
Abstract
Background and Aims:
The Fc gamma receptor IIa (FcγRIIa), encoded by FCGR2A gene, has been suggested to play a crucial role in immunity by linking immunoglobulin G antibody-mediated responses with cellular effector and regulatory functions. Polymorphisms in FCGR2A have been shown to affect FcγRIIa/antibody interactions and have been potentially implicated in several autoimmune and inflammatory conditions. This study was designed to analyze the association between ulcerative colitis (UC) and FCGR2A polymorphisms in the Chinese population.
Materials and Methods:
A total of 422 patients with UC and 710 unaffected controls were recruited. Five single nucleotide polymorphisms of FCGR2A (rs1801274, rs10800309, rs4657039, rs511278, and rs6696854) were genotyped by SNaPshot. Analyses for linkage disequilibrium (LD) and haplotype studies of FCGR2A were performed for all study subjects.
Results:
The frequency of the minor homozygote (CC) of the rs1801274 SNP of FCGR2A was shown to be significantly lower in patients with UC than in controls (7.1% vs. 12.1%, p = 0.008). Two haplotype blocks, formed by FCGR2A (rs4657039, rs6696854, and rs10800309) and FCGR2A (rs1801274 and rs511278), respectively, were observed in the subsequent LD analysis. The TC haplotype constructed by the major allele of FCGR2A (rs1801274 and rs511278) was more prevalent in UC patients compared with controls (65.2% vs. 60.2%, p = 0.017).
Conclusions:
The minor homozygote (CC) of FCGR2A (rs1801274) may contribute to decrease the susceptibility to UC and the TC haplotype formed by FCGR2A (rs1801274 and rs511278) may increase the risk of UC in the Chinese population.
Introduction
U
Histopathological studies have shown that the intestinal mucosa of patients with active IBD is repleted with various activated polymorphonuclear leukocytes, mononuclear phagocytes, dendritic cells, and immunoglobulin G (IgG) plasma cells. The clinical data also demonstrated that the colonic epithelial cells (CECs) in patients with UC were infiltrated by IgG and activated complement complex, which may be against CECs by binding the CEC surface, as well as mediating antibody-dependent cellular cytotoxicity (ADCC) (Halstensen et al., 1993; Elvington et al., 2015; Trbojević et al., 2015). Moreover, an in vitro study conducted by Uo et al. (2013) revealed that the IgG immune complex (IC) could stimulate abundant production of pro-inflammatory cytokines, for example, tumor necrosis factor (TNF) and interleukin-1β, from lamina propria mononuclear cells in UC patients through cross-linking with the Fc gamma receptors (FcγRs), which is comparable to the stimulation effect of commensal bacteria. Indeed, a previous research on experimental colitis models induced by dextran sulfate sodium salt confirmed that some antibacterial IgG antibodies were themselves pathogenic in the host, because the passive administration of anti-flagellin IgG could promote the acceleration of intestinal inflammation (Kobayashi et al., 2009). Undoubtedly, the previously mentioned observations highlight that massive and local activation of the intestinal mucosal immune, especially the humoral immunity orchestrated by the FcγRs, probably play a crucial role in the pathogenesis of UC.
FcγRs, a group of transmembrane glycoproteins, have evolved as important immunoregulatory molecules, which participate in the clearance of ICs and presentation of the complexed antigen (Lavu et al., 2016). Among them, the FcγRIIa (CD32) is a single-chain low-affinity receptor comprising an extracellular ligand-binding domain and an immunoreceptor tyrosine-based activation motif (ITAM) in the cytoplasmic domain (Gillis et al., 2014). Under physiological conditions, the activating FcγRIIa is constitutively expressed on platelets, neutrophils, macrophages, and dendritic cells, providing a significant link between cellular and humoral immunity by cross-talking IgG antibody-antigen complex (Gillis et al., 2014). Upon binding to the Fc portion of IgG IC, it can activate immune cells by means of phagocytosis, degranulation, antigen presentation, ADCC, and the release of inflammatory cytokines through ITAM. The signaling cascade produced by other pro-inflammatory FcγRs, such as the lipid-anchored neutrophil specific FcγRIIIb and FcγRI expressed on mature macrophages, has also been demonstrated to rely on FcγRIIa (Hogarth, 2002). Consequently, it has been generally accepted that FcγRIIa is a core element for cross-linking the innate and adaptive immune responses, as well as the initiation of inflammatory cascades.
The human FCGR2A gene, containing at least 800 single nucleotide polymorphisms (SNPs), is mapped on chromosome 1q23.3 (Velissari et al., 2015). Of these SNPs, FCGR2A (rs1801274, also assigned as His167Arg or His131Arg) is well described in several autoimmune diseases, including systemic lupus erythematosus (SLE) (Tsang-A-Sjoe et al., 2016), rheumatoid arthritis (RA) (Bek et al., 2017), and immune thrombocytopenia (Pavkovic et al., 2017). This polymorphism may result in a nonconservative amino acid substitution [Histidine (His) to Arginine (Arg)] at position 131 within the second Ig-like domain of the receptor (Lee et al., 2016). This substitution impacts the affinity of FcγRIIa for IgG, with the Arg isoform having a lower affinity than the His isoform (Vogelpoel et al., 2015). Moreover, Lee et al. reported that FCGR2A (rs1801274) polymorphism was associated with better responsiveness to anti-TNF-α therapy in patients with spondyloarthropathy, psoriasis, and CD, particularly in those with follow-up period of >6 months (Lee et al., 2016). Besides, the existing data suggest that the rs1801274, rs10800309, rs4657039, rs511278, and rs6696854 are common SNPs of FCGR2A in Chinese population (https://www.ncbi.nlm.nih.gov/variation/tools/1000genomes). Therefore, we selected the five common SNPs described previously as the research targets to ascertain whether FCGR2A polymorphisms might affect UC susceptibility in Chinese population.
Materials and Methods
Subjects of study
From August 2010 to September 2017, a total of 422 consecutive UC patients without a history of autoimmune diseases and tumors were admitted to The Second and The First Affiliated Hospitals of Wenzhou Medical University, as well as The Central Hospital of Wenzhou city (Wenzhou, China). Diagnosis of UC was made on clinical, radiological, endoscopic, and histopathologic findings according to Lennard-Jones Criteria. Lesion location of UC was defined by Montreal classification, and severity of UC was evaluated by Truelove and Witts criteria (Velikova et al., 2017). The controls consisted of 710 age- and sex-matched healthy blood donors who were recruited at the Health Examination Center of the Second Affiliated Hospital of Wenzhou Medical University (Wenzhou, China), after excluding autoimmune diseases, tumors, and IBD family history. All the participants were Chinese Han population, and their previous three generations were all from Zhejiang province, southeast part of China. The demographic data belonging to all subjects of study are presented in Table 1. All the patients and controls routinely gave their informed consent, and the present study was approved by the medical ethics committee of the three hospitals previously mentioned.
5-ASA, 5-aminosalicylic acid; SASP, sulfasalazine; SD, standard deviation; UC, ulcerative colitis.
Genomic DNA extraction and genotype analysis
Approximately 2 mL of peripheral venous blood was gained from each subject into the Ethylene Diamine Tetraacetic Acid tube and stored at −20°C. DNeasy Blood and Tissue Kit (Qiagen Co., Hilden, Germany) was applied to extract the genomic DNA from the peripheral blood in accordance with the manufacturer's instructions. The genomic DNA was diluted to 10 ng/μL and preserved at 4°C for the subsequent identification of gene polymorphisms.
Genotypes of FCGR2A were determined by ABI Prism SNaPshot Multiplex Kit (Applied Biosystems, Foster City, CA) (Xia et al., 2015). The details of primers used for the polymerase chain reaction (PCR) and single base extension are listed in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/gtmb). The multiplex PCR was performed in a 10 μL volume, containing 1 μL 1× PCR buffer, deoxynucleotides (dNTPs) (final concentration: 0.2 mmol/L), Mg2+ (final concentration: 1.5 mmol/L), 1 μL DNA (10 ng/μL), 1 U HotStarTaq DNA polymerase (Qiagen Co.), and 0.1 μmol/L of each amplification primer. Cycling consisted of an initial 4 min for the predenaturation step at 94°C, 10 cycles using the following conditions: denaturation at 94°C for 30 s, primer annealing at 66°C for 50 s, cutting at 0.5°C per cycle, primer extension at 72°C for 1 min, and eventually another 25 cycles with conditions which are presented as follows: denaturation at 94°C for 30 s, primer annealing at 61°C for 50 s, primer extension at 72°C for 1 min, and final extension at 72°C for 2 min. The 7.5 μL of PCR products were purified to remove excessive primers and un-incorporation dNTPs with 1 U of Exonuclease I (Epicentre, Madison, WI) and 2.5 U of shrimp alkaline phosphates (Promega Co., Madison, WI) at 37°C for 60 min and enzyme inactivation at 75°C for 15 min. The PCR products were examined by electrophoresis in a 3% agarose gel. The SNaPshot multiplex sequencing reaction was performed in a 10 μL reaction volume with 5 μL of SNaPshot Multiplex Mix (Applied Biosystems), 2 μL of purified PCR products, and 0.2 μL of each specific primer (final concentration of each specific primer: 0.1 μmol/L). Thermocycling was performed at 96°C for 1 min, followed by 28 cycles at 96°C for 10 s, at 50°C for 5 s, and at 60°C for 30 s, with a final extension at 60°C for 1 min. The 10 μL of extension products were purified with 1 U of shrimp alkaline phosphates (Promega Co.) at 37°C for 60 min and enzyme inactivation at 75°C for 15 min. The 1 μL of purified extension products were mixed with 9 μL of Hi-Di™ Formamide and 0.5 μL of GeneScan-120 Liz Size Standard (Applied Biosystems). Data were generated by the platform of ABI 3130 Genetic Analyzer and analyzed by GeneMapper 4.0 (Applied Biosystems).
For quality control, 40 cases of the samples were regenotyped under the same conditions described earlier and further confirmed by direct sequencing. Genotypes determined by direct sequencing showed 100% concordance with those examined by SNaPshot Multiplex Kit.
Statistical analyses
The deviation from Hardy-Weinberg equilibrium for each SNP of FCGR2A was calculated by Chi-square test both in UC patients and in the controls. The Chi-square test was used to compare the clinical categorical variables (e.g., sex) between UC patients and the controls, and average age was evaluated by Student's t-test. After the adjustment of age and sex, unconditional logistic regression analysis was applied for investigation of any significant associations between FCGR2A polymorphisms and the overall risk of UC, as well as clinicopathological characteristics of UC patients. Odds radios (ORs) and 95% confidence intervals (CIs) were applied for logistic regression analysis. The linkage disequilibrium (LD) and haplotype frequencies were estimated using the Haploview 4.2 software, which is developed and maintained by Dr. Mark Daly's Lab (Boston, MA). Haplotype blocks were identified using the solid spine of LD method in Haploview 4.2 software (Barrett et al., 2005). All statistical data were input into SPSS 17.0 software (Chicago, IL). Bonferroni correction for multiple comparisons was used to adjust the statistical threshold (0.05/5 = 0.01). Other p-values <0.05 were considered significant.
Results
Comparison of FCGR2A polymorphisms between UC patients and the controls
The alleles and genotypes of FCGR2A (rs1801274, rs10800309, rs4657039, rs511278, and rs6696854) are summarized in Table 2. The genotypic distributions of the five SNPs in FCGR2A conformed to the Hardy-Weinberg equilibrium rule both in UC patients and in the controls (p > 0.05). The minor homozygote (CC) of FCGR2A (rs1801274) was shown to be less frequent in UC patients compared with the controls (7.1% vs. 12.1%, OR = 0.555, 95% CI: 0.360-0.857, p = 0.008). After Bonferroni correction, the previously mentioned result was found to be statistically significant (p < 0.05/5 = 0.01). However, the allele and genotype frequencies of the other four SNPs were not significantly different between UC patients and the controls (Table 2). As described in Tables 3 and 4, after the adjustment of age and gender, there were no significant associations of the five SNPs with the clinicopathological characteristics of UC patients (p > 0.05).
After Bonferroni correction, the statistical threshold was adjusted to 0.05/5 = 0.01.
p Value of statistical significance is in bold.
Adjusted for age and sex.
CI, confidence interval; NA, not calculated due to the low frequencies; OR, odds ratio.
After Bonferroni correction, the statistical threshold was adjusted to 0.05/5 = 0.01.
Adjusted for age and sex.
After Bonferroni correction, the statistical threshold was adjusted to 0.05/5 = 0.01.
Adjusted for age and sex.
Analyses of LD and haplotype for FCGR2A
Figure 1 illustrates pairwise D′ values and predicted haplotype blocks for the five SNPs of FCGR2A. Two blocks were identified using the solid spine of LD method. Block 1 was constructed by FCGR2A (rs4657039, rs6696854, and rs10800309), which were in strong LD with each other [rs4657039/rs6696854 (D′ = 0.970, r2 = 0.752); rs4657039/rs10800309 (D′ = 0.917, r2 = 0.788); and rs6696854/rs10800309 (D′ = 0.993, r2 = 0.841)]. Block 2 consisted of the two SNPs (rs1801274 and rs511278) which were also in a strong LD and formed three haplotypes [rs1801274/rs511278 (D′ = 0.907, r2 = 0.023)]. The frequencies of haplotypes in blocks 1 and 2 are presented in Table 5. Afterward, it was attempted to carry out the haplotype analysis in all subjects of study and evaluate which haplotype was linked to the susceptibility to UC. To minimize potentially false positive associations observed in the haplotype analysis, the haplotypes whose frequencies were simultaneously <3% in UC patients and those in the controls were not calculated. In block 1, no significant association was observed between each haplotype and UC in the present study (p > 0.05). In block 2, however, the frequency of TC haplotype was significantly higher in UC patients than in the controls (65.2% vs. 60.2%, OR = 1.240, 95% CI: 1.039-1.480, p = 0.017).
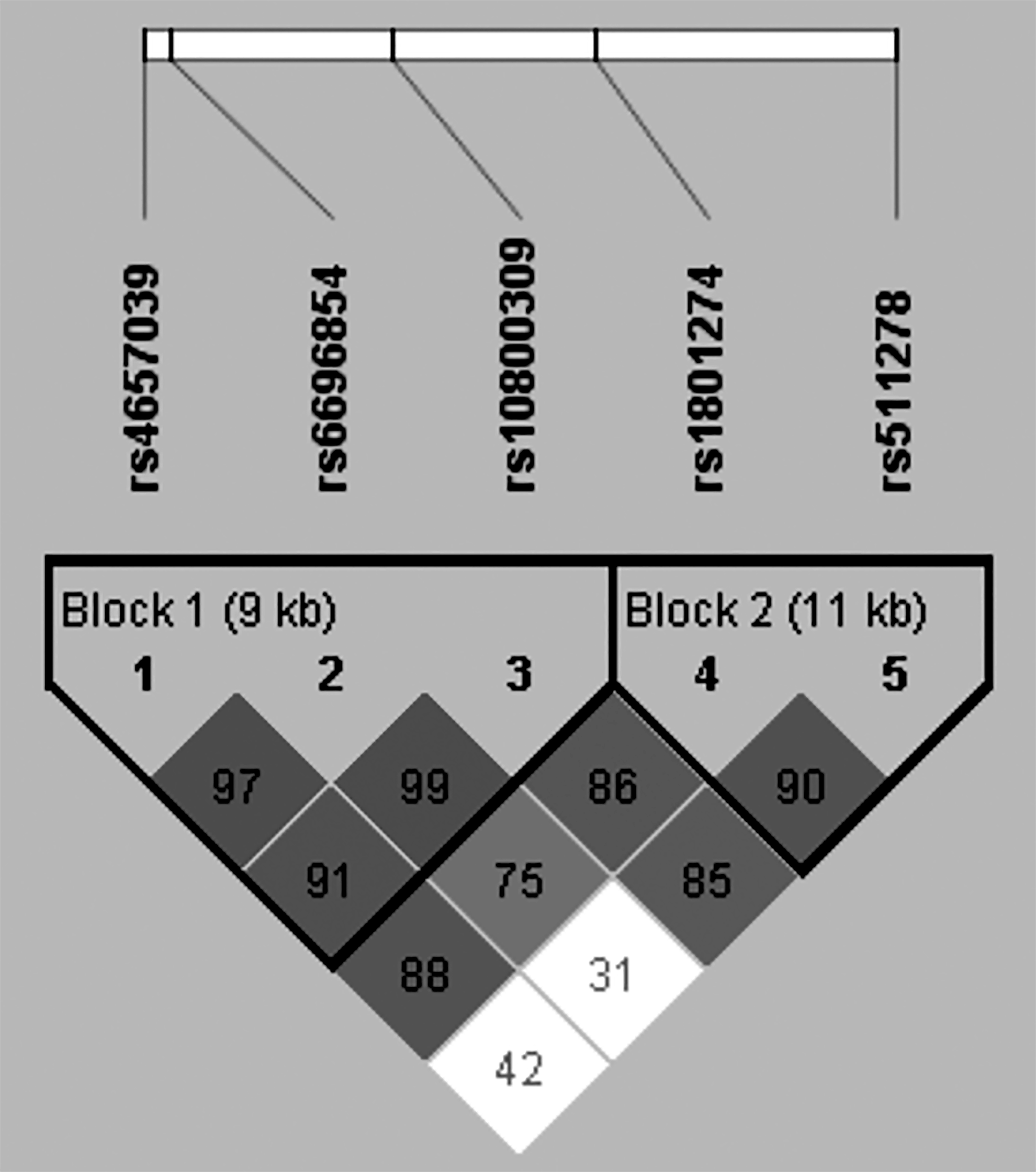
The LD patterns of FCGR2A gene among Han population from southeast of China. The number in a square represents D′ value for each of the two SNPs. Shaded square indicates that there is a strong LD between the two SNPs. Blocks were identified using the solid spine of LD method by Haploview 4.2 software. LD, linkage disequilibrium; SNPs, single nucleotide polymorphisms.
The frequencies simultaneously <3% in both UC patients and the controls were not taken into account.
p = 0.017, OR = 1.240, 95% CI = 1.039-1.480.
Discussion
FCGR2A (rs1801274), which was positioned at the exon 4, has been demonstrated to affect the binding ability of the IgG-ICs to FcγRIIa. Previous studies have implicated such a SNP in the predisposition of many diseases, including giant cell arteritis (Morgan et al., 2006), SLE (Li et al., 2014), sarcoidosis (Typiak et al., 2016), and RA (Dávila-Fajardo et al., 2015). The three SNPs of rs10800309, rs4657039, and rs6696854 were located in the fifth upstream region of FCGR2A, while another polymorphic locus of rs511278 exists in the 3rd downstream. Kim et al. (2009) reported that the genetic polymorphism of FCGR2A (rs6696854) was significantly related with the risk of ischemic stroke in the Korean population. To explore whether this SNP might regulate the transcription of FCGR2A, they also investigated the transcription factor-binding sites using the AliBaba 2.1 (http://www.gene-regulation.com/pub/programs/alibaba2). As a result, they found that the T-containing sequence at this polymorphic locus could bind to the serum response factor, a transcription factor that drives smooth muscle cell-specific gene expression (Kim et al., 2009). In addition, a multiethnic cohort study conducted by Hosgood et al. (2011) revealed that FCGR2A (rs10800309) polymorphism engendered a lower risk of follicular lymphoma.
The present study primarily found that the minor homozygote (CC) of FCGR2A (rs1801274) might contribute to reducing the risk of UC in this cohort of Chinese patients. This finding to some extent was basically consistent with the existing studies from the Japanese (Asano et al., 2009), Caucasian (Weersma et al., 2010; Jostins et al., 2012), and Korean (Yang et al., 2011) populations. Notably, Asano et al. (2009) conducted a genome wide association study in the Japanese population, which highlighted that the minor C allele of FCGR2A (rs1801274) was the most significant nonhuman leukocyte antigen (non-HLA) polymorphism resulting in a decreased risk of UC. Theoretically, FCGR2A (rs1801274) polymorphism can cause the substitution of a His (H131) to an Arg (R131) at amino acid position 131, occurring at the IgG-binding site of FcγRIIa. In contrast with the FcγIIa-H131 isoform, the FcγRIIa-R131 binds to IgG-ICs less efficiently, which ultimately may reduce the levels of cell activation and the induction of cytokines, including TNF-α, matrix metalloproteinase-1, and superoxide (Vogelpoel et al., 2015). In addition, Sanders et al. (1995) showed that neutrophils with the FcγRIIa-H131 isoform efficiently bound to IgG with a threefold higher phagocytosis rate and contained a sevenfold bactericidal activity higher than those with the FcγRIIa-R131. In the current study, we conjectured that the FcγRIIa-R131 determined by FCGR2A (rs1801274) polymorphism might attenuate the risk of UC due, in part, to the downregulation of inflammatory response and inflammation in UC patients.
Although the mentioned individual SNP analysis showed that the allele and genotype distribution of rs511278 did not statistically differ between UC patients and the controls, the further haplotype analysis disclosed that the TC haplotype constructed, respectively, by the major allele of FCGR2A (rs1801274 and rs511278) engendered the increased risk of UC. This implies that the TC haplotype might be a specific genetic marker for predicting the predisposition of UC in Chinese population. It is worth mentioning that the association of the TC haplotype might be partly explained by the T allele of rs1801274 orchestrating higher levels of inflammatory cascades. However, it is still possible that undefined genetic polymorphisms in LD with the TC haplotype existing in other regions of FCGR2A or the adjacent genes (e.g., FCGR3A and FCGR2C) might participate in the occurrence of UC. Morgan et al. (2006) detected a strong LD between FCGR2A (rs1801274) and FCGR3A (158F/V) and found that the TC haplotype formed by FCGR2A-FCGR3A (131R-158F) was significantly associated with the enhanced susceptibility to giant cell arteritis in a cohort of Caucasian patients. Accordingly, exhaustive resequencing is required to find or rule out the possibility of any as-yet-unidentified causal SNP in LD with the TC haplotype.
In conclusion, our findings implied that the minor homozygote (CC) of FCGR2A (rs1801274) may play a protective role in UC patients. Moreover, carriers with the TC haplotype formed by FCGR2A (rs1801274 and rs511278) might be more susceptible to UC in Han people at southeast of China. However, the borderline significance of these associations necessitates further verification such as studies based on a larger sample size. In addition, it was not possible to elucidate the cellular and molecular mechanisms by which FCGR2A polymorphisms might affect the risk of UC. Thus, further studies are required to address the complex interactions among FCGR2A genotypes, IgG levels, and other confounding factors; for example, gut flora, to understand the genetic impacts on the pathogenesis of UC.
Footnotes
Acknowledgments
The authors appreciate the First Affiliated Hospitals of Wenzhou Medical University and the Wenzhou Central Hospital for supplying the blood samples of UC patients. The authors also acknowledge the constructive input provided by the Ministry of Education Key Laboratory of Contemporary Anthropology, School of Life Sciences, Fudan University during the experiments. This study was supported by the Zhejiang Provincial natural science foundation (Grant Nos. LY14H030012, LY15H030018, LY16H160055, and LY17H030011) and the Wenzhou Municipal Science and Technology Bureau (Grant No. Y20160102).
Author Disclosure Statement
The authors declare that there is no conflict of interest regarding the publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.