
Abstract
Aim:
The early gene factor-2 (E2F), a family of transcription factors, is involved in cell cycle regulation. Deregulated expression of most of the members of the E2F family is associated with various human cancers. In this study, we investigated the association between the E2F1 genetic variants rs3213173 (C/T) (Val276Met) and rs3213176 (G/A) (Gly393Ser) with the risk of lung cancer (LC) and head and neck cancer (HNC) in 190 patients and 230 control samples.
Materials and Methods:
We used polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) and mutagenic primer-based PCR-RFLP methods to genotype all target polymorphisms.
Results:
The rs3213173 (C/T) polymorphism was associated with LC risk in the homozygous model (odds ratio [OR] = 2.954, 95% confidence interval [CI] 1.366-6.386; p = 0.004) as well as in heterozygous model (OR = 2.314; 95% CI = 1.369-3.912; p = 0.001). A significant association was also observed for the rs3213176 (G/A) polymorphism with LC risk in homozygous model, GG versus AA (OR = 2.750; 95% CI = 1.236-6.118; p = 0.01); in heterozygous model, GG versus GA (OR = 2.111; 95% CI = 1.256-3.549; p = 0.004); and in combined mutant GG versus GA+AA (OR = 2.214; 95% CI = 1.343-3.650; p = 0.001). The rs3213176 (G/A) marker was also associated with HNC risk.
Conclusions:
Our findings reveal that the rs3213173 (C/T) and rs3213176 (G/A) polymorphisms of the E2F1 gene are genetic risk factors for susceptibility to LC and HNC in the North Indian Population.
Introduction
L
The E2F transcription factors are present downstream to the retinoblastoma (RB) protein in RB/E2F pathway and determine cell fate and cancer development (Polager and Ginsberg, 2009). Both RB and E2F are crucial in proliferation/antiproliferative processes of the cell (Polager and Ginsberg, 2008) and are involved in chromatin assembly/condensation, chromosome segregation, DNA damage response, mitotic spindle checkpoint, apoptosis induction, and development (Ishida et al., 2001; Müller et al., 2001; Ren et al., 2002). Many human cancers have altered expression and activity of various E2F family members (Whibley et al., 2009). E2F1, the best-characterized member, regulates the key genes involved in the DNA replication and G1/S transition to promote the cell cycle (Kowalik et al., 1995; Dyson, 1998; Helin, 1998; DeGregori, 2002). E2F1 is an important player in carcinogenesis as it acts as both an oncogene and a tumor suppressor gene (Polager et al., 2008). Deregulated expression of E2F1induces proliferation and p53-dependent apoptosis in transgenic mice (Pierce et al., 1998a, 1998b). Mice lacking E2F1 have increased susceptibility to develop tumors (Polager and Ginsberg, 2008, 2009). Moreover, E2F1transgenic mice with keratin five promoter develop spontaneous neoplasm in a variety of K5-expressing tissues (Pierce et al., 1999). Contrary to this, E2F1 activation leads to apoptosis by both p53-dependent (Kowalik et al., 1995; Hsieh et al., 2002) and p53-independent pathways (Nahle et al., 2002). In addition, etoposide-treated cells lead to checkpoint kinase-2-dependent phosphorylation and activation of E2F1, thus promoting apoptosis (Polager and Ginsberg, 2009). It is well known that hereditary variants of the coding, promoter, and miRNA binding sites can alter gene functions (Gupta et al., 2005; Lu et al., 2012; Randhawa et al., 2015; Agarwal et al., 2016; Vij et al., 2016, 2017). The roles of nonsynonymous SNPs (nsSNPs) of E2F1gene in the susceptibility of LC and HNC have not been described so far. We hypothesized that nsSNPs in E2F1 might alter their function and transform it to an oncogene, which may predispose individuals to risk of LC and HNC. Therefore, we analyzed the association of its variants rs3213173 (C/T) and rs3213176 (G/A) with LC and HNC risk in patients from North Indian population.
Materials and Methods
Selection of nsSNPs
In this study, we selected two nsSNPs of E2F1, which were predicted as disease risk SNPs of E2F1 by National Institute of Environmental Health Sciences (NIEHS) SNP Program.
Population studied
This study included 230 controls and 190 (100 LC and 90 HNC) patients with an assessed cancer stage and histologically confirmed tumor type. The samples were collected from July 2013 through November 2015 from patients attending Department of Radiotherapy and Oncology, Indira Gandhi Medical College, Shimla. Information regarding demographic variables is given in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/gtmb). The Institutional Ethical Committee of the Jaypee University of Information Technology, Solan, Himachal Pradesh, India, approved this study.
Genotyping
DNA was isolated from peripheral blood following inorganic method (Miller et al., 1988). We used polymerase chain reaction-restriction fragment length polymorphism for genotyping these variants and optimized conditions are given in Supplementary Data. The representative images of genotyping are given in Figure 1A, B. The details of genotyping are presented in Supplementary Data.
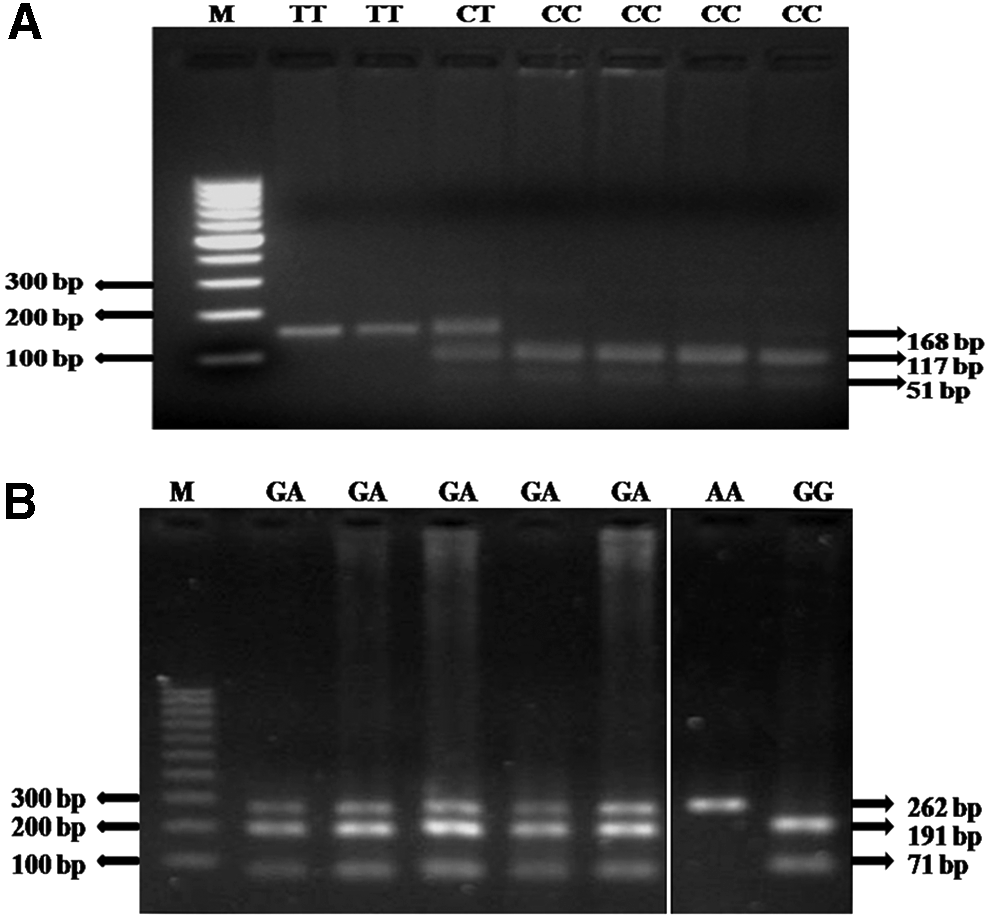
Representative agarose gel images of rs3213173 (C/T) and rs3213176 (G/A) genotyping.
Statistical analysis
Hardy-Weinberg equilibrium was tested for both nsSNPs in patients as well as in the control group. The association between E2F1 polymorphisms and genetic susceptibility to LC and HNC was examined by multiple logistic regression analyses, and odds ratio (OR) and 95% confidence intervals (CIs) were calculated. p-value of <0.05 was considered significant.
Results
In silico analysis of Val276Met and Gly393Ser polymorphisms
In this study, we have used different computational tools to check the effect of amino acid substitutions at the positions 276 and 393 of E2F1 protein. This analysis revealed that Val276Met showed the damaging effect on E2F1 by SNAP, SNPs&GO, and Fathmm, while in silico analysis Gly393Ser polymorphism of E2F1 showed a damaging effect in SNAP tool. The results of the computational analysis are shown in Supplementary Table S2.
Allelic distribution of rs3213173 (C/T) and rs3213176 (G/A) among control and patient groups
The frequencies of the mutant allele (T) of polymorphism rs3213173 (C/T) were 38.6% and 26.11% in LC and HNC patients and 36.4% in controls. rs3213173 (C/T) nsSNP presents a decreased risk of HNC in the allelic model. The rs3213176 (G/A) was associated with higher risk of LC and HNC in the allelic model (Table 1). The frequencies of the mutant allele (A) were 42% and 57.8% in LC and HNC and 29% in controls and patients, respectively.
Bold values represent statistically significant odds ratio and p-values. p-values <0.05 were considered statistically significant.
CI, confidence interval; HNC, head and neck cancer; LC, lung cancer; OR, odds ratio.
Genotypic analysis of rs3213173 (C/T) and rs3213176 (G/A) polymorphisms
Logistic regression analysis of various genetic models revealed a significant association of LC with homozygous, heterozygous, and dominant models for rs3213173 (C/T) polymorphism, while it does not predispose individuals of this region to the risk of HNC (Table 1). We observed a significant association of rs3213176 (G/A) with LC and HNC risk in all the studied genetic models (Table 1).
Association of genotypes with clinical parameters
To elucidate the influence of these polymorphisms on the clinicopathological status of LC and HNC, we analyzed parameters such as tumor node and metastasis (TNM) clinical staging, tumor size, and lymph node metastasis. These polymorphisms were not associated with the clinicopathological status of LC and HNC (Supplementary Tables S3 and S4).
rs3213176 (G/A) is strongly associated with HNC in smoking patients
As we found a highly statistically significant association of rs3213176 (G/A) with LC and HNC (Table 1), we further analyzed if smoking plays any role in the susceptibility of these cancers. Smokers and nonsmokers were segregated in the patient and control groups and analysis revealed a strong association of HNC in all the genetic models (Table 2).
Bold values represent statistically significant odds ratio and p-values. p-values <0.05 were considered statistically significant.
Discussion
The marked box domain and its adjacent region of E2F1 are responsible for their unique ability to strongly induce apoptosis (Hallstrom and Nevins, 2003). The analyzed nsSNPs of this gene are located in these regions, which interact with Jab1 (apoptosis inducer cofactor factor) (Hallstrom and Nevins, 2006). The selected nsSNPs in this domain could alter its structure and interactions with Jab1, leading to reduced apoptosis, and may play a role in cancer susceptibility. Various studies have reported the role of SNPs in structural and functional changes in the protein and susceptibility of various diseases (Gupta et al., 2005; Randhawa et al., 2015; Agarwal et al., 2016; Vij et al., 2016, 2017). SNPs in the regulatory region of E2F1 and E2F2 were associated with the early onset of squamous cell carcinoma of the HNC in American population (Lu et al., 2012). Moreover, a recent study on Indian population has also shown rs3213172 (C/T), rs3213173 (C/T), and rs3213176 (G/A) as a risk factor for cervical cancer (Singh et al., 2018).
In this study, we observed a significant association of Val276Met and Gly393Ser variants of E2F1 protein with the cancer risk. The allele and genotype frequencies of these two SNPs exhibited significant differences between the case and control groups. This is the first report investigating the role of E2F1 coding region variants in LC and HNC risk. We found a strong association of rs3213176 (G/A) polymorphism with the risk of HNC in the homozygous model (11.39-fold), the heterozygous model (3.469-fold), as well as in the dominant model (4.745-fold). This SNP also increases the risk of LC in the homozygous model (2.750-fold), heterozygous model (2.111-fold), and dominant model (2.214-fold) (Table 1). Furthermore, the risk of HNC (in homozygous, heterozygous, and dominant models) is increased among smokers with rs3213176 (G/A) polymorphism (Table 2).
A study on E2F1 gene polymorphism [rs35301225 (C/A)] in Chinese population revealed its involvement in the occurrence of colorectal cancer by the upregulation of E2F1 gene. Also, this SNP was associated with tumor size, tumor differentiation, and metastasis in colorectal cancer (CRC) patients (Jiang et al., 2017). In our population, we could not find any association with the clinicopathological parameters (TNM clinical staging, tumor size, and lymph node metastasis) (Supplementary Tables S3 and S4).
Conclusion
In this study, we observed a statistically significant association between the rs3213173 (C/T) polymorphism with LC risk and rs3213176 (G/A) polymorphism with LC and HNC risk. These polymorphisms of E2F1 gene might serve as a valuable prognostic biomarker for genetic susceptibility to LC and HNC and hence need to be explored further in replication studies from other populations with larger patient sample size.
Footnotes
Acknowledgments
This work was supported by grants BT/PR6784/GBD/27/466/2012 and SB/FT/LS-440/2012 to H.C. from Department of Biotechnology, Government of India, and Department of Science and Technology, Government of India, respectively. S.S. is thankful to Jaypee University of Information Technology, Solan, Himachal Pradesh, India, for Junior Research Fellowship.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.