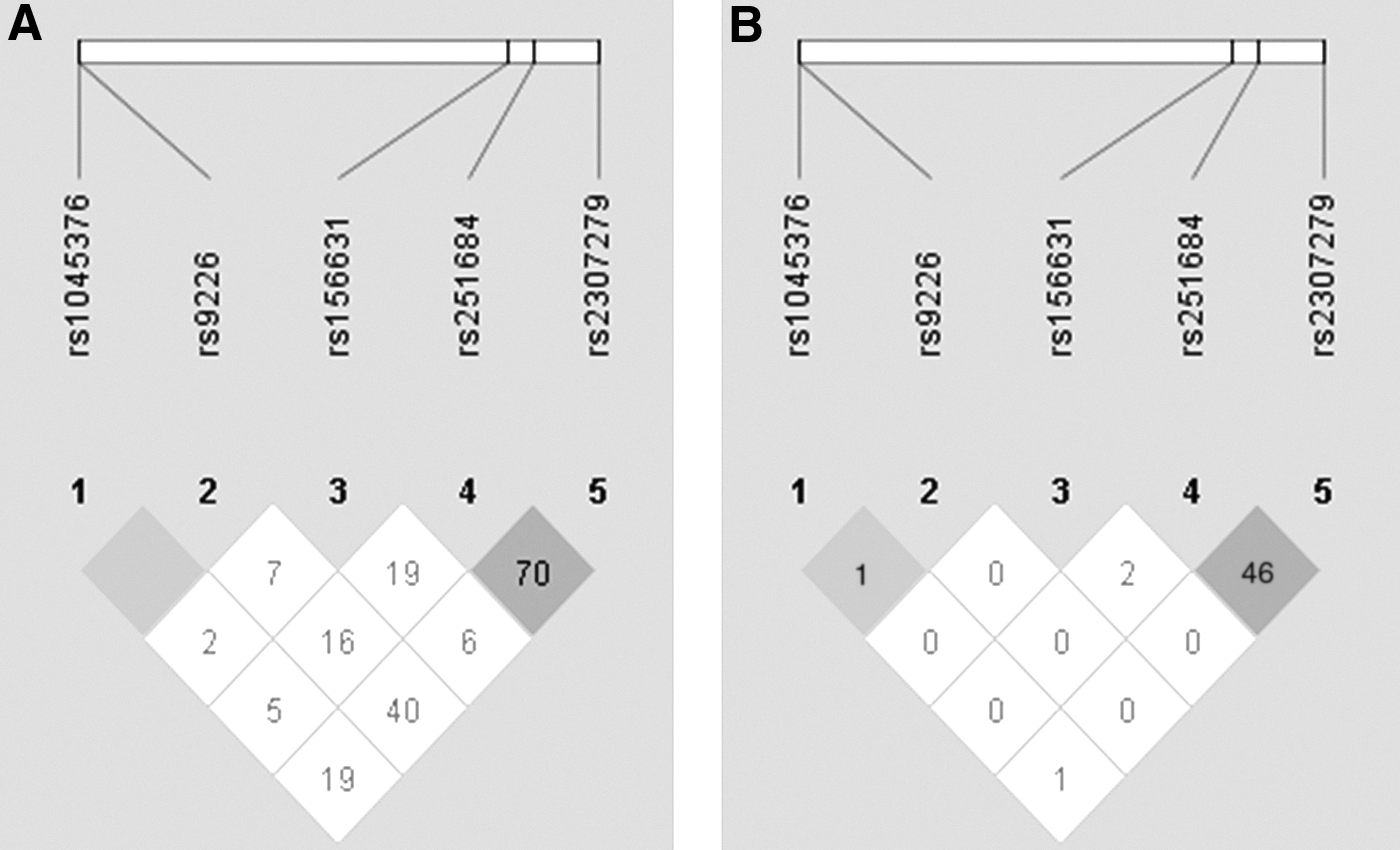
LD structure and relative chromosomal positions of the five SNPs in PLA2G4C.
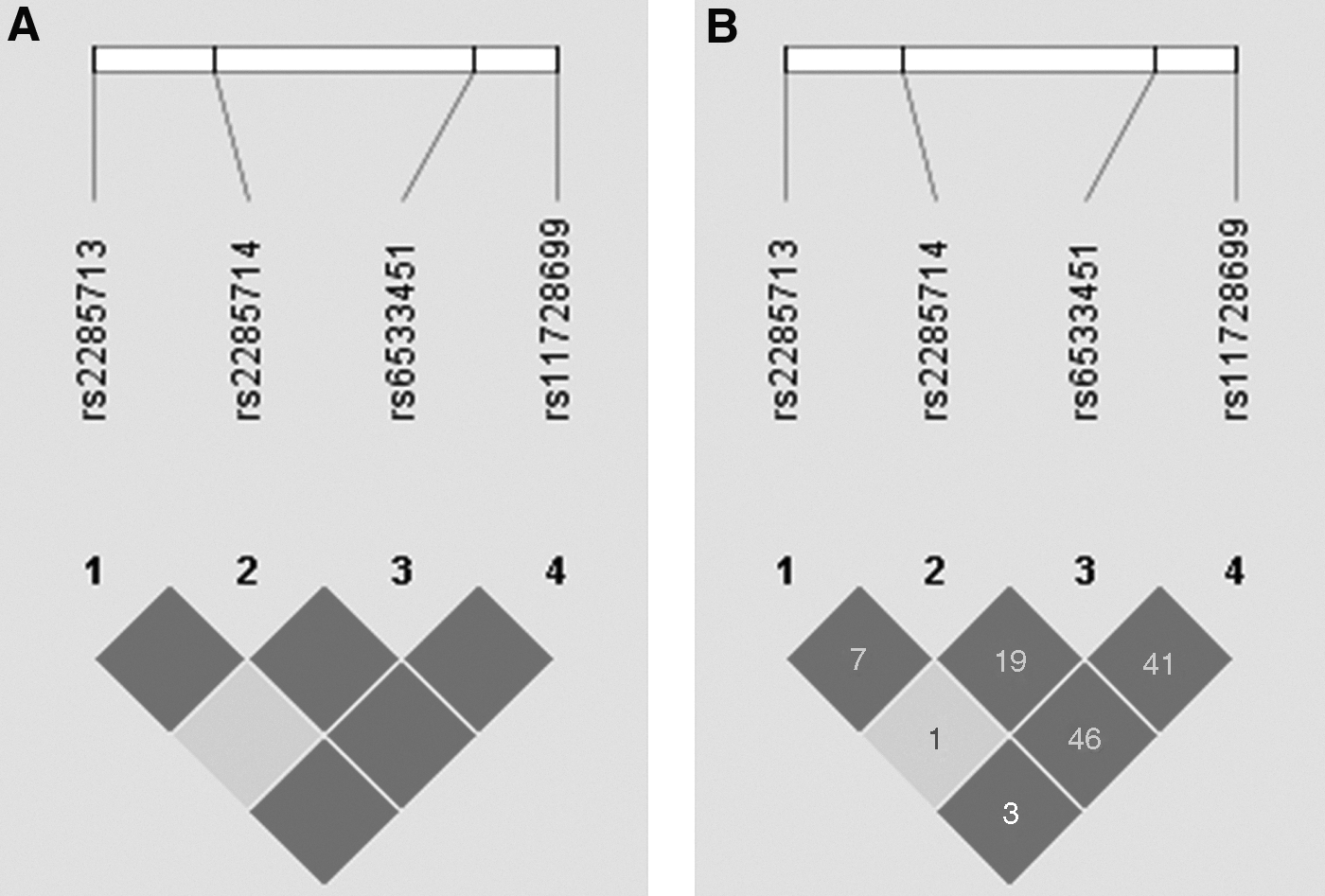
LD structure and relative chromosomal positions of the four SNPs in PLA2G12A.
Discussion
In our study, we investigated 5 SNPs (rs9226, rs1045376, rs251684, rs2307279, and rs156631) in PLA2G4C and 4 SNPs (rs6533451, rs2285714, rs2285713, and rs11728699) in PLA2G12A from 65 ASD trios using TDT. Our results revealed a significant association between ASD and the rs251684 variant of PLA2G4C. However, we did not find a significant association between ASD and the other eight SNPs in the Northeast Han Chinese population. Moreover, our results showed no preference in the transmission of any haplotypes.
PLA2 is critical for normal brain function and maintaining basal levels of lysophospholipid, AA, and platelet activating factor (Farooqui et al., 2006). Dysregulated phospholipid metabolism is involved in ASD, schizophrenia, and other neurodevelopmental disorders (Brown and Austin, 2011). Moreover, the PLA2 polymorphism is substantially implicated in PLA2 function (Masbuchin et al., 2015; Ong et al., 2015). Thus, the relationship between PLA2 and ASD may provide a helpful clue to the etiology of ASD.
cPLA2γ plays an essential role in phospholipid remodeling and AA metabolism (Murakami et al., 2003). The plasma DHA/AA ratio correlates with plasma ceruloplasmin levels and represents a surrogate biomarker for the behavioral symptoms of ASD patients (Yui et al., 2016a, 2016b). AA is converted into PGE2 via the COX-1 pathway during the immediate prostaglandin biosynthetic response, but is converted via the COX-2 pathway in the delayed response (Murakami et al., 2003). PGE2 is an endogenous modulator of cerebellar development during the second postnatal week (Dean et al., 2012). Because the cerebellum is indispensable for cognitive and sensory functions of the brain, cerebellar abnormalities strongly correlate with autistic traits (Allen and Courchesne, 2003). In addition, PGE2 modulates membrane excitability and synaptic transmission in the hippocampal CA1 pyramidal neurons (Chen and Bazan, 2005). Moreover, dysfunctional synaptic transmission partially results in ASD (Tabuchi et al., 2007; van Spronsen and Hoogenraad, 2010). PGE2 also interacts with canonical Wnt signaling via protein kinase A and phosphatidylinositide 3-kinases, and affects Wnt-dependent cell behaviors and gene expression in neuroectodermal stem cells (Wong et al., 2014). Wnt signaling regulates diverse neuronal functions (Ciani and Salinas, 2005). Thus, an altered Wnt pathway may affect a subset of individuals with ASD (Kalkman, 2012). PLA2G4C polymorphisms are also associated with schizophrenia, another neurodevelopmental disorder (Tao et al., 2005; Yu et al., 2005). When we investigated the relationship between ASD and the PLA2G4C polymorphisms, we found that the rs251684 variant of PLA2G4C is associated with ASD. We also investigated the association between PLA2G12A polymorphisms and ASD, but we did not find an association.
To the best of our knowledge, this is the first study on the association between PLA2 and ASD in the Northeast Han Chinese population. We performed a family-based study instead of the traditional case-control study because family-based studies are less prone to false-positive results. Furthermore, the case-control approach requires unrelated controls with a population admixture and stratification (Laird and Lange, 2006).
Our study had some limitations. First, our study may be biased by the small sample size, which may have limited our statistical power to detect any significant findings in our study population. Second, ASD is a complex disease with many factors contributing to its development, such as multiple genetic and environmental factors. Third, our discoveries were obtained from only the Northeast Han Chinese population, which means that generalizations cannot be made to other populations based on our findings. Fourth, the functional mechanism by which PLA2G4C and PLA2G12A SNPs lead to ASD is not well known. Thus, our study loci may be in LD with other casual SNPs responsible for ASD.
In conclusion, our results indicate that the rs251684 polymorphism of PLA2G4C is associated with ASD risk in the Northeast Han Chinese population.
Footnotes
Acknowledgments
This work was supported by the International Cooperation of Science and Technology Department of China (Grant No. 2015DFA31580), Scientific Research Foundation of the Health Bureau of Jilin Province of China, Jilin Provincial Key Laboratory of Neuronal Plasticity (Grant Nos. 20140622001JC and 20160622020JC), and China Postdoctoral Science Foundation (Grant No.: 2013M530989).
Author Disclosure Statement
No competing financial interests exist.