
Abstract
Background:
Single nucleotide polymorphisms of the IL10 gene have been linked to the occurrence of autoimmune liver disease.
Methods:
We performed a meta-analysis to assess the association between three IL10 promoter polymorphisms (rs1800896, rs1800871, and rs1800872) and the risk of autoimmune hepatitis, primary biliary cholangitis, and primary sclerosing cholangitis.
Results:
In total, 1420 articles were initially identified through database retrieval. After screening, seven eligible articles were ultimately included in the meta-analysis. A fixed-effect model was used for all Mantel-Haenszel statistics due to the absence of large between-study heterogeneity (all I2 < 50%, p > 0.1). No association between any of the studied polymorphisms and risk of autoimmune liver disease was detected in the allele, homozygote, heterozygote, dominant, recessive, or carrier genetic models (passociation > 0.05). Potential publication bias was excluded using Begg's and Egger's tests. Similar negative results were observed in subgroup analyses and in an analysis of the three haplotypes of rs1800896/rs1800871/rs1800872 (G/C/C, A/C/C, and A/T/A).
Conclusion:
Our meta-analysis strongly suggests that the IL10 rs1800896, rs1800871, and rs1800872 polymorphisms are not associated with the risk of autoimmune liver disease.
Introduction
A
Interleukin 10, encoded by IL10 located on chromosome 1 (1q31-32) (Eskdale et al., 1997), is a multifunctional cytokine that has been implicated in various biological events and diseases (Walter, 2014; Saxena et al., 2015). Single nucleotide polymorphisms (SNPs) are variations in a single nucleotide within a gene exon or intron and may result in alteration of a protein's structure or function (Benitez et al., 2017; Matsuda, 2017). Three commonly occurring human IL10 SNPs, rs1800896 (-1082A/G), rs1800871 (-819 T/C), and rs1800872 (-592 T/C), are situated within the gene's promoter region (Matsushita et al., 2002; Liu et al., 2015; Liu and Zheng, 2016). These IL10 SNPs are reportedly associated with susceptibility to several clinical diseases, such as tuberculosis (Liu et al., 2015), type 2 diabetes mellitus (Tarabay et al., 2016), liver cancer (Wei et al., 2011), or autoimmune thyroid disease (AITD) (Jung et al., 2016). In the present study, we performed a meta-analysis to investigate the genetic relation between those IL10 SNPs and the risk of autoimmune liver disease. To our knowledge, no such meta-analysis has been reported previously.
Materials and Methods
Database retrieval
Based on preferred reporting items for systematic reviews and meta-analyses (PRISMA) (Moher et al., 2009), we carried out retrieval from the PubMed, Embase, Scopus and Web of Science (WOS) databases on November 23, 2017, without any restriction of publication language, region, and article type. The search terms used are listed in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/gtmb).
Article screening
After excluding duplicate articles by Endnote X7 software (Thomson Corporation, Thomson ResearchSoft), we further removed articles using the following exclusion criteria: (1) cell or animal experiments; (2) review articles or case studies; (3) meeting abstracts; (4) other genes/diseases; (5) meta-analyses; (6) unselected SNPs; (7) lack of genotype data. The included studies should report the genetic association between IL10 SNPs and autoimmune liver disease risk, and contain sufficient data of genotype frequency distribution in case and control groups.
Basic information
The specific data were separately collected from the eligible articles by three investigators and summarized as basic information, including first name, publication year, ethnicity, SNP, case/control number, specific type of autoimmune liver disease, age, gender, genotyping assay, source of control, and p-value for Hardy-Weinberg equilibrium (pHWE) in control group. The genotype frequency distribution was considered to be in Hardy-Weinberg equilibrium, when pHWE was larger than 0.05.
Quality assessment
We also assessed the quality of the studies using the Newcastle-Ottawa Scale (NOS) (Stang, 2010; Ye et al., 2016). The following items were evaluated: (1) Case Definition; (2) Representativeness of cases; (3) Selection of controls; (4) Definition of controls; (5) Important factors of comparability; (6) Other factors of comparability; (7) Secure record of exposure; (8) Same method of ascertainment; (9) Nonresponse rate. NOS scores larger than 7 indicate high quality. We will fully discuss the uncertain or controversial points.
Statistical analysis
STATA software (version 12.0; Stata Corporation, College Station, TX) was used for analysis. Pheterogeneity values derived from Cochran's Q statistic >0.1 or I2 value <50% indicated the application of a fixed-effect model. Otherwise, a random-effect model was used.
Odds ratios, 95% confidence intervals, and passociation were obtained from Mantel-Haenszel statistics under the allele (G vs. A for rs1800896, C vs. T for rs1800871, C vs. A for rs1800872), homozygote (GG vs. AA, CC vs. TT, CC vs. AA), heterozygote (AG vs. AA, TC vs. TT, AC vs. AA), dominant (AG+GG vs. AA, TC+CC vs. TT, AC+CC vs. AA), recessive (GG vs. AA+AG, CC vs. TT+TC, CC vs. AA+AC), and carrier (G vs. A, C vs. T, C vs. A) models. A statistical significant difference between case and control was considered, when passociation was less than 0.05.
Begg's and Egger's tests were used to assess the potential publication bias. p-Value for Begg's and Egger's tests larger than 0.05 indicated the absence of publication bias. Subgroup meta-analyses based on ethnicity (Caucasian or Asian), pHWE (>0.05 or <0.05), and disease type (AIH, PBC, or PSC) were also performed under the allele, homozygote, heterozygote, dominant, recessive, and carrier models, respectively.
Results
Identification of eligible case-control studies
Figure 1 shows our flow chart for identifying eligible case-control studies. In total, 1420 articles were initially identified through comprehensive retrieval from the PubMed (n = 228), Embase (n = 284), Scopus (n = 254), and WOS (n = 654) databases. We then excluded 515 articles as duplicates and another 896 articles based on our exclusion criteria. Evaluation of the full text of the remaining nine articles led to the removal of additional two articles due to unselected SNPs (Mitchell et al., 2001) or lack of genotype data (Ma and Qiu, 2004). As a result, seven articles (Zappala et al., 1998; Cookson et al., 1999; Bathgate et al., 2000; Donaldson et al., 2000; Matsushita et al., 2002; Chen et al., 2004; and Fan et al., 2005) were ultimately selected for meta-analysis. From the eligible articles, we obtained 11 case-control studies for rs1800896, 8 for rs1800871, and 9 for rs1800872. Table 1 and Supplementary Table S2 show the characteristics of the participants in the included studies and their genotype distributions. Notably, we found that the genotype distributions for some control groups of two studies were not in line with Hardy-Weinberg equilibrium (Supplementary Table S2, pHWE <0.05) (Bathgate et al., 2000; Matsushita et al., 2002). Summarized in Supplementary Table S3 are results from NOS system evaluations, which showed that all the included studies were of high quality (NOS score >8).
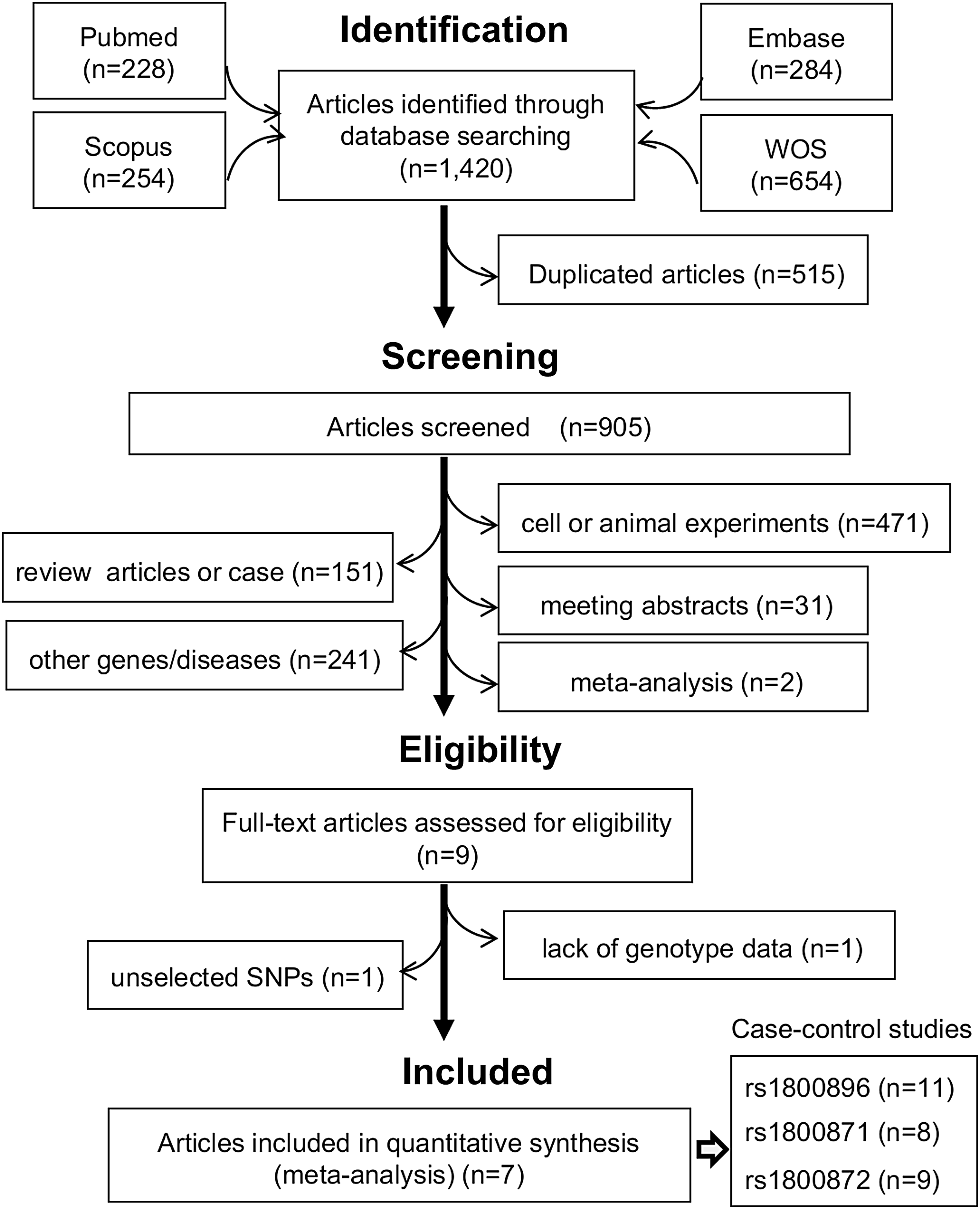
Selection process for eligible case-control studies. WOS, Web of Science.
The data of age are shown as mean ± standard deviation, or age range.
AIH, autoimmune hepatitis; F, female; M, male; NR, not reported; PBC, primary biliary cholangitis; PSC, primary sclerosing cholangitis; PCR, polymerase chain reaction; RFLP, restriction fragment length polymorphism; SNP, single nucleotide polymorphism; SSCP, single-strand conformation polymorphism; SSP, sequence specific primer.
IL10 rs1800896
In the first meta-analysis, we analyzed the relationship between the IL10 rs1800896 A/G SNP and the risk of autoimmune liver disease. Enrolled were 703 cases and 1870 controls. As shown in Supplementary Table S4, there was not a high degree of heterogeneity across the studies (all I2 < 50%, pheterogeneity >0.1), and a fixed-effect model was used for all Mantel-Haenszel statistics. Pooled data (Table 2) showed that there was no difference in the risk of autoimmune liver disease between case and control groups (allele G vs. A, passociation = 0.207; GG vs. AA, passociation = 0.388; AG vs. AA, passociation = 0.081; AG+GG vs. AA, passociation = 0.081; GG vs. AA+AG, passociation = 0.938; carrier G vs. A, passociation = 0.351).
CI, confidence interval; OR, odds ratio; passociation, p-value based on Mantel-Haenszel statistics of association test.
Subgroup analysis based on ethnicity (Caucasian or Asian), pHWE (>0.05 or <0.05), and disease type (AIH, PBC, or PSC) yielded similar results in all genetic models for all subgroups (Table 3, all passociation >0.05). The forest plot in Figure 2A shows the subgroup analysis based on disease type in a G versus A model, whereas Supplementary Figure S1 shows data for the ethnicity subgroup. These data indicate that IL10 rs1800896 is not linked to the risk of autoimmune liver disease.
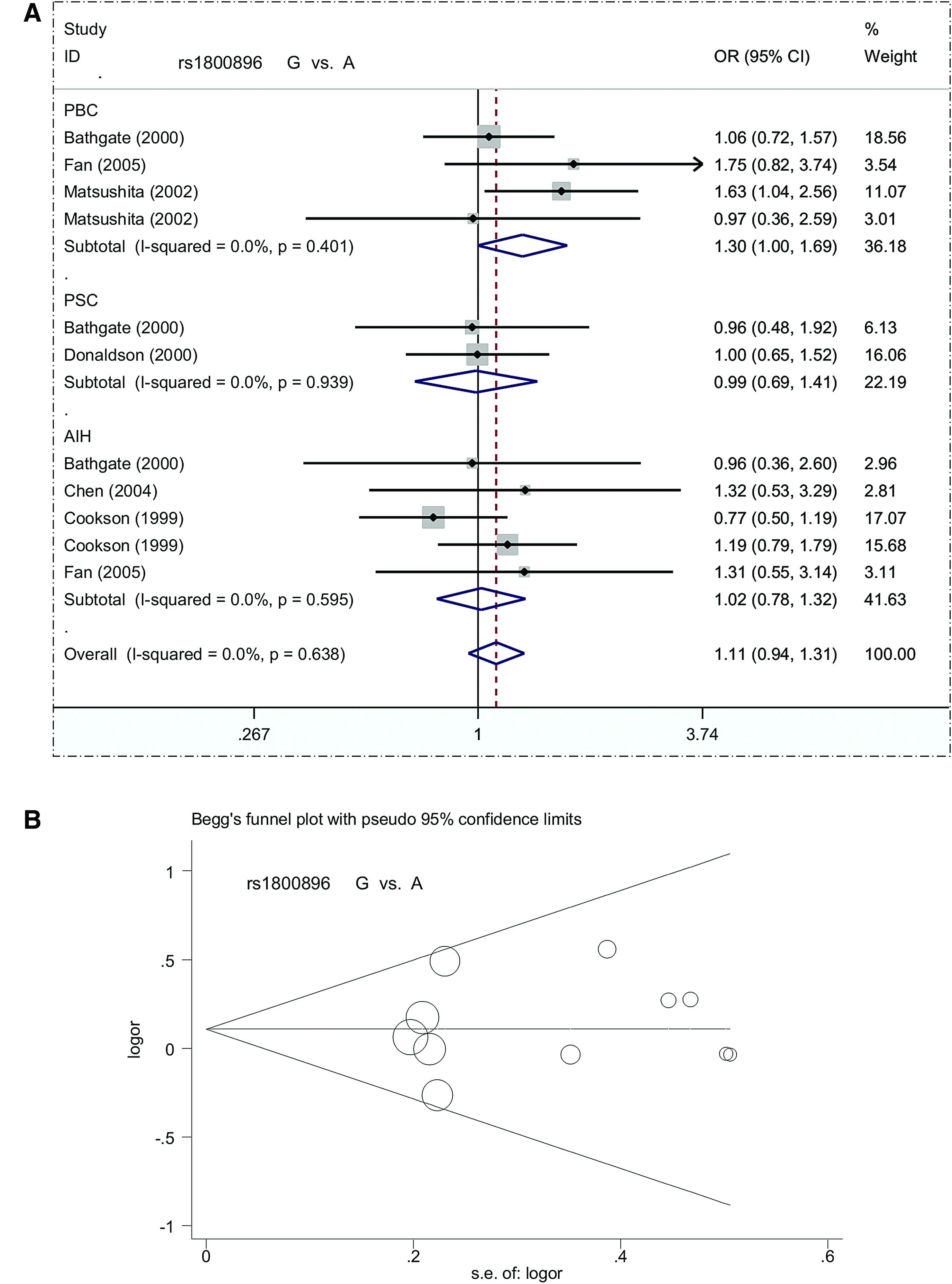
Forest plot and Begg's funnel plot for subgroup analysis of IL10 rs1800896 based on disease type under a G versus A model.
pHWE, p-value for Hardy-Weinberg equilibrium.
Begg's and Egger's tests showed an absence of potential publication bias (Supplementary Table S4, all pBegg >0.05, pEgger >0.05). The Begg's funnel plot in Figure 2B presents the pseudo 95% confidence limits in the G versus A model.
IL10 rs1800871
Our meta-analysis of IL10 rs1800871 included 8 case-control studies containing 424 cases and 400 controls. A fixed-effect model was used (all I2 = 0.0%, pheterogeneity >0.1) for this analysis (Supplementary Table S4). As with IL10 rs1800896, there was no significant difference between the cases and population-based control group under any genetic model (Table 2, all passociation >0.05). The subsequent subgroup analyses yielded similar negative results (Table 4, Fig. 3A, and Supplementary Fig. S2, all passociation >0.05). Begg's and Egger's tests showed no meaningful publication bias (Supplementary Table S4 and Fig. 3B, all pBegg >0.05, pEgger >0.05). This result provides no support for a positive correlation between IL10 rs1800871 and risk of autoimmune liver disease.
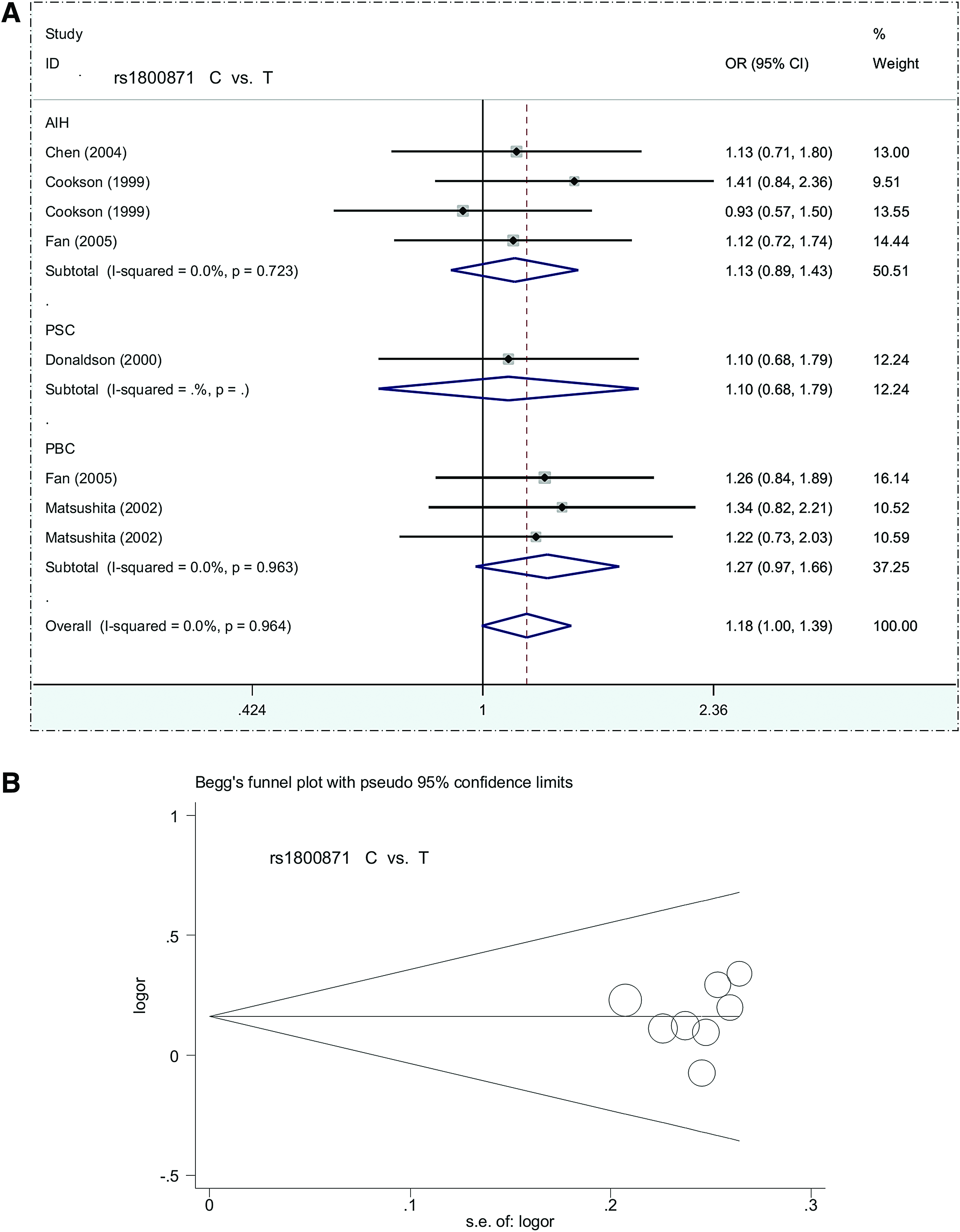
Forest plot and Begg's funnel plot for subgroup analysis of IL10 rs1800871 based on disease type under a C versus T model.
IL10 rs1800872
Nine case-control studies with 804 cases and 1005 controls were included in the meta-analysis of IL10 rs1800872. Again, a fixed-effect model was used (Supplementary Table S4, all I2 = 0.0%, pheterogeneity >0.1), and no significant difference was detected between the overall case and control groups (Table 2, all passociation >0.05). Similar negative results were observed in most subgroup analyses (Table 5, Fig. 4A, and Supplementary Fig. S3, passociation >0.05), apart from the subgroup of Asian (passociation = 0.036) and AIH (passociation = 0.046) in the CC versus AA genetic model. Begg's and Egger's tests indicated an absence of publication bias (Supplementary Table S4 and Fig. 4B, all pBegg >0.05, pEgger >0.05). Thus, IL10 rs1800872 seems not to be associated with the risk of autoimmune liver disease.
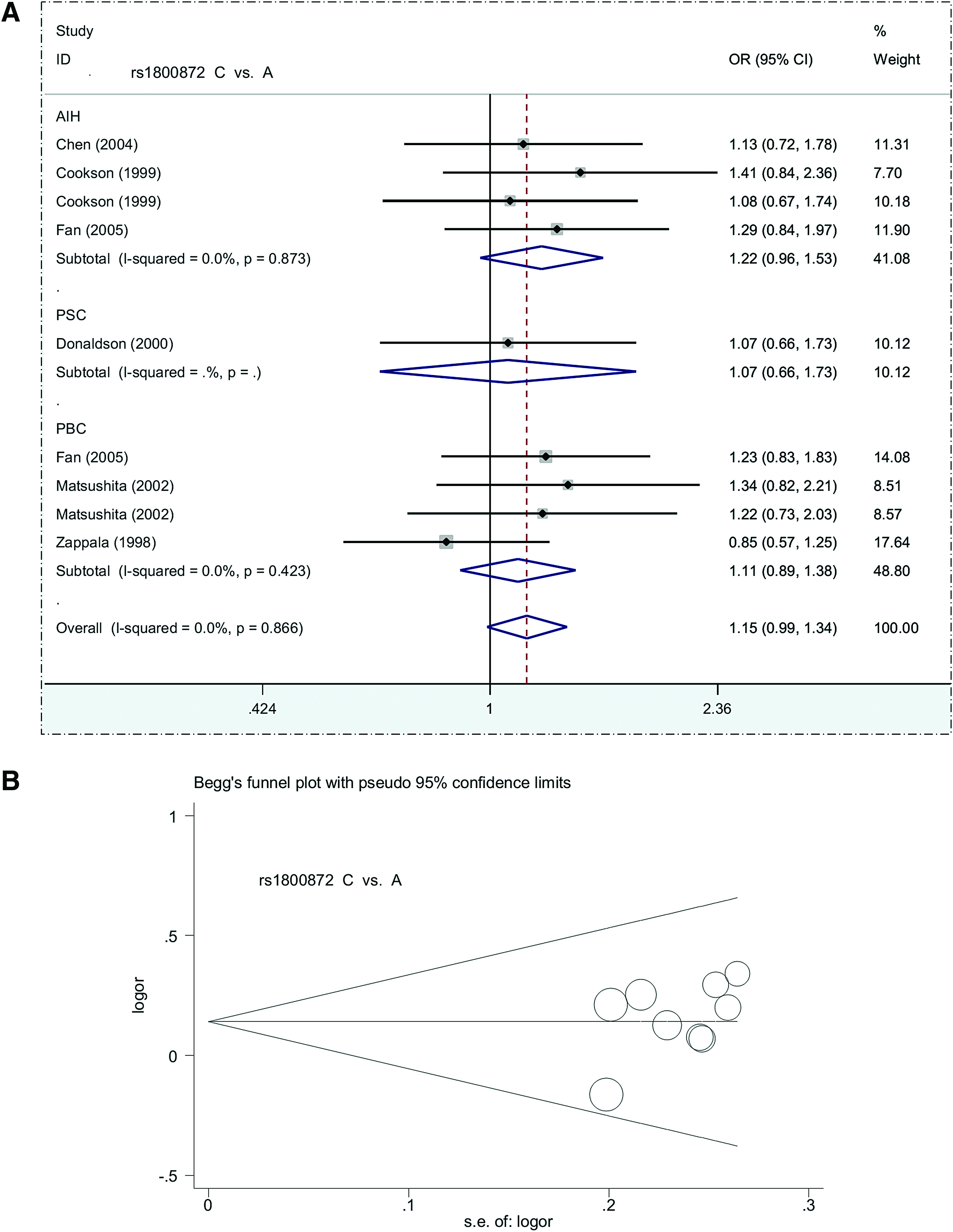
Forest plot and Begg's funnel plot for subgroup analysis of IL10 rs1800872 based on disease type under a C versus A model.
IL10 haplotypes
Finally, we investigated the role of IL10 haplotypes in the risk of autoimmune liver disease. Three promoter haplotypes, “G/C/C” (rs1800896G/rs1800871C/rs1800872C), “A/C/C” (rs1800896A/rs1800871C/rs1800872C), and “A/T/A” (rs1800896A/rs1800871T/rs1800872A), were studied using a fixed-effect model (Supplementary Table S5, all I2 = 0.0%, pheterogeneity >0.1). No genetic relationship between the G/C/C, A/C/C, or A/T/A haplotype and risk of autoimmune liver disease was identified in the overall meta-analysis or in the subgroup analysis of Caucasians (Supplementary Table S5, all passociation >0.05). However, pEgger for “A/C/C” equaled 0.013, suggesting possible potential publication bias. Other pEgger and pBegg values were larger than 0.05 (Supplementary Table S5), suggesting no large publication bias.
Discussion
Genome-wide association studies (GWAS) were previously performed to identify the susceptibility foci for AIH, PBC, and PSC in different populations. For instance, GWAS data from a Chinese population that included 1122 PBC cases and 4036 controls identified risk loci within two cytokine genes, IL21 and IL16 (Qiu et al., 2017). In addition, GWAS data from a European population that included 536 PBS cases and 1536 controls revealed susceptibility foci at rs6441286 and rs574808 within IL12α and rs3790567 in IL12Rβ2 (Hirschfield et al., 2009). Likewise, a potential link between SNPs and the risk of PSC was observed in GWAS data from a Caucasian population that included 4796 cases and 19,955 controls (Ji et al., 2017). Nevertheless, we found no significant association between cytokine gene SNPs and autoimmune liver disease in the results of several GWAS (de Boer et al., 2014; Webb and Hirschfield, 2016; Higuchi et al., 2017). In spite of this, the potential contributions to cytokine gene polymorphisms and relevant immunoregulatory signaling axes to the risk of autoimmune liver disease cannot be excluded. In this study, we examined three polymorphisms within the IL10 gene promoter: rs1800896, rs1800871, and rs1800872.
Although a link between IL-10 and autoimmune responses has been reported (Saxena et al., 2015), results have been inconsistent. For example, the IL10 rs1800896 G/G genotype may be associated with susceptibility to PBS in Italians, but not Japanese (Matsushita et al., 2002). The IL10 rs1800896 A/G SNP was also reported to be a risk factor for PSC in a population from the United Kingdom (Bathgate et al., 2000). The results of the present meta-analysis point to a lack of involvement of IL10 promoter variants (rs1800896, rs1800871, and rs1800872) in the risk of autoimmune liver disease. We also observed a similar negative result for the three IL10 haplotypes (“G/C/C,” “A/C/C,” and “A/T/A”).
The case-control studies included in our analysis were screened using strict inclusion and exclusion criteria. All the included studies contained population-based control data and were of high quality. In addition, no heterogeneity was observed in any of the Mantel-Haenszel statistics, and large publication bias was excluded. Furthermore, the stability of the statistical results was confirmed through sensitivity analysis (data no shown).
Nonetheless, there are several limitations to this study. The first is the small sample size in the included case-control studies. Only 11 case-control studies for rs1800896, 8 for rs1800871, and 9 for rs1800872 were extracted from the 7 eligible articles (Zappala et al., 1998; Cookson et al., 1999; Bathgate et al., 2000; Donaldson et al., 2000; Matsushita et al., 2002; Chen et al., 2004; and Fan et al., 2005). Moreover, the sample size for the subgroup meta-analyses based on disease type (AIH, PBC, PSC) and ethnicity (Caucasian/Asian) were even smaller. Indeed, only one case-control study (Donaldson et al., 2000) was included in the subgroup analysis of the association between IL10 rs1800871 and the risk of autoimmune liver disease. Second, despite the absence of a significant effect in all subgroup meta-analyses, the departure from Hardy-Weinberg equilibrium in some studies weakens the power of the statistical analysis. Third, we only investigated three SNPs of IL10 in our meta-analysis, in that sufficient data were essential for the successful implementation of meta-analysis. The genetic effects of other SNPs (e.g., rs3135932, rs1554286, and rs3024490) or the effect of IL10 in combination with other cytokine genes should be analyzed in the future.
Fourth, AIH, PBC, and PSC all have complex and different etiologies. Additional confounding factors such as age, sex, and patient characteristics should be taken into consideration in future analyses. Fifth, IL10 rs1800896 is reportedly linked to the risk of other autoimmune diseases, including AITD (Jung et al., 2016) and systemic lupus erythematosus (Liu et al., 2013), while IL10 rs1800871 and rs1800872 are reportedly associated with risk of ulcerative colitis (Zou et al., 2014). In that context, and given the essential function of IL10 in autoimmune diseases (Miyagaki et al., 2015), additional studies using both in vitro and in vivo models are needed so as to reach a definite conclusion regarding the negative associations observed here.
In summary, our meta-analysis demonstrated the absence of a relationship between IL10 SNPs (rs1800896, rs1800871, rs1800872) and the risk of AIH, PBC, and PSC.
Footnotes
Acknowledgments
This work was supported in part by grants from the National Natural Science Foundation of China (31370749 and 31670759), the Key Science Foundation of Tianjin Health and Family Planning Commission (16KG151), and National Science and Technology major projects (2017ZX10302202).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.