
Abstract
Aims:
Iran is located in the Asian esophageal cancer belt. It is a high-risk region for esophageal squamous cell carcinoma (ESCC). The extent to which genetic components, especially variants within miRNAs or their binding sites, contribute to risk of ESCC in the region is not yet fully understood. Herein, tests were done on an Iranian cohort to evaluate the association of miRNA-related polymorphisms in miR-423 (rs6505162) and peroxisomal biogenesis factor 6 (PEX6) (rs1129186 within a miR-149-5p-binding site) with the risk of ESCC risk.
Methods:
This study recruited 200 ESCC patients and 300 healthy individuals. Genotyping was performed using the polymerase chain reaction-restriction fragment length polymorphism method. Target genes and biological processes that are regulated by miR-423 and may be affected by a change in miR-423 expression were identified by in silico analysis.
Results:
Logistic regression analyses revealed an association between rs6505162 and ESCC, assuming codominant (AA vs. CC, odds ratios, OR [95% confidence interval, CI]: 0.32 [0.15-0.69], p-value: 0.0076), recessive (AA vs. CC+CA, OR [95% CI]: 0.35 [0.16-0.73], p-value: 0.0027), and log-additive models (OR [95% CI]: 0.69 [0.52-0.91], p-value: 0.0084). No significant association was observed for PEX6 rs1129186. In silico analyses revealed several genes and biological processes that are regulated by miR-423 in ESCC.
Conclusion:
This study identified the first evidence of an association of a miRNA-related variant with risk of ESCC in an Iranian cohort. PEX6 rs1129186 may not modulate the risk of ESCC in the cohort.
Introduction
E
There is increasing evidence to suggest that genetic susceptibility may play an important role in the etiology of EC, especially ESCC (Wu et al., 2011). Genome-wide and candidate gene association studies have yielded enormous progress in illuminating the genetic contributors of ESCC and several molecular mechanisms contributing to the pathophysiology of ESCC have been revealed (Lin et al., 2013). Specifically, the regulatory networks of miRNAs have been shown to be functionally relevant to a broad spectrum of cellular processes from differentiation to metabolism (Ardekani and Naeini, 2010; Alizadeh et al., 2015), and to human diseases such as ESCC (Wang et al., 2012; Gu et al., 2013; Zhou et al., 2013; Saadatian et al., 2014; Nariman-Saleh-Fam et al., 2016). miRNAs regulate the cellular processes altered during tumorigenesis; and the dysregulation of many miRNAs has been described in ESCC (Olive et al., 2010; Harada et al., 2016). Recently, single-nucleotide polymorphisms (SNPs) within miRNAs or their binding sites have emerged as a new category of variants that indicate susceptibility to disease (Landi et al., 2012). There is increasing evidence suggesting that SNP-induced alteration in miRNA-mediated regulation of gene expression may be important in individuals' susceptibility to many types of diseases, including ESCC (Landi et al., 2012; Zhou et al., 2013; Ghaedi et al., 2015, 2016; Bastami et al., 2016a, 2016b; Nariman-Saleh-Fam et al., 2016; Shen et al., 2016). However, the extent to which these types of variations are relevant to ESCC in the Iranian population is not yet clearly understood. This study evaluated the association of miRNA-related polymorphisms in miR-423 (rs6505162) and peroxisomal biogenesis factor 6 (PEX6) (rs1129186 within a miR-149-5p-binding site in the mRNA of this gene) with susceptibility to ESCC in an Iranian cohort. rs6505162 C>A is located in miR-423 precursor, a miRNA with altered the expression in ESCC tumors (Wang et al., 2013b). It has been reported that this SNP regulates mature miRNA expression and may contribute to the risk of ESCC (Wang et al., 2013b; Su et al., 2015; Zhao et al., 2015). PEX6 encodes a member of the AAA family of ATPases (provided by RefSeq) that has been reported as downregulated in ESCC tumors (Wang et al., 2013a) (NCBI GEO ID: GSE26886). Interestingly, there is an experimentally identified interaction between this gene and another ESCC-related element, miR-149-5p (Helwak et al., 2013). Evidence suggests that this miRNA, which is upregulated in ESCC tissue (Guo et al., 2008), regulates PEX6 transcript and this miRNA-mediated regulation of PEX6 may be hampered by a functional polymorphism in PEX6 (rs1129186) (Nariman-Saleh-Fam et al., 2016).
Materials and Methods
Study cohort
A total of 500 unrelated Iranian subjects were enrolled in this study, including 200 ESCC patients and 300 participants in a control group that were matched in terms of age (patients: 61.40 ± 12.40, control group: 62.77 ± 8.28, p-value: 0.17) and sex (sex ratio in patients: 1.02, in controls: 1.13, p-value: 1). ESCC was diagnosed by evaluations of upper gastrointestinal endoscopy and histopathology. Patients were recruited from the cancer institute of the Imam Khomeini Hospital (Tehran, Iran) with cooperation of the Medical Genetics Department of Tehran University of Medical Sciences, Reza Radiation Oncology Center (Mashhad, Iran) with cooperation of the Avicenna Research Institute of Mashhad University of Medical Sciences, and Imam Reza hospital (Tabriz, Iran) with cooperation of the Medical Genetics Department, and the Liver and Gastrointestinal Disease Research Center of Tabriz University of Medical Sciences. The mean age of patients at diagnosis was 61.4 years (range within 20-87 years); 101 patients (50.5%) were male and 99 (49.5%) were female. Participants in the control group had no personal or family history of cancer and were recruited from the individuals who had been referred for routine check-ups. They were recruited from Modarres Hospital (Tehran, Iran) with the cooperation of Medical Genetics Department of Shahid Beheshti University of Medical Sciences and Department of Genetics and Biotechnology of Islamic Azad University. The mean age for the control group was 62.7 years (range within 50-86 years); 151 control subjects (50.33%) were male and 149 (49.66%) were female. Written informed consent was obtained from all participants. The study was approved by the ethics committee of Tehran University of Medical Sciences.
DNA extraction and genotyping
Genomic DNA was extracted from peripheral blood using standard protocol (Gustafson et al., 1987). Genotyping was performed using the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) procedure. The genomic regions containing the studied SNPs were amplified with specific primers outlined in Table 1. Mismatches were inserted into the primers to create corresponding restriction endonuclease sites. The 25 μL reaction system consisted of genome DNA (∼50 ng), 12.5 μL of 2 × Taq DNA Polymerase Master Mix Red (Ampliqon), 1.5 μL forward primer (5 μM), 1.5 μL reverse primer (5 μM), and 8.5 μL sterilized water. For miR-423 rs6505162, a 143 bp genomic region was amplified using touchdown PCR. An initial denaturation of 5 min at 95°C was followed by 7 cycles at 94°C for 30 s, 66°C for 30 s, and 72°C for 30 s with annealing temperature decreasing 1°C per cycle, and then 28 cycles at 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s and a final extension step at 72°C for 5 min. The PCR product was digested with the restriction enzyme (RsaI; Thermo Scientific) according to the manufacturer's instructions. For PEX6 rs1129186, a 145 bp genomic region was amplified using touchdown PCR. An initial denaturation of 5 min at 95°C was followed by 7 cycles at 94°C for 30 s, 66°C for 30 s, and 72°C for 30 s with annealing temperature decreasing 1°C per cycle, and then 28 cycles at 94°C for 30 s, 60°C for 30 s, and 72°C for 30 s and a final extension step at 72°C for 5 min. The PCR product was digested with the restriction enzyme (XhoI; Thermo Scientific) according to the manufacturer's instructions. Six samples of the assigned genotypes were confirmed by Sanger sequencing (one sample per SNP genotype) and these samples were then served as controls for the digestion process. The no template control (NTC) PCR and digestion reactions were also performed for each SNP to monitor for possible contamination. Each NTC PCR contained all reagents, except template DNA.
Nucleotides that were mismatched to create the corresponding restriction sites are underlined.
SNP, single-nucleotide polymorphism.
Statistical analyses
Statistical analyses were performed using the R programming language (version 3.1.0). Differences between patients and the control group in terms of mean age and sex ratio (ratio of males to females) were assessed using Student's t-test and χ2 test, respectively. SNPs were assessed for significant departure from the Hardy-Weinberg equilibrium among patients, the control group, and all subjects using χ2 test. The association of miR-423 rs6505162 and PEX6 rs1129186 with ESCC was analyzed using logistic regression analyses implemented in the SNPassoc package (version 1.9-2) (Gonzalez et al., 2007). Odds ratios (OR) and 95% confidence intervals (95% CI) were calculated assuming codominant, dominant, recessive, overdominant, and log-additive genetic models. The best-fitting genetic model was selected based on the lowest Akaike's information criterion (AIC) value, as calculated by the SNPassoc (Gonzalez et al., 2007).
In silico analyses
Using in silico analyses, genes and biological processes that were regulated by miR-423 were identified. The experimentally identified targets of miR-423-5p (MIMAT0004748) and miR-423-3p (MIMAT0001340) were retrieved from miRTarBase (release 6.0, available at http://mirtarbase.mbc.nctu.edu.tw) (Chou et al., 2016). The target genes list was subjected to gene ontology annotation analysis using the Functional Annotation Clustering (FAC) tool, which was implemented in the DAVID database (Release 6.8, available at https://david.ncifcrf.gov/home.jsp) (Huang da et al., 2009a, 2009b) to obtain an overview of the biological processes that are regulated by miR-423. In FAC analysis, the classification stringency option was set to medium and other parameters were set as follows: EASE score = 0.05, similarity term overlap = 3, similarity threshold = 1.0, initial group membership = 3, final group membership = 3, and multiple linkage threshold = 0.50. These data were then integrated with EC expression data from TCGA. Specifically, data generated by the TCGA Research Network (https://cancergenome.nih.gov) were implemented to identify genes that may be regulated by miR-423 in ESCC. The TCGA mRNAseq and miRseq data for 183 EC samples were downloaded from FireBrowse (http://firebrowse.org/, data version 2016_01_28), as RSEM normalized counts and reads per million miRNAs mapped, respectively. Correlation among the expression of miR-423 (either 5p or 3p arm) and mRNAs was measured using the Gini correlation coefficient that was implemented in the RSGCC R package (Ma and Wang, 2012). The two-sided p-values of correlations were calculated by performing 1000 permutations and adjusted using false discovery rate (FDR) method. An FDR of 0.05 was considered significant. Genes that had a negatively correlated expression with miR-423 (i.e., had a negative coefficient) were identified. In this way, the verified targets that had a negatively correlated expression with miR-423 in EC were identified. In the next step, these genes were further screened for ESCC-related genes to further narrow down candidate targets that may contribute to ESCC pathogenesis. A list of ESCC-related genes was retrieved from the supplementary files of a previous study (Nariman-Saleh-Fam et al., 2016). These genes were linked to ESCC by either genetic association or expression (Nariman-Saleh-Fam et al., 2016).
Results
The selected SNPs
Figure 1 shows the genomic context of rs6505162 relative to the host genes. rs6505162 overlaps pre-miRNA sequence of hsa-mir-423, intronic region of NSRP1, and upstream region of hsa-mir-3184. rs1129186 is located in the 3′-UTR (relative to ENST00000244546) or the last exon (relative to ENST00000304611) of PEX6. This interaction was verified by crosslinking, ligation, and sequencing of hybrids (CLASH) method (Helwak et al., 2013) and is documented in the PolymiRTS database (Bhattacharya et al., 2014).
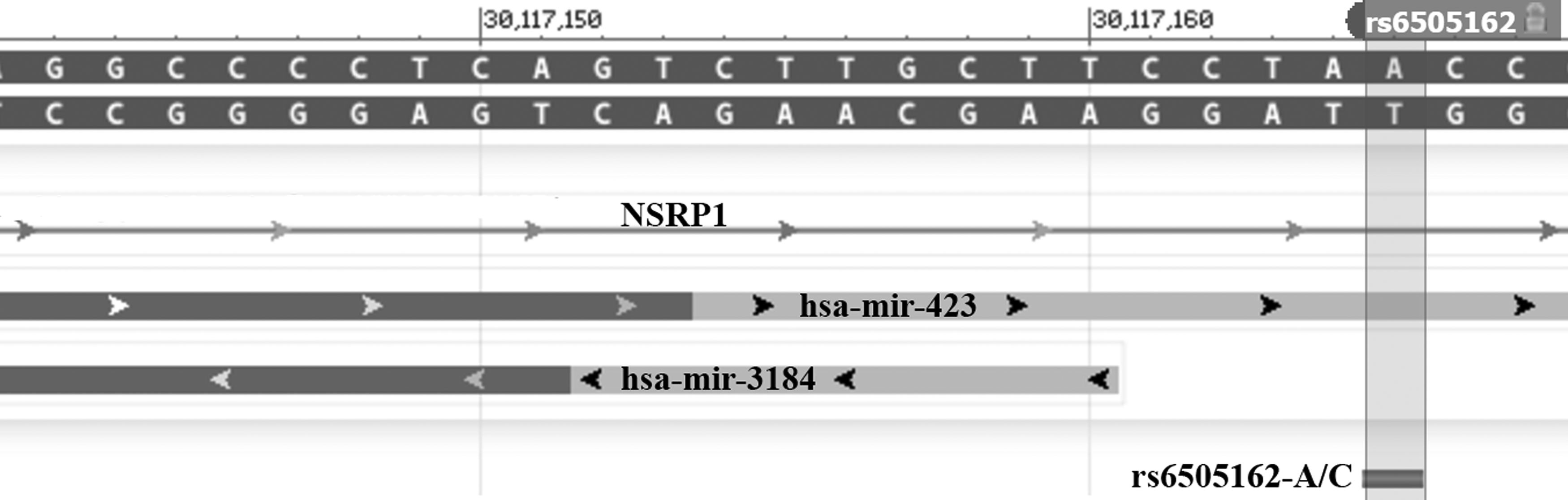
NCBI GeneView graphics of the genomic contexts of rs6505162. Genomic context of rs6505162 (chr17:30117143-30117167). This SNP overlaps pre-miRNA sequence of hsa-mir-423, intronic region of NSRP1 and upstream region of hsa-mir-3184. SNP, single-nucleotide polymorphism.
Restriction enzyme digestions
Figure 2A shows the recognition and the cleavage site of RsaI in the PCR product of miR-423. Digestion of miR-423 PCR product with RsaI resulted in a 143 bp DNA fragment for the AA genotype, three fragments (143, 124, and 19 bp) for the CA genotype, and two fragments (124 and 19 bp) for the CC genotype (Fig. 2A). However, the small fragment (19 bp) usually exits off the bottom end and it did not appear on the gel. Figure 2B shows an example of agarose gel electrophoresis of miR-423 digestion for three unknown samples along with digestion controls and NTC. Digestion of PEX6 PCR product with XhoI resulted in a 145 bp fragment for the TT genotype, three fragments (145, 122, and 23 bp) for the CT genotype and two fragments (122 and 2323 bp) for the CC genotype (Fig. 3A). As in miR-423, the smallest fragment (i.e., 2323 bp) did not appear on the gel. Figure 3B represents agarose gel electrophoresis of PEX6 digestion for three unknown samples along with digestion controls and NTC. Figures 2C and 3C illustrate the results of Sanger sequencing for the regions encompassing the studied SNPs in the six samples.
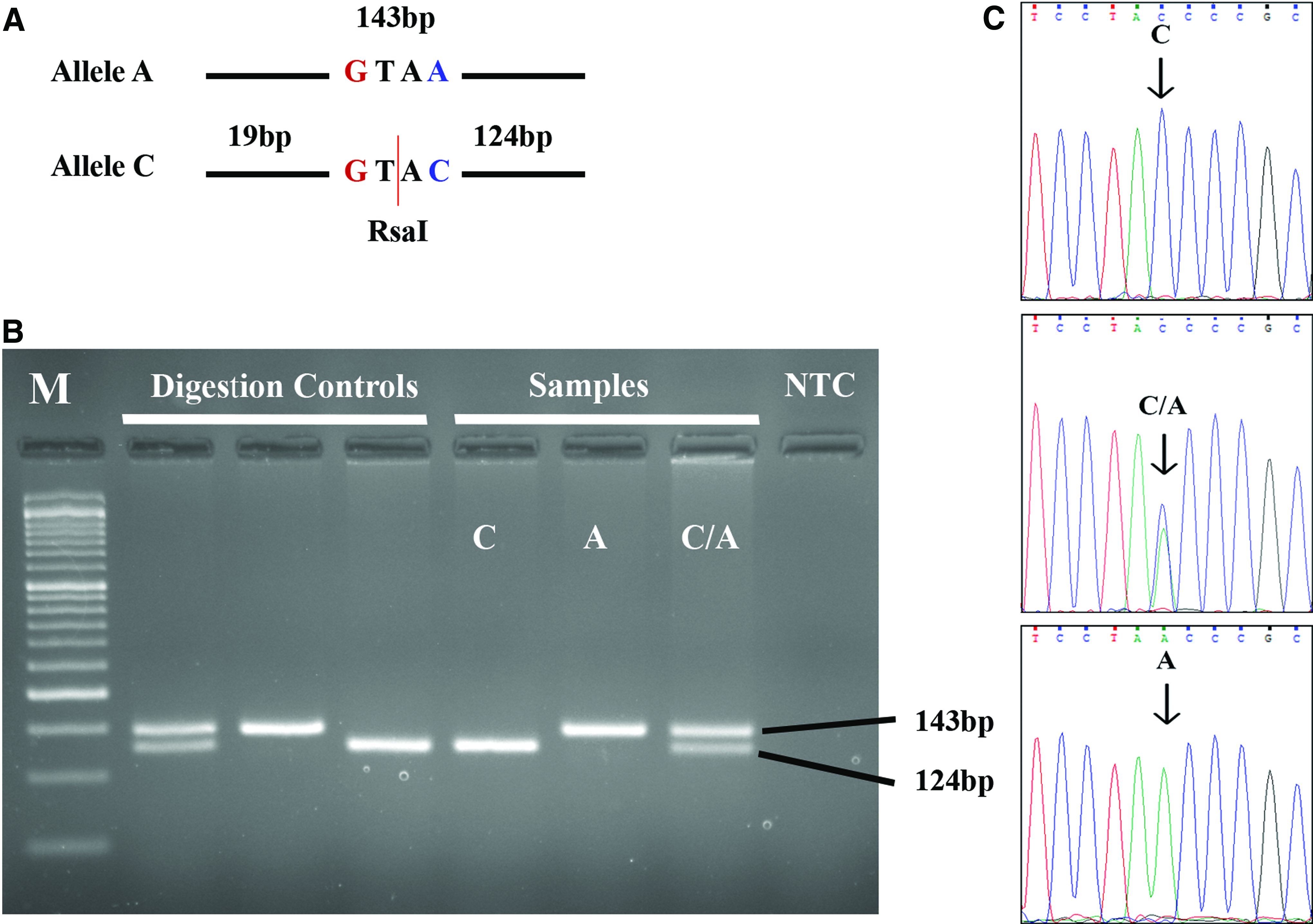
Restriction site and examples of genotyping results for miR-423 rs6505162.
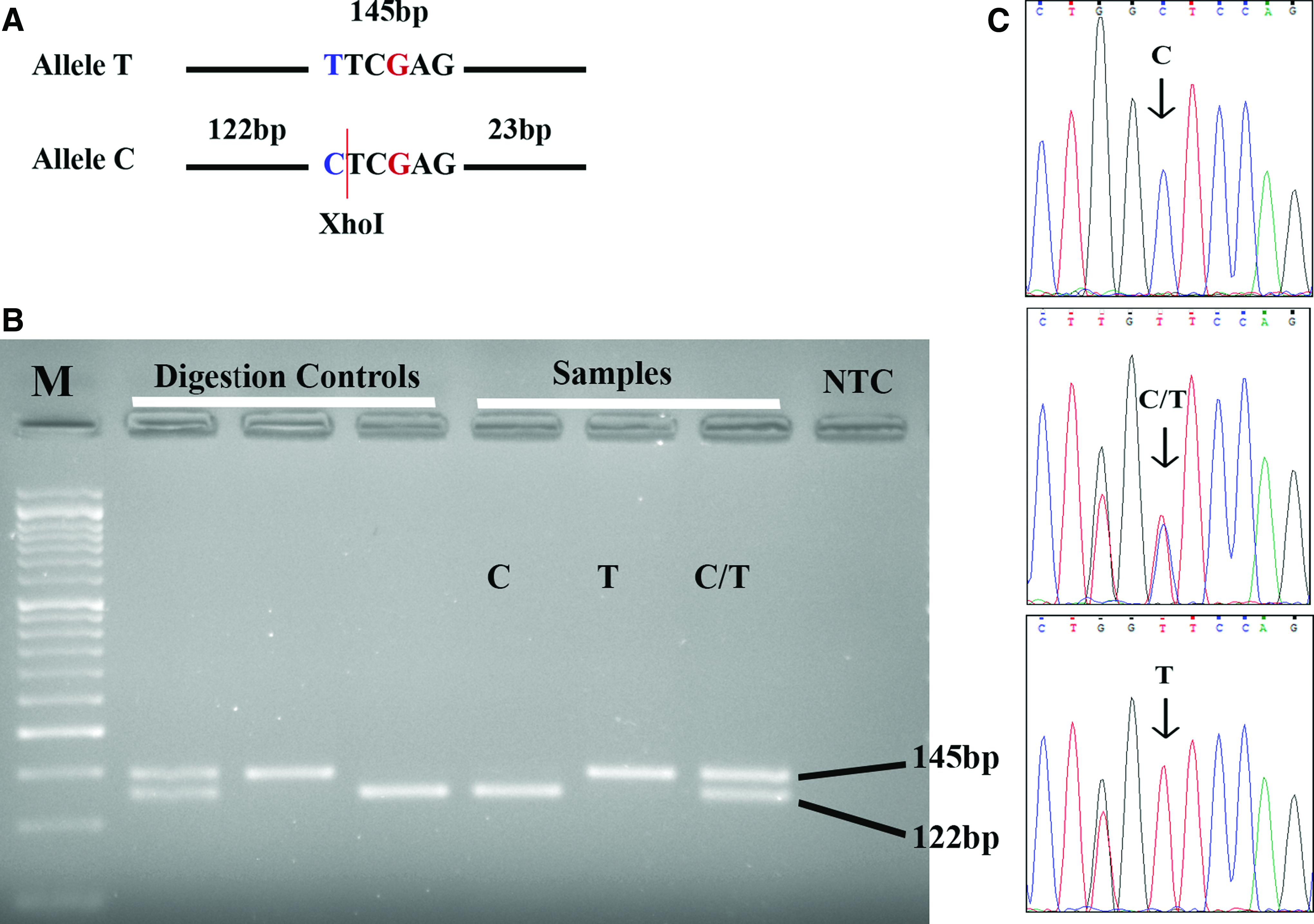
Restriction site and examples of genotyping results for PEX6 rs1129186.
miR-423 rs6505162 and ESCC risk
Genotype frequencies of miR-423 rs6505162 showed nonsignificant deviation from the Hardy-Weinberg equilibrium among members of the control group, patients, or all subjects (p-values were 0.2918, 0.2586, and 0.7464, respectively). The genotype frequency distributions of the studied SNPs are shown in Table 2. The frequency of the minor allele (A) of this SNP was 0.325 among the control group members, 0.247 among patients, and 0.294 for all participants. Logistic regression analysis revealed that miR-423 rs6505162 was associated with ESCC assuming codominant, recessive, and log-additive models. In the codominant model with the CC genotype as a reference, subjects carrying the AA genotype had a significantly decreased risk of ESCC (AA vs. CC, OR [95% CI]: 0.32 [0.15-0.69], p-value: 0.0076). In the recessive model with the CC+CA genotypes as reference, individuals with the AA genotype were found to have a decreased risk of ESCC (AA vs. CC+CA, OR [95% CI]: 0.35 [0.16-0.73], p-value: 0.0027). Moreover, results were also significant assuming the log-additive model (OR [95% CI]: 0.69 [0.52-0.91], p-value: 0.0084). According to the lowest AIC value, the recessive model had the best fit to the data.
AIC, Akaike's information criteria; CI, confidence interval; OR, odds ratio.
In silico analyses of miR-423 rs6505162
This SNP is located in the pre-miRNA sequence of hsa-mir-423 (MI0001445) that encodes two mature miRNAs (hsa-miR-423-5p and hsa-miR-423-3p) (Fig. 1). Theoretically, the SNP in pre-miRNA sequence can affect miRNA processing and, thereby, alter expression of mature miRNAs. This in turn may lead to dysfunction of those biological processes that are normally regulated by these mature miRNAs. Totally, 287 and 221 unique genes were identified as experimentally verified targets of miR-423-5p and miR-423-3p, respectively (these genes are accessible from the miRTarBase server). FAC analyses of miR-423 targets revealed 8 and 10 enriched clusters of biological processes (Table 3 and Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/gtmb). Expression correlation analysis showed that 125 verified target genes were negatively correlated with miR-423 (either 5p or 3p arm) in terms of expression levels in EC (Supplementary Table S2). Screening the genes for those that were already related to ESCC showed that CSTB was linked to ESCC. The expression levels of miR-423 and CSTB were negatively correlated (coefficient: −0.248, p-value: 0, Supplementary Table S2).
Enriched clusters of biological processes are shown in the order of decreasing enrichment scores. For each cluster one representative biological process is shown. Please refer to Supplementary Table S1 for the complete details.
PEX6 rs1129186 and ESCC risk
No significant departure from the Hardy-Weinberg equilibrium was observed for PEX6 rs1129186 genotypes among the control group members, the patients, or all subjects (p-values were 0.8108, 0.1423, and 0.5171, respectively). The frequency of the minor allele (T) of this SNP was 0.406 among members of the control group, 0.4 among patients and 0.404 overall. The results of logistic regression analysis showed that PEX6 rs1129186 was not associated with ESCC under any analyzed genetic model (Table 2).
Discussion
Increasing evidence has proposed that functional polymorphisms influencing miRNA regulatory network may predispose individuals to a variety of cancers, especially ESCC (Zhou et al., 2013; Shen et al., 2016). Although Iran is considered as a high-risk region for ESCC (Mahboubi et al., 1973), little is currently known about the role of genetic components, especially miRNA-related variations in determining susceptibility to disease. In this study, a genetic association was conducted to evaluate the contribution of two such variations to the susceptibility to ESCC in an Iranian cohort and it was shown that miR-423 rs6505162 was associated with a decreased risk of ESCC in the cohort.
The miR-423 rs6505162-A allele had a significant protective effect on ESCC in the cohort (Table 2). The association of this SNP with a risk of EC risk had also been reported in Caucasians and Black South Africans (Ye et al., 2008; Wang et al., 2013b). Ye et al. (2008) reported the first evidence of an association of this SNP with risk of EC in a Caucasian cohort. Although about 85% of patients in their study had EAC (Ye et al., 2008) (rather than ESCC in our study), the association was in the same direction with the present study, proposing a protective role for the rs6505162-A allele. The same SNP has also been reported to be associated with risk of ESCC in Black South Africans assuming dominant, recessive, and additive genetic models (Wang et al., 2013b). In contrast to the Caucasians (Ye et al., 2008) and Iranians (the present study), the major allele for rs6505162 in the Black South Africans was the A allele. However, this allele had a protective role compared with the C allele in all of these cohorts. In contrast to these findings, no evidence of an association with ESCC was found in a study on the Chinese Han population (Shen et al., 2016), suggesting that miR-423 rs6505162 may pose an ethnic-specific effect on the risk of ESCC. This SNP is located in miR-423 precursor and influences processing and expression of this miRNA (Zhao et al., 2015). Although this SNP is not located in the seed region (Fig. 1), it has been suggested that rs6505162 C>A substitution can affect not only the production of mature miR-423 (Zhao et al., 2015), but that it may also affect its targeting efficiency (Su et al., 2015). Using a dual luciferase assay, Su et al. (2015) indicated that the A allele can be more effective in suppressing the expression of PA2G4 than the C allele. Moreover, this miRNA was differentially expressed in ESCC tumor tissues compared with adjacent normal tissues by a recent study (Wang et al., 2013b), but rs6505162 genotypes were not correlated with miR-423 expression. Therefore, the exact mechanisms by which this SNP contributes to pathogenesis of ESCC have yet to be fully understood. In silico insights into the biological processes that are regulated by miR-423 were provided and target genes that were negatively correlated with miR-423 in terms of expression level were identified. Among these genes, CSTB was reported as downregulated in ESCC tumors compared with adjacent normal tissues both at mRNA (Shiraishi et al., 1998; Hu et al., 2010) and protein levels (Shiraishi et al., 1998; Zhang et al., 2011). Decreased expression of cystatin B (CSTB) that encodes one of the cysteine protease inhibitors was associated with a high frequency of lymph node metastasis and more advanced clinical stages. Cystatin B regulates the activity of lysosomal proteases (Shiraishi et al., 1998), and it is believed to protect against the proteases leaking from lysosomes (provided by RefSeq, Jul 2016). Therefore, there are pieces of evidence suggesting that miR-423 and CSTB may have a functional interaction in ESCC. First, interaction of miR-423 and CSTB transcript has been experimentally identified (Chou et al., 2016). Second, miR-423 and CSTP were negatively correlated in TCGA EC samples. Finally, miR-423 and CSTB were reported as upregulated and downregulated in ESCC, respectively (Shiraishi et al., 1998; Wang et al., 2013b). Future experiments will determine possible role of this interaction in pathogenesis of ESCC and whether rs6505162 may contribute to this process. Moreover, the influence of rs6505162 on expression level of miR-423 needs to be experimentally verified in ESCC. As it is apparent from Figure 1, rs6505162 is also located upstream of the miR-3184 precursor on the opposite strand. Hence, another alternative that should be considered in future studies is the potential influence of rs6505162 on miR-3184 expression and function.
PEX6 encodes a member of the AAA family of ATPases that is a predominantly cytoplasmic protein with a direct role in peroxisomal protein import (provided by RefSeq). Mutations in this gene cause peroxisome biogenesis disorders of complementation groups 4 and 6 (provided by RefSeq) (Zhang et al., 1999). It has been reported that PEX6 is downregulated in ESCC (Wang et al., 2013a) (NCBI GEO ID: GSE26886), suggesting that it has a role in pathogenesis of the disease. rs1129186 is a C>T synonymous substitution in PEX6 that, according to the GTEx portal (available at: http://gtexportal.org/home), is correlated with PEX6 expression in multiple tissues, including esophagus mucosa (GTEx Consortium, 2013). Interestingly, this SNP is located in a CLASH-verified target site for miR-149-5p (Helwak et al., 2013), a miRNA that is upregulated in ESCC (Guo et al., 2008). Based on this evidence, it is hypothesized that this miRNA-related polymorphism may be associated with risk of ESCC. In spite of evidence for the functional role of this SNP, the data of the current study did not suggest the contribution of this SNP to the risk of ESCC in the tested cohort. In conclusion, this study presents the first evidence for an association of miR-423 rs6505162 with risk of ESCC in an Iranian cohort and target genes and biological processes have been identified that are potentially influenced by rs6505162-induced changes in miR-423 function.
Footnotes
Acknowledgment
This work was supported by a grant from the research deputy of Tehran University of Medical Sciences (TUMS) (grant no. 25787).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.