
Abstract
Aims:
The glucocorticoid receptor (GR) and mineralocorticoid receptor (MR) play important roles in the hypothalamic-pituitary-adrenal axis and stress regulation. The current study aimed to address the genetic association and gene-gene interactions between GR and MR gene polymorphisms and aggressive behavior.
Methods:
A haplotype-based, case-control study was designed to examine the association between human MR and GR genes and aggressive behaviors, including robbery and intentional interpersonal injury, in a central south Chinese Han population. Eight well-studied polymorphisms of the MR gene (rs2070951, rs2871, rs5522, and rs5525) and the GR gene (rs10482605, rs6190, rs1800445, and rs6196) were genotyped using SNaPshot.
Results:
The MR and GR gene polymorphisms were not associated with aggressive behaviors, including in robbery and intentional interpersonal injury subgroups (p > 0.05). The frequency of haplotype TAAT of GR was significantly higher in the control group than that in the case group (p = 0.004, padj = 0.02). In addition, a two-way interaction between GR rs1800445 and MR rs2070951 and aggressive behavior was also found (p = 0.01).
Conclusions:
The haplotype TAAT of GR might be a protective factor against aggressive behavior, while gene-gene interactions between GR rs1800445 and MR rs2070951 might be a risk factor for aggressive behavior in the Central South Chinese Han population.
Introduction
A
The hypothalamic-pituitary-adrenal (HPA) axis plays a major role in the development, elicitation, and enhancement of aggressive behavior in animals and humans. Both acute and basal HPA axis activities have been shown to influence aggressive behavior in rats (Newman et al., 2005). In response to stress, the HPA system is activated by binding to corticotropin-releasing hormone (CRH) receptors (CRHR1 and CRHR2) and then stimulates the release of adrenocorticotrophic hormone (ACTH) and cortisol (Naughton et al., 2014; Grabe et al., 2010). Cortisol can also regulate the activity of the HPA axis by binding to mineralocorticoid receptor (MR) and glucocorticoid receptor (GR) (Kanitz et al., 2012; Ventura-Juncá et al., 2014). GRs mediate many of the effects of cortisol on target tissues via direct binding to hormone-responsive elements in the DNA and via interactions with other transcription factors, resulting in modulation of gene transcription (Gądek-Michalska et al., 2013). MRs are restricted to limbic areas, have a 10-fold higher affinity for cortisol, and play a role in tonic inhibition of the HPA axis (Gądek-Michalska et al., 2013).
A recent study suggested that polymorphisms of the GR gene are major factors in the variability of HPA activity (Wagner, 2014). Four common GR gene polymorphisms (rs6195(N363S), rs6189-6190(ER22/23EK), rs41423247(BclI) and rs6198) have been reported to be associated with variable sensitivity to glucocorticoids (GC) (Kumstaa et al., 2008; Moraitis et al., 2016; van Rossum et al., 2003; Stevens et al., 2004; Varricchio and Migliaccio, 2014). Given this evidence, the functional relevance of these gene variants for glucocorticoid sensitivity seems likely. However, knowledge on the influence of these variants on aggressive behavior is limited. Genetic variation of the MR might be another factor underlying individual susceptibility to stress-related disorders. Recently, MR rs5522 and rs2070951 were reported to change expression or cause dysfunction of the MR protein (DeRijk et al., 2006; Bortoluzzi et al., 2014). Therefore, it is of great importance to elucidate the individual specificity of GR and MR variants, which contribute to HPA axis changes.
To date, no evidence supporting the genetic association between GR and MR variations and aggressive behavior has been reported. To identify any variations in GR and MR related to aggressive behavior, we analyzed eight single-nucleotide polymorphisms (SNPs) in GR and MR genes in violent criminals with aggressive behavior, including robbery (aggressive behavior toward property) and intentional interpersonal injury (physically aggressive behavior toward other people), in a central south Chinese Han population.
Materials and Methods
Subjects
All procedures involving human participants were conducted in accordance with the ethical standards of the Changsha Medical University, the 1964 Declaration of Helsinki, and its latter amendments. Informed consent was obtained from all participants included in the current study. A total of 291 violent criminals (mean ± standard deviation age = 34 ± 11.2 years) with aggressive behavior toward property or other people were recruited at a prison in Changsha, China. Of them, 136 (46.7%) were convicted of robbery, 134 (46.0%) of intentional interpersonal injury, and 21 (7.3%) for other types of aggression against persons or property. Detailed information on each violent criminal is listed in Table 1. In addition, 300 unrelated healthy control individuals (mean ± standard deviation age = 27 ± 15 years) without a crime record were included after undergoing clinical examination at the First Affiliated Hospital of Changsha Medical University. All cases and controls were Han Chinese males.
SNP selection and genotyping
Eight SNPs in MR (rs2070951, rs2871, rs5522, and rs5525) and GR (rs10482605, rs6190, rs1800445, and rs6196) that have been well-studied and associated with psychiatric disorders were selected. The standard phenol-chloroform method was used for extracting genomic DNA. The multiplex polymerase chain reaction (PCR) was carried out on an ABI Veriti Thermal Cycler (Applied Biosystems, Foster City, CA). Genotyping of MR and GR polymorphisms was conducted by using SNaPshot (Applied Biosystems).
Statistical analyses
Allele and genotype frequencies of MR and GR polymorphisms were compared between criminals and controls using a two-sided Fisher's exact test. Hardy-Weinberg equilibrium calculations, allele and genotype frequencies, haplotype analyses, and p-value permutations were conducted using SHEsis software (http://analysis.bio-x.cn/myAnalysis.php). Logistic regression was performed to test for associations between the polymorphisms and aggressive behavior, as well as associations between aggressive behavior subgroups (intentional interpersonal injury and robbery). If a significant relationship was found, stringent Bonferroni correction was carried out to adjust the p-value (padj) in multiple comparisons. Gene-gene interaction analysis was assessed by multifactor dimensionality reduction (MDR; version 1.0). Statistical significance was set at p < 0.05.
Results
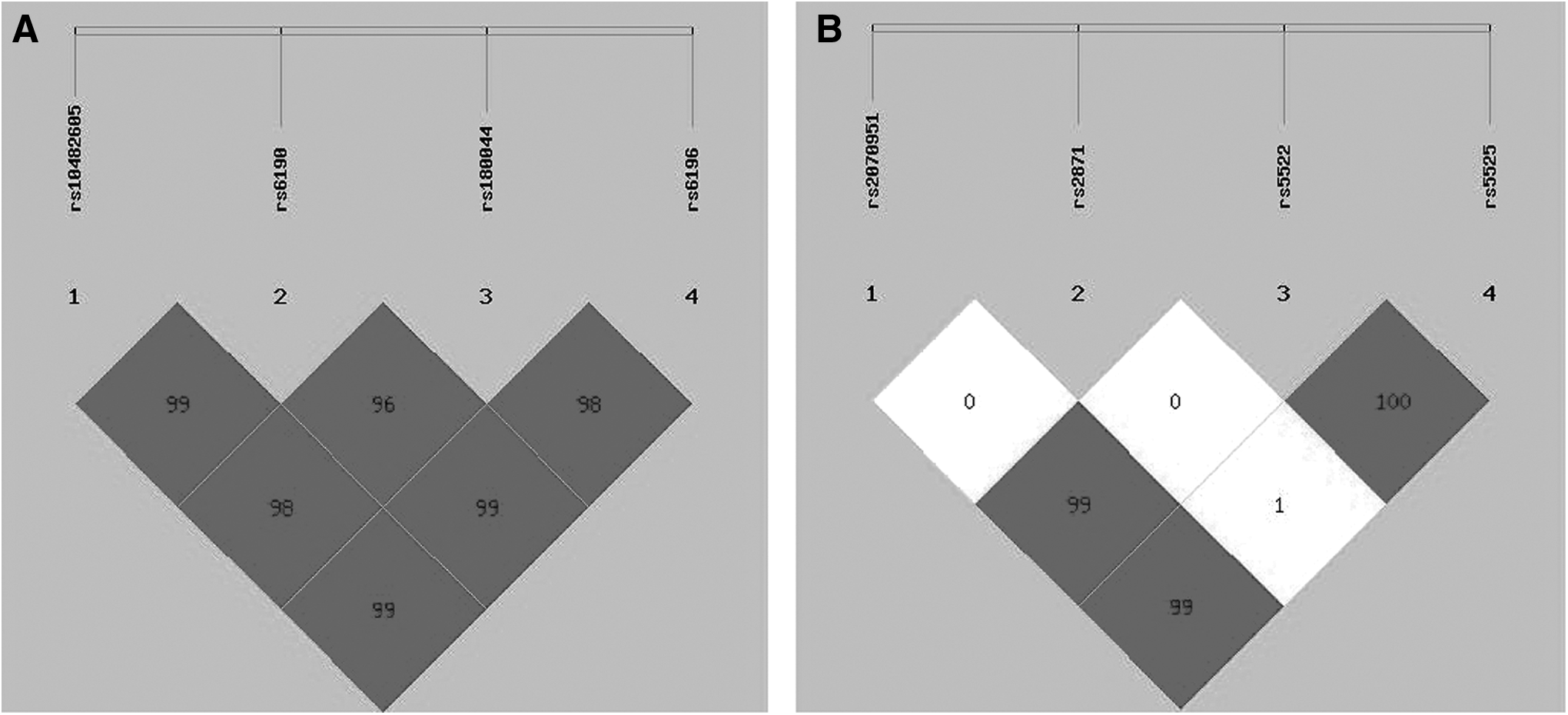
With an average call rate of 99.8%, the eight MR and GR SNPs were successfully genotyped in a total of 591 subjects. All variants (except for NR3C1 rs1800445 in the control group, p = 0.005) in cases and controls were in Hardy-Weinberg equilibrium (p > 0.05). The minor allele frequencies of the eight SNPs were similar to the data from the International HapMap Project (Chinese Han population). Four GR SNPs (rs10482605, rs6190, rs1800445, and rs6196) and three MR SNPs (rs2070951, rs5522, and rs5525) were in strong linkage disequilibrium (D′ > 0.85 and D′ > 0.85, Fig. 1A, B, respectively). However, no significant difference in allelic or genotypic frequency distributions for any of the MR and GR SNPs was found (p > 0.05; Table 2).
p-value was calculated using Fisher's exact test. padj: The Bonferroni's correction was carried out to correct the p-value; “—” not significant.
OR [95%CI] was estimated by logistic regression analysis.
HWE, Hardy-Weinberg equilibrium test for case and control.
OR, odds ratio, CI, confidence interval.
Comparison of the two aggressive behavior subgroups (robbery and intentional interpersonal injury) with controls showed no significant association between the MR and GR SNPs and aggressive behavior subgroups even after Bonferroni correction (p > 0.05; Table 3). Evaluation of the risk of alcohol abuse, smoking, economic status, and education conditions related to aggressive behavior in general, and each behavioral subgroup showed no significant association between GR and MR SNPs and aggressive behavior (p > 0.05; Supplementary Tables S1-S4; Supplementary Data are available online at www.liebertpub.com/gtmb).
p-value was calculated using Fisher's exact test.
The Bonferroni's correction was carried out to correct the p-value. “—” not significant.
OR [95%CI] was estimated by logistic regression analysis.
Haplotype analyses of MR and GR SNPs were conducted to assess their association with aggressive behavior susceptibility. As shown in Table 4, five major haplotypes were identified by MR SNPs (rs2070951, rs2871, rs5522, and rs5525), and four were identified by GR SNPs (rs10482605, rs6190, rs1800445, and rs6196). Results revealed that HapCTTG (rs2070951C-rs2871T-rs5522T-rs5525G) of MR and HapTGAT (rs10482605T-rs6190G-rs1800445A-rs6196T) of GR were the most common haplotypes in both cases and controls (HapCTTG: criminals = 0.628, controls = 0.601; HapTGAT: criminals = 0.507, controls = 0.512). No statistical difference was observed in these two haplotypes between cases and controls both in overall aggressive behavior group and behavioral subgroups (p > 0.05; Table 4). However, the distribution of HapTAAT of GR in the controls was significantly higher than that in the cases (p = 0.004, padj = 0.02, odds ratio = 0.09, 95% confidence interval = 0.01-0.71), indicating a protective haplotype for aggressive behavior (Table 4). High-dimensional gene-gene interaction analysis showed that the highest testing-balanced accuracy (52.57%) was found for the GR rs1800445-MR rs2070951 interaction, which revealed a two-way interaction between MR and GR gene polymorphisms and aggressive behavior (cross-validation consistency, 9/10; p = 0.01; Supplementary Table S5 and Fig. 2).
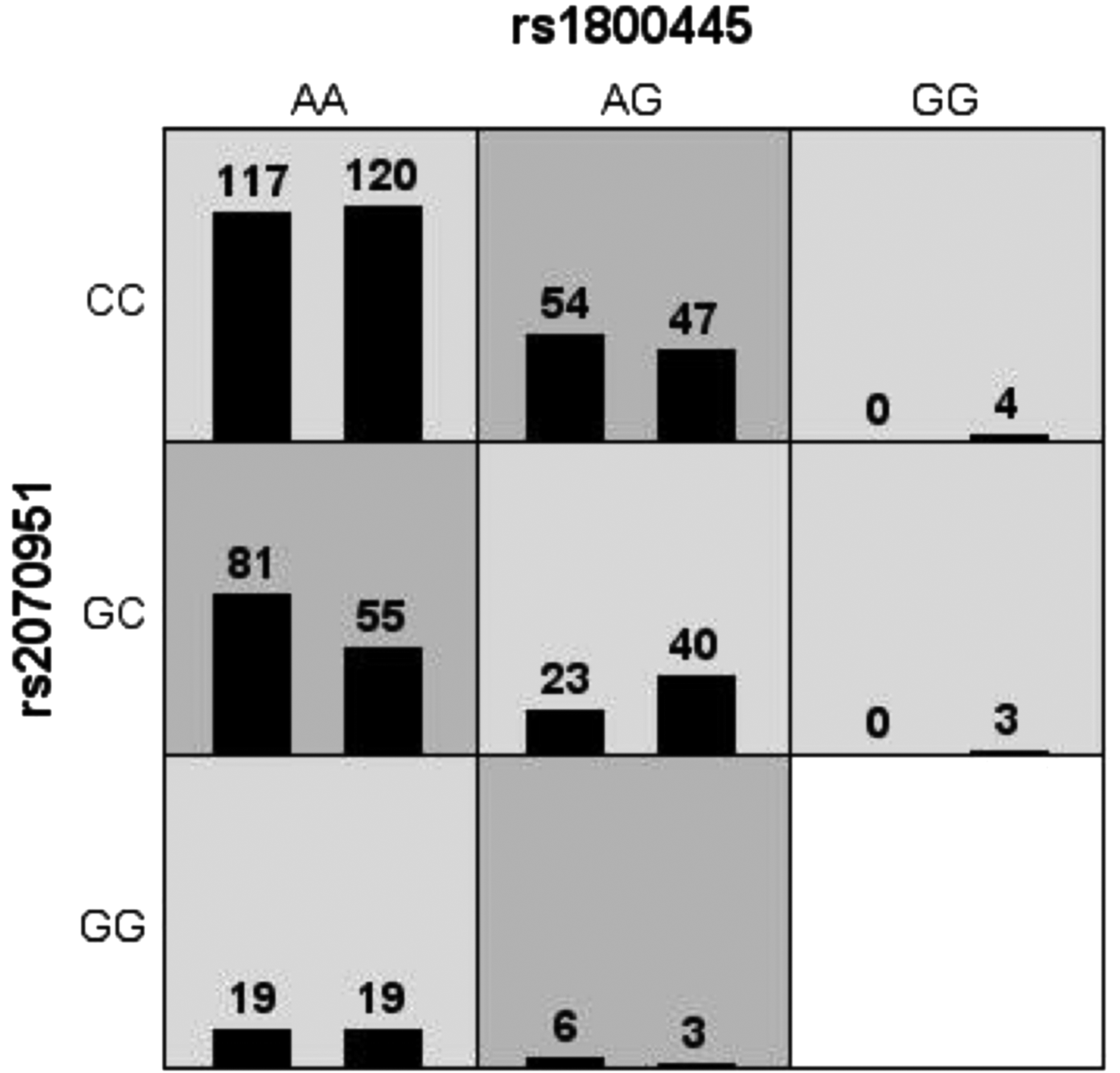
Interaction between GR rs1800445 and MR rs2070951 in aggressive behavior in a central south population. High-risk genotypes are in dark gray cells, and low-risk genotypes are in light cells. The percentages of patients (left bar in the boxes) and control subject (right bar in boxes) are shown for each two-locus genotype combination.
Haplotype structures of MR were rs2070951 (C/G), rs2871 (C/T), rs5522 (C/T), and rs5525 (A/G). Haplotype structures of GR were rs10482605(C/T), rs6190(A/G), rs1800445(A/G), and rs6196(T/C).
p value was calculated using Fisher's exact test. padj: The Bonferroni's correction was carried out to correct the p-value; “—” not significant.
OR [95%CI] was estimated by logistic regression analysis.
R group, robbery group; I group, intentional injury group; Non-R, crimes without robbery; Non-I, crimes without intentional injury.
Discussion
The current study evaluated the genetic association between four GR SNPs (rs10482605, rs6190, rs1800445, and rs6196) and four MR SNPs (rs2070951, rs2871, rs5522, and rs5525), as well as their gene-gene interactions and aggressive behavior in a central south Chinese Han population. Results revealed a protective role for HapTAAT of GR in aggressive behavior and a two-way interaction between GR rs1800445 and MR rs2070951 against aggressive behavior. To our knowledge, this is the first study to investigate the association between GR and MR SNPs and aggressive behavior in a central south Chinese Han population.
Large number of genetic variants of GR has been reported to be related to glucocorticoid resistance, heightened cortisol levels, negative HPA-axis feedback, and stress-induced activation of the HPA system (Koper et al., 1997; Lian et al., 2012; Streit et al., 1970; Harris et al., 2013). Previous studies have reported associations between GR polymorphisms and stress-related disorders. Bet et al. (2009) confirmed that GR polymorphisms and childhood adversity were associated with depression and a gene-environment interaction between common variants of the GR gene and childhood adversity was also detected. Wüst et al. (2004) investigated the effect of rs41423247 (BclI), rs6195 (N363S), and rs6189/6190 (ER22/23EK) of GR on cortisol and ACTH responses in healthy males and found that male N363S AG carriers displayed the highest ACTH (main effect genotype) and total cortisol responses, whereas male BclI GG carriers showed diminished responses (Kumsta et al., 2007). However, no significant association was detected between five polymorphisms of GR and depressive psychosis (Feng et al., 2000). To the best of our knowledge, haplotype-based association analysis of the role of GR in HPA axis changes, and aggressive behavior has not yet been reported.
In our study, the frequency of HapTAAT of GR in our sample was lower than that in a Belgian population with major depression disorder (0.01 vs. 0.005), but higher than that in a Swedish population with major depression (0.005 vs. 0.000) (West et al., 2006). Moreover, HapTAAT of GR was found significantly less often in cases compared with healthy controls (padj = 0.02). The GR gene maps to chromosome 5q31. The rs10482605 was found in nearly complete linkage with rs6819 in exon 9beta. Also, the C allele of the rs10482605 showed reduced transcriptional activity (Kumsta et al., 2009). Rs6190 (R23K) has been reported to be located in exon 2 and leads to an R23K mutation, which resulted in resistance to glucocorticosteroids (Feng et al., 2000), while rs6196 (N766 N), located in exon 9, causes a synonymous mutation in the NR3C1 protein (Supriyanto et al., 2011). These results indicate that aggressive behavior might be influenced by multiple polymorphic sites rather than a single site.
Interestingly, significant association was detected between HapTAAT of GR and aggressive behavior, but not between any of the MR polymorphisms or haplotypes examined and aggression. Studies (Juruena et al., 2013; Perroud et al., 2011) have shown that GR activation is necessary for HPA feedback regulation in the presence of high levels of glucocorticoids. Baseline levels of glucocorticoids are inversely related with aggressiveness in various species (Baugh et al., 2013). In contrast, increasing glucocorticoid levels in rodents may promote aggressive behavior by a fast feed-forward mechanism (Vilela et al., 2013). Thus, the indication of possible molecular mechanisms underlying the enhanced negative feedback effect via GR must be investigated in further studies.
Herein, no significant association was found between MR (NR3C2) and criminals with aggressive behavior in the present central south Chinese Han population. In fact, the functional variants of MR have already been characterized. For example, a 75-kDa protein encoded by a splice variant of MR was found to be a ligand-independent transactivator capable of recruiting coactivators (Zennaro et al., 2001). Variants associated with transcription resulting from the start codons in exon 2 were reported to have impacted the transcriptional activities of MR-A and MR-B proteins (Pascual-LeTallec et al., 2004), and MR rs2070951 (−2G/C), located two nucleotides before the translation site of exon 2, was reported to change transactivational capacity in vivo. Also, C-allele carriers have an increased MR expression (van Leeuwen et al., 2010). Importantly, MR rs5522 is located in exon 2. It substitutes isoleucine (A) for valine (G) in the N-terminal of the MR protein and was identified to increase cortisol secretion during Trier Social Stress Test (TSST) (Varricchio and Migliaccio, 2014). These results provide evidence that MR variants are causative of individual stress responsivity, affecting vulnerability to disorders, such as aggressive behavior. Although, no significant association was found between the MR variants examined in the current study and aggressive behavior, we could not exclude the impact of MR on the pathogenesis of aggressive behavior since this receptor is widely known to mediate corticosterone signaling. These MR SNPs may help regulate gene expression by associating with other SNPs. In particular, previous studies have reported an association between MR polymorphisms and other psychotic disorders except for aggressive behavior. In these cases, diseases heterogeneity might also explain differences between their data and ours. Follow-up studies with a larger sample population and other HPA axis genes to understand the complex relationship between genetic and environmental factors in the development of aggressive behavior are required.
MDR analysis is a method for detecting and characterizing high-order gene-gene interactions in case-control studies with relatively small sample numbers (Liu et al., 2011). Herein, MDR analysis detected a significant interaction between GR rs1800445 and MR rs2070951. To our knowledge, this is the first study to detect genetic interactions related to aggressive behavior in a Chinese population. This interaction indicates that the pathogenesis of aggressive behavior is very complex considering these two genes are involved in the same biological pathway.
Several limitations of this research must be considered. In particular, the sample size recruited in the current study was relatively small. To confirm the role of HPA axis gene variants in the development of aggressive behavior, studies including a larger number of subjects are necessary. Second, other HPA axis genes, such as CRH, CRHR1, and CRHR2, may also play roles in the regulation of ACTH and cortisol release and should, therefore, be investigated as well. Finally, our study was predominantly performed in a Chinese Han population with substance dependencies, and it is not clear whether the current results can be generalized or compared to other populations or patient groups.
Conclusions
The present results suggested that HapTAAT of GR and the gene-gene interaction between GR rs1800445 and MR rs2070951 may contribute to the development of aggressive behavior in a central south Chinese Han population. Our results have important implications for future research on the role of HPA axis function in aggressive behavior susceptibility.
Footnotes
Acknowledgments
We are grateful to all the individuals who participated in this study. This study was funded by the Foundation of the Health Department of Hunan Province, China (B2016096), Foundation (15C0513, 11C0141) and Key Foundation (15A023, 16A027) of the Education Department of Hunan Province, National Science Foundation of Hunan Province (2015JJ6010), Basic Research Program (2015JC3059) and Research Program (2014FJ6072) of the Science and Technology Commission Foundation of Hunan Province, and the Construct Program of the Key Discipline in Hunan province.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.