
Abstract
Aims:
The presence of single nucleotide polymorphisms contributes to genetic diversity, and some are associated with cancer progression. Recent studies concerning the relationship between polymorphisms in miR-146a and the risk of papillary thyroid carcinoma (PTC) have produced conflicting results. Here, a meta-analysis of previous studies was performed to evaluate this relationship.
Materials and Methods:
Electronic databases, including PubMed, China National Knowledge Infrastructure, Cochrane Library, Embase, and Web of Science, were searched for studies concerning miR-146a and PTC published between January 1, 2000 and January 1, 2018. Fixed/random-effects models were used to calculate the pooled odds ratios (ORs) estimated in each study according to the level of heterogeneity.
Results:
Eight studies involving 3993 cases and 9919 controls were assessed. Pooled results showed no association between the miR-146a rs2910164 polymorphism and PTC (OR = 1.001, 95% confidence interval [CI] 0.893-1.121). Subgroup analysis showed that the GG/GC genotype did not significantly increase PTC risk versus CC among Asians (OR = 0.939; 95% CI 0.828-1.066). Similarly, the combination of the GG and GC genotypes did not increase the risk of PTC for Caucasians (OR = 1.571, 95% CI 0.949-2.601).
Conclusions:
The results of our meta-analysis indicated that the miR-146a rs2910164 variant genotype has no effect on susceptibility to PTC.
Introduction
Thyroid cancer is the most prevalent malignancy of endocrine organs worldwide, accounting for ∼90% of all malignancies of the endocrine system. The American Cancer Society estimates the number of new cancer cases in the United States each year, and the estimated new cases of thyroid cancer are ∼54,000 in 2018 (Siegel et al., 2018). The incidence of papillary thyroid carcinoma (PTC) has increased sharply over the past decades (Colonna et al., 2015). Overall, thyroid cancer is now the fifth most common type of cancer affecting women. At a histological level, PTC accounts for ∼80% of thyroid cancer cases (Xing, 2013). Although the genetic predisposition of PTC has been evaluated in pedigree, association, and genome-wide association studies (He et al., 2013, 2018; Tomsic et al., 2016), the detailed mechanism of this disease remains unclear.
MicroRNAs (miRNAs or miRs) are small, noncoding, single-stranded RNA molecules that play an important role in various biological processes, and act as regulators of oncogenes or tumor suppressor genes (Wu et al., 2007). miRNAs function as gene regulators in part by pairing with 3′-untranslated regions in target mRNAs, which usually results in translational suppression (He and Hannon, 2004; Floor et al., 2014). Single nucleotide polymorphisms in mature miRNAs can enhance or decrease the interaction between miRNA and its mRNA target (Ryan et al., 2010).
The role of miRNA in progression of many types of malignant tumors, including thyroid cancer, has been well established (Bartel, 2004; Carthew and Sontheimer, 2009). Studies in mice suggested that miR-146a functioned in oncogenic transformation pathway (Boldin et al., 2011). For example, miR-146a is also involved in the development of cervical cancer (Yue et al., 2011), oral squamous cell carcinoma (Hung et al., 2013), breast and ovarian cancer (Pastrello et al., 2010). Several recent studies evaluated miR-146a polymorphisms in PTC (Jazdzewski et al., 2008; Jones et al., 2012; Marino et al., 2013; Wei et al., 2013; Dong et al., 2015; Zhang et al., 2015), finding that miR-146a could bind to 3′-untranslated regions of the PRKCE gene and decrease the expression of protein kinase C epsilon, which negatively regulates Ras/Raf-1 signal transduction pathways, cellular proliferation, and apoptosis (Zhang et al., 2014). miR-146a can also promote nuclear factor-κB activation through negative feedback on several signal transduction pathways.
However, studies concerning the role of miR-146a in thyroid cancer have produced inconsistent results (Zhang et al., 2014). Therefore, we conducted a meta-analysis of published case-control observational studies to elucidate the relationship between miR-146a rs2910164 polymorphism and risk of PTC. In particular, we focused on studies that examined genetic factors that may be associated with PTC susceptibility to facilitate the establishment of novel and effective methods of detecting and preventing this disease.
Materials and Methods
Literature search strategy
Several electronic databases, including PubMed, China National Knowledge Infrastructure, Cochrane Library, Embase, and Web of Science, were searched using the terms “miR-146a,” “microRNA-146a,” “pre-miR-146a,” and “rs2910164” in combination with “papillary thyroid carcinoma” and “PTC” to identify studies published between January 1, 2000 and January 1, 2018.
Inclusion and exclusion criteria
Studies included in the present meta-analysis fulfilled four criteria: (1) evaluated miR-146a polymorphism and PTC risk in humans, (2) case-controlled studies, (3) identified PTC by histology or pathology, and (4) published in English or Chinese. Studies were excluded if they lacked sufficient data or were not primary research reports (e.g., review articles, letters, conference abstracts, expert opinion, editorials, or case reports). If serial studies or studies involving the same population were identified, only the most complete and/or recent study was included.
Eligible studies and characteristics of selected studies
In the initial literature search, 18 publications were retrieved, and 9 were retained after title and abstract review. Of the remaining nine publications, two were excluded due to lack of controls, and one was a review article (Fig. 1). Thus, a total of six publications conformed to the inclusion criteria. Notably, a publication by Jazdzewski et al. referenced three studies that were conducted in Finland (206 cases, 274 controls), Poland (201 cases, 475 controls), and the United States (201 cases, 152 controls). These studies were added to the pooled analyses of the six publications, which in total described eight studies that involved 3993 cases and 9919 controls. The selected study characteristics are listed in Table 1. Of the eight studies, five involved Caucasian and three involved Asian populations. Almost all of the cases were confirmed by histology, and the control groups were dominated by a healthy population that was matched by age and sex.
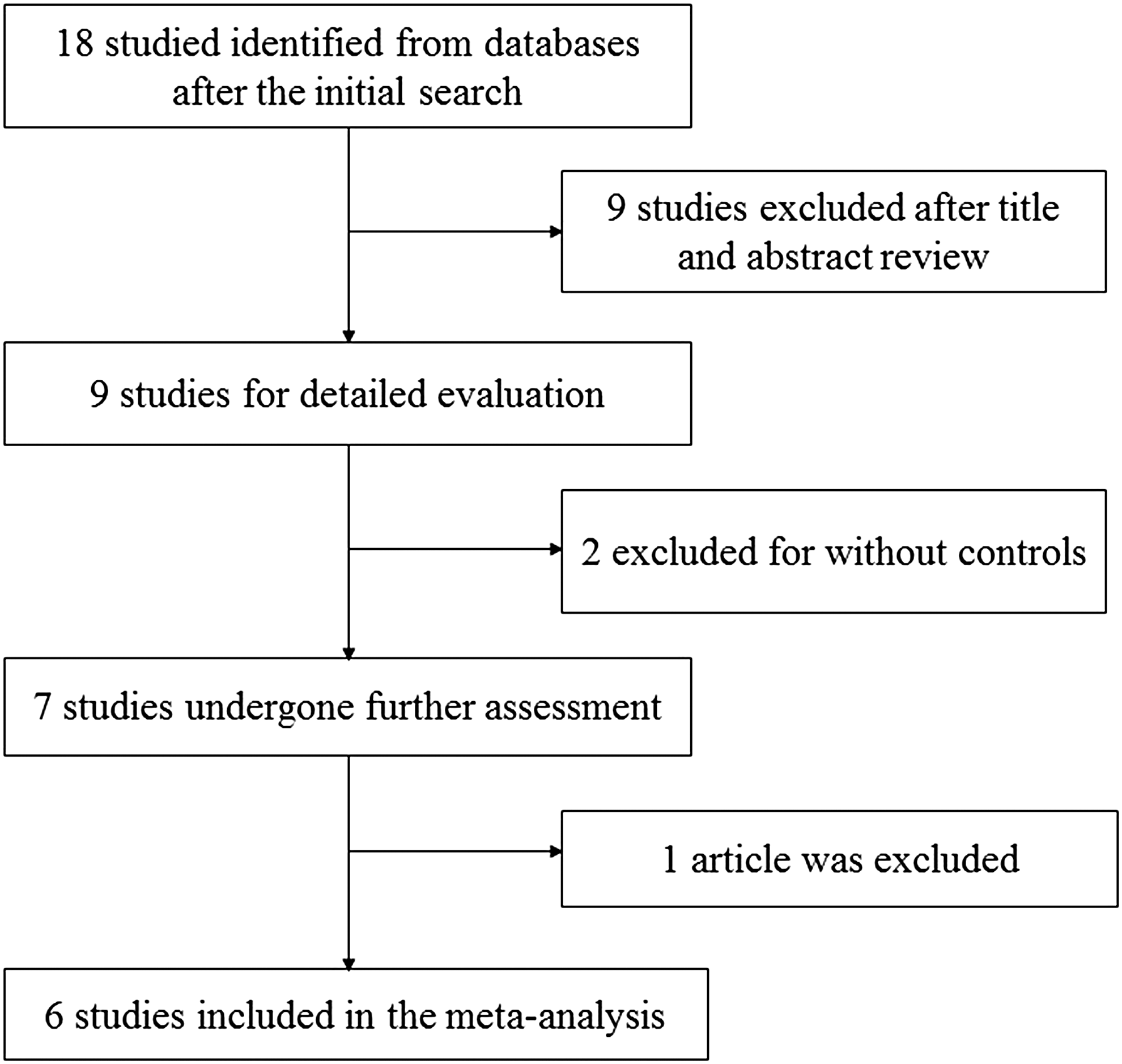
Flow diagram illustrating the process of selecting eligible studies for inclusion in the meta-analysis.
Characteristics from Studies Included in the Meta-Analysis
PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism.
Data extraction
Two authors extracted data from the published studies according to the above inclusion criteria, and disagreements were resolved by discussion. If an agreement could not be reached, a third author was consulted, and the final decision was made based on the majority opinion. The following information was gathered from each study: first author's surname, publication year, country and race of study population, characteristics of matched criteria for controls, total number of cases and controls, genotype distributions in cases and controls, and genotyping methods.
Statistical analysis
Cochrane's Q I2 test was used to conduct the heterogeneity analysis, and a p < 0.10 or I2 > 50% was considered to have statistically relevant heterogeneity. A fixed-effects model was used to calculate the pooled odds ratio (OR) estimates when no significant heterogeneity existed among the studies. A random-effects model was used for all other cases. We also carried out sensitivity analysis to examine the influence of an individual data set on the pooled ORs by deleting each study singly. Begg's funnel plots and Egger's test were used to assess publication bias. Statistical analyses were carried out using Stata 12.0 (Stata Corp., College Station, TX), and a p < 0.05 was considered to be statistically significant. Hardy-Weinberg equilibrium was examined by Pearson's goodness-of-fit χ2 test for each study with p < 0.05 being considered as disequilibrium significance in the control group. Calculation of ORs with 95% confidence intervals (CIs) was used to evaluate the strength of the association between PTC risk and miR-146a rs2910164 polymorphism. ORs with 95% CIs were used to assess the association strength between miR-146a rs2910164 polymorphism and PTC risk. Pooled ORs were obtained by combining each study for the following genotypes: homozygous comparison (GG vs. CC), heterozygous comparison (GC vs. CC), dominant models (GG/GC vs. CC), recessive models (GG vs. CG/CC), and allele comparison (G vs. C). Subgroup analyses were stratified by ethnicity for studies that had sufficient data.
Results
Meta-analysis results
Among the eight included studies involving 3993 cases and 9919 controls, the pooled OR was 1.001 (95% CI 0.893-1.121) for G allele carriers (C/G+G/G) relative to the homozygous wild-type genotype (C/C) (Table 2). Although the p-value for subgroup analysis by ethnicity approached 0.05, no significant association between increased PTC risks and miRNA-146a genetic polymorphism among Caucasian groups with population-based controls for C/G+G/G versus CC (OR = 1.571, 95% CI 0.949-2.601) was found. There was also no significant correlation in the G allele carriers compared with C/C among Asian populations (OR = 0.939, 95% CI 0.828-1.066; Fig. 2A-C).
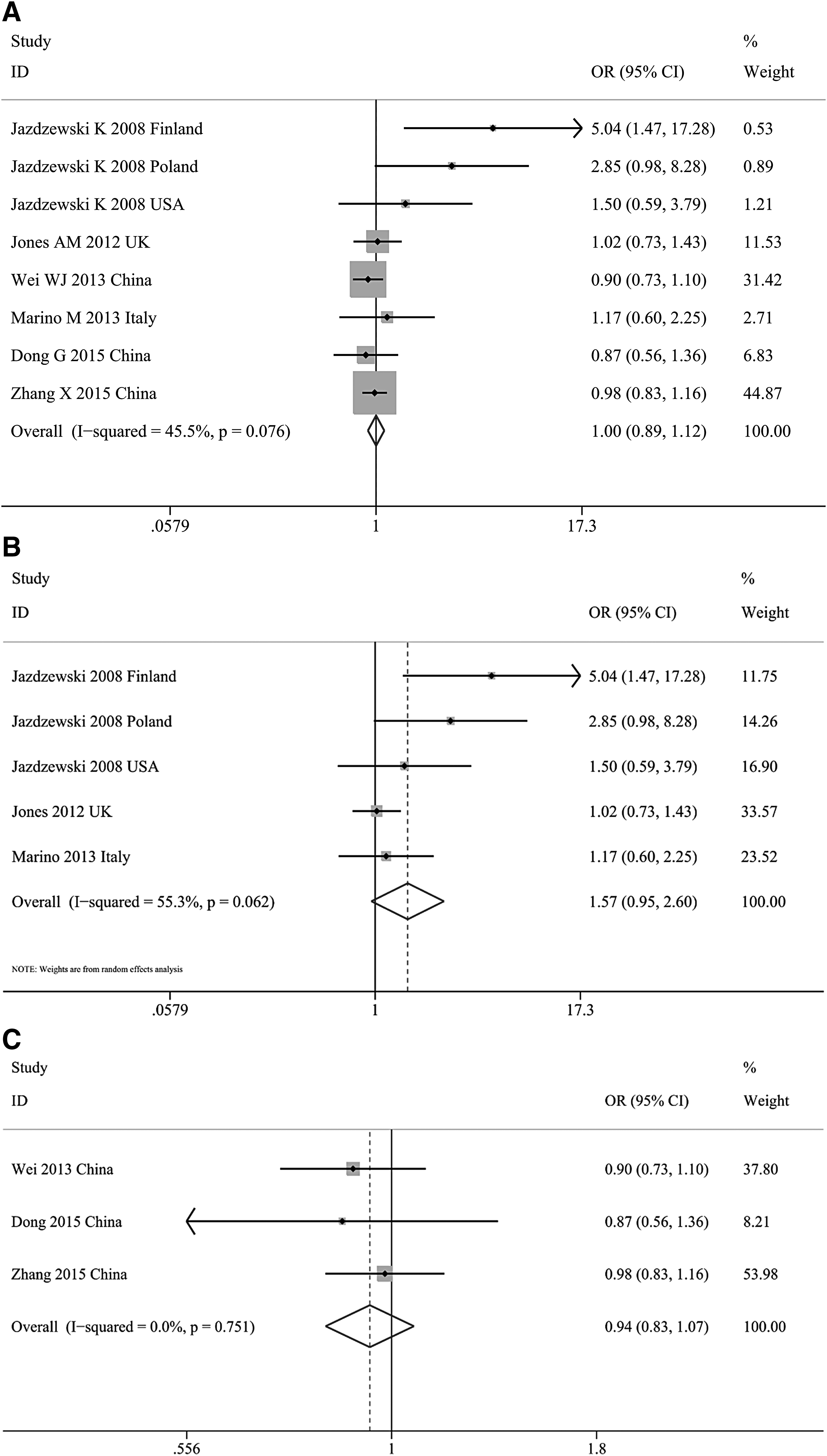
Forest plot of PTC susceptibility associated with miR-146a rs2910164 polymorphism in all studies
Results for Pooled Odds Ratios in the Meta-Analysis
OR, odds ratio; CI, confidence interval; p, test of ORs determined by the Z-test; P-H, test of heterogeneity determined by Cochrane's Q-test.
Sensitivity analyses
We deleted each study in this meta-analysis singly to determine the effect of the individual data set on the pooled ORs. Deletion of each study produced no material change in the corresponding pooled ORs.
Publication bias
The funnel plot showed no obvious asymmetry. Egger's test, an assessment of statistical evidence of funnel plot symmetry, also revealed no publication bias.
Discussion
miR-146a rs2910164 polymorphism occurs in the main area opposite to the mature miR-146a sequence. This G-C polymorphism leads to a mismatch that changes a G:U pair to C:U within the stem of the miR-146a precursor, which in turn affects production of mature miR-146a. This polymorphism is predicted to contribute to the genetic predisposition to PTC and play a role in PTC tumorigenesis. Moreover, changes in miR-146a expression may affect the activity of the nuclear factor-κB signaling pathway in PTC (Leonardi et al., 2012; Zhang et al., 2014). Although researchers have recently focused on the effect of variations in the miR-146a gene in cancer etiologies (Hou et al., 2012; Orsos et al., 2013), the underlying mechanism of miR-146a action in PTC remains unclear. The pathways affected by miRNAs are complex, and changes in miRNA sequences may be manifested at both an organ and cellular level. Environmental changes as well as changes in expression levels can affect miRNA interactions with respective targets, and these interactions can result in miRNA deregulation that drives diverse responses in target genes (Ryan et al., 2010; Leonardi et al., 2012).
Several previous studies have concentrated on the role of miR-146a genetic polymorphisms in PTC risk (Jazdzewski et al., 2008; Jones et al., 2012; Marino et al., 2013; Wei et al., 2013; Dong et al., 2015; Zhang et al., 2015), whereas some studies indicated that miR-146a rs2910164 is related to PTC risk (Jazdzewski K et al., 2008; Dong et al., 2015), others did not detect an association (Jones et al., 2012; Marino et al., 2013; Wei et al., 2013; Zhang et al., 2015). A meta-analysis of these studies that includes a subgroup analysis could provide a different perspective on the relationship between miR-146a rs2910164 polymorphism and PTC risk.
The current meta-analysis summarized the data for ∼4000 PTC cases and ∼10,000 controls to provide a comprehensive evaluation of the relationship between miR-146a rs2910164 polymorphism and risk of PTC. The results showed no evidence to support a genetic association between miRNA-146a rs2910164 and PTC susceptibility. There are likely multiple reasons for this outcome. First, mature miRNAs can interact with a variety of target genes to produce different effects. When a single nucleotide polymorphism occurs in miR-146a, two isoforms can be generated: miR-146a*G, which carries a G allele, and miR-146a*C, which carries a C allele. Individuals who are homozygous for GG and CC can produce two subsets of mature miRNAs, whereas those heterozygous for GC can produce three mature miRNAs (miR-146a, miR-146a*G, and miR-146a*C). Production of these different miRNAs and regulation of distinct target genes by heterozygotes compared with homozygotes might be one reason why some studies detected a predisposition to PTC in heterozygous individuals (Jazdzewski et al., 2009).
Meanwhile, the lack of association may be due to the sample sizes in our meta-analysis. The subgroup analysis of Caucasians yielded a p-value that approached 0.05, which could be indicative of an association between miR-146a single nucleotide polymorphisms and PTC risk, although this possibility awaits additional study. Last, we should consider other aspects, such as different inheritance patterns and gene-environment interaction, regarding the relationship between miR-146a and PTC risk. The miRNA itself could be affected by environmental factors, such as low oxygen (Pineles et al., 2007) or exposure to air pollution (Micolucci et al., 2016). Moreover, a recent study showed that Na+/I− symporter-mediated uptake of radioiodine by thyroid can be modulated by miRNAs (Lakshmanan et al., 2015), whereas Shen et al. found that PTC patients with non-131I and 131I-avid lung metastasis showed differential miRNA expression (Shen et al., 2016). Notably, our analysis did not account for the potential effect of the state of dietary iodine.
This study has several limitations. First, only papers published in English and Chinese were searched. Thus, articles published in other languages may not have been identified, which could have introduced potential selection bias. Second, the included studies all involved Asian and Caucasian populations, and did not consider other ethnicities. Since gene variations can be associated with ethnic background, additional studies are needed to determine whether the association between miRNA polymorphisms and PTC risk is affected by ethnicity.
In summary, the present meta-analysis indicated that the miR-146a rs2910164 polymorphism, particularly the G allele, is not related to the risk of PTC. However, the subgroup analysis of Caucasian study populations reported a p-value that approached 0.05. This suggests that a larger study sample that includes a broader range of ethnic groups with a homogeneous population of PTC patients could reveal associations between miR-146a and PTC that were not detected herein.
Footnotes
Acknowledgment
A grant from the Six Talent Peaks Project in Jiangsu Province, China (No. 2015-WSN-036), supported this study.
Author Disclosure Statement
No competing financial interests exist.