
Abstract
Background:
Human amniotic membrane-derived mesenchymal stem cells (hAM-MSCs) are considered a new and favorable source of stem cells for cell replacement-based therapy. Some microRNAs (miRNAs) have been reported to participate in the regulation of immune responses. Our aim was to investigate the effects of miR-21 on the biological characteristics, immunoregulatory properties, and potential mechanisms of hAM-MSCs.
Methods:
hAM-MSCs were isolated from the placental amnion membrane of a newborn. Cell proliferation, cell cycle, apoptosis, and expressions of cell surface markers were measured by CCK-8 and flow cytometric assays in hAM-MSCs. The expression of mesenchymal-specific antigens vimentin and stage-specific embryonic antigen-4 (SSEA-4) were identified by immunofluorescence staining. Tumor necrosis factor alpha (TNF-α), monocyte chemotactic protein-1 (MCP-1), and interleukin-10 (IL-10) expressions in the cocultured supernatant of hAM-MSCs and peripheral blood mononuclear cells (PBMC) were detected via enzyme-linked immunosorbent assays (ELISA).
Results:
Flow cytometric analyses revealed that the positive expression rates of the cell surface markers CD29, CD44, CD73, and CD90 in hAM-MSCs were 97.3%, 96.3%, 97.8%, and 98.2%, respectively, while the rates of CD34 and CD45 expression were only 0.6% and 0.84%, respectively. The immunofluorescent staining results showed that vimentin and SSEA-4 were positive in hAM-MSCs. CCK-8 assays revealed that miR-21 overexpression significantly promoted hAM-MSC proliferation. Cell cycle analyses revealed that the number of hAM-MSCs-miR-21+ cells during the synthesis phase (S phase) was significantly increased. miR-21 overexpression also significantly inhibited apoptosis in hAM-MSCs. The ELISA analyses revealed that miR-21 overexpression enhanced the inhibitory effect of hAM-MSCs on the secretion of TNF-α and MCP-1 as well as the promotive effect on the secretion of IL-10 in PBMC cocultured with miR-21-hAM-MSCs. In addition, miR-21 downregulation reduced the inhibitory effect of hAM-MSCs on the secretion of TNF-α, MCP-1, and the promotive effect on the secretion of IL-10 in PBMC cocultured with anti-miR-21-hAM-MSCs.
Conclusions:
Our data showed that miR-21 promoted hAM-MSCs proliferation, inhibited apoptosis, and was involved in controlling the immunoregulatory capacity of hAM-MSCs.
Introduction
Stem cells are rare and uniquely capable of both reproducing themselves and generating differentiated cell types; as such, they play a pivotal role in the regeneration of damaged tissues, as well as aid in the recovery of functional absences in human diseases (Zeng et al., 2011). Mesenchymal stem cells (MSCs) have been found in multiple tissues, including but not limited to bone marrow, adipose tissue, the umbilical cord, and Wharton's jelly (Crisan et al., 2008). They are a promising cell resource for tissue engineering and cell-based therapeutics, due to their capability of self-renewal and differentiation into specific functional cell types (Jiao et al., 2012). MSCs transplanted into animal models that have damaged brains and spinal cords can survive, migrate, proliferate, and differentiate; they have also been reported to decrease neuronal injuries and improve the recovery of functional deficits (Khoo et al., 2008). Previous studies have shown that amniotic membrane-derived MSCs have low immunogenicity and anti-inflammatory functions making them a potential source of stem cells for the treatment of central nervous system disorders (Chao et al., 2009; Wolbank et al., 2009). Presently, the number of studies evaluating the roles that microRNAs (miRNAs) play on the viability, proliferation, and apoptosis of human amniotic membrane derived-MSCs (hAM-MSCs) are limited.
There has been extensive research focused on multifold target genes of histone modification, miRNAs, DNA methylation, MSCs, and their ability to help with the tissue-specific repair of many different organs. The mechanisms of how they interact and cooperate in a biological context, however, are not yet fully understood. Increasing numbers of miRNAs appear to modulate differentiation in MSCs, potentially providing a tissue-type-specific therapeutic vehicle to combat various diseases (Zhao et al., 2014; Tang et al., 2018). Within the context of immunoregulation, miRNAs have been reported to participate in the regulation of some immune responses. miR-937 in MSCs may significantly enhance the therapeutic effects of MSCs for the treatment of Alzheimer's disease (Liu et al., 2015). A previous study has exhibited that miR-146a plays a vital role in modulating the immunoregulatory potential of bone marrow-derived MSC (BMSCs) (Matysiak et al., 2013). It has been presented by Trohatou et al. (2014) that miR-21 might specifically function by suppressing sex determination region Y-box2 (Sox2) expression in human MSCs and acts as a key molecule for determining MSC proliferation and differentiation. Recently, Shi et al. (2018) have reported that BMSCs-derived exosomal miR-21 could protect C-kit+ cardiac stem cells from oxidative injury by the PTEN/PI3K/Akt axis. Based on these studies, we speculated that miRNA-21 might affect the fate of hAM-MSCs and their functions in the immunoregulation of hAM-MSCs by affecting the release of inflammatory cytokines.
To test this hypothesis, we determined the effects of miRNA-21 on cell proliferation, cell cycle control, and apoptosis of hAM-MSCs. Moreover, we analyzed the expression of miR-21 in hAM-MSCs to assess their immunoregulatory potential.
Materials and Methods
Isolation and culture of hAM-MSCs
hAM-MSCs were isolated via enzyme digestion from the placental amnion membrane of a newborn from a healthy donor mother after a cesarean section. This procedure was carried out at the First Affiliated Hospital of Zhengzhou University. The placenta was rinsed in phosphate-buffered saline (PBS; Sigma Chemical Co., St. Louis, MO) containing 1% penicillin-streptomycin (Nacalai Tesque, Kyoto, Japan). Amniotic membranes were isolated according to previously described methods (Jiao et al., 2012). The amniotic membranes were isolated with sterile forceps and washed thoroughly with sterile PBS. The membranes were then mechanically fragmented, washed with PBS three times and subjected to an enzymatic digestion treatment. The digest was a two-step process: (1) incubation with 0.25% trypsin (Sigma) at 37°C for 1 h to eliminate epithelial amniotic cells, (2) treatment with 0.1% collagenase IV (Gibco Invitrogen, Pais-ley, United Kingdom) for 2 h at 37°C, followed by inactivation with bovine fetal serum. The cell suspension was then filtered and the isolated cells were seeded in a 75-cm2 cell culture flask. The flask contained Dulbecco's modified Eagle's medium/nutrient mixture F-12 (DMEM/F12; Gibco BRL, Life Technologies, Grand Island, NY) medium with 10% fetal bovine serum (FBS; Invitrogen), 20 μg/L basic fibroblast growth factor (Invitrogen, California), and 1% penicillin-streptomycin. The cells were then incubated and grown at 37°C in a 5% CO2 atmosphere. hAM-MSCs in the third cell passage (P3) were used for the further experiments. The study protocol was approved by the Ethics Committee of the First Affiliated Hospital of Zhengzhou University, and written informed consent was obtained from the patient.
Flow cytometry analyses of cell surface antigens, cell cycle, and apoptosis
hAM-MSCs from the P3 were plated at a density of 1 × l04 cells/cm2 and were analyzed by flow cytometry. As described by the previous research methods (Zeng et al., 2011), the cells were cultured for 24 h and then harvested for flow cytometry immediately after trypsinization. The cells were incubated with the following fluorescein-labeled antibodies: CD29-PE, CD44-PE, CD73-PE, CD90-FITC, CD34-PE, and CD45-FITC (San Jose, CA) for 30 min at 4°C. IgG1-PE and IgG1-FITC antibodies were used as negative controls. The samples were placed in 0.3-0.4 mL of PBS buffer with 1% bovine serum albumin (BSA) and 0.05% sodium azide. Then, the cell suspension was fixed with 1% paraformaldehyde and 2% BSA for 20 min at room temperature. The cells were analyzed via a FACSCalibur flow cytometer.
The cells (P3) were cultured for 48 h. The cell cycle analysis was performed using propidium iodide (Sigma-Aldrich) according to previous method (Queiroz et al., 2014). The cell cycle analysis was carried out using the FACSCalibur flow cytometer.
The cells (P3) were cultured for 24 h. Cell apoptosis was detected using the Annexin V-phycoerythrin/7-amino-actinomycin D Apoptosis Detection Kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China) according to the manufacturer's instructions. Cell apoptosis was analyzed using the FACSCalibur flow cytometer.
Immunofluorescence assays
1 × l04 cells (P3) were plated on a 48-well dish. After 24 h, the cells were washed twice with PBS containing 1% penicillin-streptomycin and fixed in 4% paraformaldehyde overnight at 4°C. The cells were then treated with 0.1% Triton × 100 in PBS at 4°C for 15 min. Next, the cells were incubated with 3% H2O2 for 10 min and then washed twice with PBS. Following the wash step, the cells were blocked with 10% goat serum in PBS for 30 min and then incubated with the primary antibodies (vimentin and stage-specific embryonic antigen-4 [SSEA-4]) (Invitrogen) overnight at 4°C. The next day, the cells were washed again with PBS and incubated with the FITC-labeled secondary antibodies (Invitrogen) overnight at 4°C. The cells were then stained with 2-(4-amidinophenyl)-6-indolecarbamidine dihydrochloride and photographed using an immunofluorescence microscope.
Plasmids and lentivirus packaging
The lentiviral vectors that can express a green fluorescent protein, including pCDH-CMV-MCS-EF1-copGFP-T2A-Puro (Lv-Con1) and pLent-hU6-GFP-Puro plasmids (Lv-Con2), were obtained from the GenePharma Company (Shanghai, China). The packaging plasmids were pCMV-Δ8.2 and pCMV-VSV-G (GenePharma). The miR-21 sequence was amplified from the genomic DNA of HEK293-T cells. The anti-miR-21 sequence was synthesized by GenePharma Company.
Before lentivirus packaging, HEK293-T cells were plated in 10 cm cell culture dishes at density of 2 × 106 cells/dish. The lentiviral vector packaging system was prepared with the following steps: a solution of 1 mL was prepared consisting of 14 μg of plasmid pCMV-Δ8.2, 7 μg of pCMV-VSV-G of, 21 μg of expression plasmid (pLent-miR-21 or pLent-anti-miR-21), and 50 μL of Lipofectamine 2000 (Invitrogen) in DMEM medium. The lentiviral vector packaging systems were added to the HEK293-T cells with a confluence of 70-80%, respectively. At 48 h posttransfection, the supernatant was harvested by centrifugation at 1000 rpm for 5 min at 4°C. The ratio of positive cells was measured by real-time quantitative polymerase chain reaction (qRT-PCR). We obtained the lentiviruses and determined that the infection rate was higher when the virus titer was multiplicity of infection (MOI) = 20. Finally, hAM-MSCs were infected with the lentivirus (MOI = 20) to produce stable target cell line.
Real-time quantitative RT-PCR assays
Total RNA was extracted from cultured cells using TRIzol reagents (Invitrogen) according to the manufacturer's instructions. For the miRNA analyses, miRNA was transcribed into first-strand cDNA using the miRcute miRNA First-strand cDNA Synthesis Kit (Tiangen, Beijing, China). The miRNA expression data were normalized to RNU6B (U6) and calculated with the comparative threshold cycle (Ct) (2−ΔΔCt) method.
CCK-8 assays
hAM-MSCs were plated into 96-well plates at a density of 5 × 103 cells/well and cultured for the indicated times (1, 2, 3, 4, or 5 days). Ten microliters of CCK-8 solution (CCK8; Dojindo, Kumamoto, Japan) was added to the wells and then incubation was continued for an hour. The absorbance was then measured at 450 nm using an MRX II microplate reader (Dynex Technologies, Chantilly, VA).
Coculture of hAM-MSCs with peripheral blood mononuclear cells
Peripheral blood mononuclear cells (PBMC) were obtained from heparinized blood using a cell separation tube and separation medium (Ficoll Paque Plus; GE Healthcare, Freiburg, Germany), according to the manufacturer's instructions. The cells were resuspended in RPMI1640 (GIBCO, Grand Island, NY) supplemented with 10% FBS (GIBCO). The sample preparation protocols were approved by the Ethics Committee of The First Affiliated Hospital of Zhengzhou University.
hAM-MSCs were plated into 96-well plates at density of 5 × 103 cells/well and cultured for 24 h at 37°C in an incubator with 5% CO2. The cells were treated with 5 μL of mitomycin C (MMC; 1 μg/μL) (Sigma) for 30 min at 37°C, and then washed twice by PBS. To assess the effects of hAM-MSCs on cytokine secretion of PBMC, phytohemagglutinin (PHA) was used to stimulate the activation of lymphocytes from PBMC as described by the previous studies (Upratya et al., 2010; Poltavtsev et al., 2017; Diedrichs-Mohring et al., 2018). PBMC were seeded at a density of 1 × 105 cells/well on hAM-MSCs and treated with PHA (1 μg/μL).
Enzyme-linked immunosorbent assays
hAM-MSCs and PBMC were cocultured for 24 h. The cell supernatants were collected and then the levels of tumor necrosis factor alpha (TNF-α), monocyte chemotactic protein-1 (MCP-1), and interleukin-10 (IL-10) were measured in the conditioned medium with enzyme-linked immunosorbent assay (ELISA) kits (Invitrogen), according to the manufacturer's instructions.
Statistical analysis
All experiments were performed using at least three biological replicates. Among three or more groups, an one-way analysis of variance was used to assess the differences. p values <0.05 were considered significant.
Results
Growth characterization of hAM-MSCs
The enzyme digestion was performed to isolate hAM-MSCs from the placental amnion membrane of a newborn after a cesarean section. The growth features of hAM-MSCs were observed under an inverted microscope. The majority of cells grew with adherence and a polygonal shape in the primary culture (at passage 0, P0). In addition, when hAM-MSCs were passed up to the third passage (P3), the adherent cells presented with a spindle-shaped like morphology and pseudopodia formation. Moreover, the cells were close together and arranged neatly (Fig. 1).

Morphology and growth of hAM-MSCs. (P0) hAM-MSCs were cultured in the primary passage. (P3) hAM-MSCs were cultured to the third passage and displayed a spindle-shaped-like morphology with pseudopodia formation. The cells were close together and arranged neatly. All the scale bars represent 100 μm. (magnification, × 100). hAM-MSC, human amniotic membrane-derived mesenchymal stem cell.
Identification of hAM-MSCs
hAM-MSCs (P3) were analyzed by flow cytometric analyses and immunofluorescence staining. As shown in Figure 2, the results from the flow cytometric analyses revealed that the positive expression rates of the cell surface markers CD29, CD44, CD73, and CD90 in hAM-MSCs were 97.3%, 96.3%, 97.8%, and 98.2% at P3, respectively. The rates of CD34 and CD45 expression were only 0.6% and 0.84%, respectively. Moreover, immunofluorescence staining results revealed that vimentin and SSEA-4 were positive in hAM-MSCs (Fig. 3). These data showed that hAM-MSCs could be successfully isolated and cultured, while maintaining the characteristics of MSCs, as previously demonstrated by other studies (Magatti et al., 2008).

Expression of the cell surface markers in hAM-MSCs (P3). Cells were incubated with fluorescein-labeled antibodies against: CD29-PE, CD44-PE, CD73-PE, CD90-FITC, CD34-PE, and CD45-FITC for 30 min at 4°C. IgG1-PE and IgG1-FITC antibodies were used as negative controls. The cell surface markers CD29, CD44, CD73, and CD90 in hAM-MSCs were detected by flow cytometry analyses.
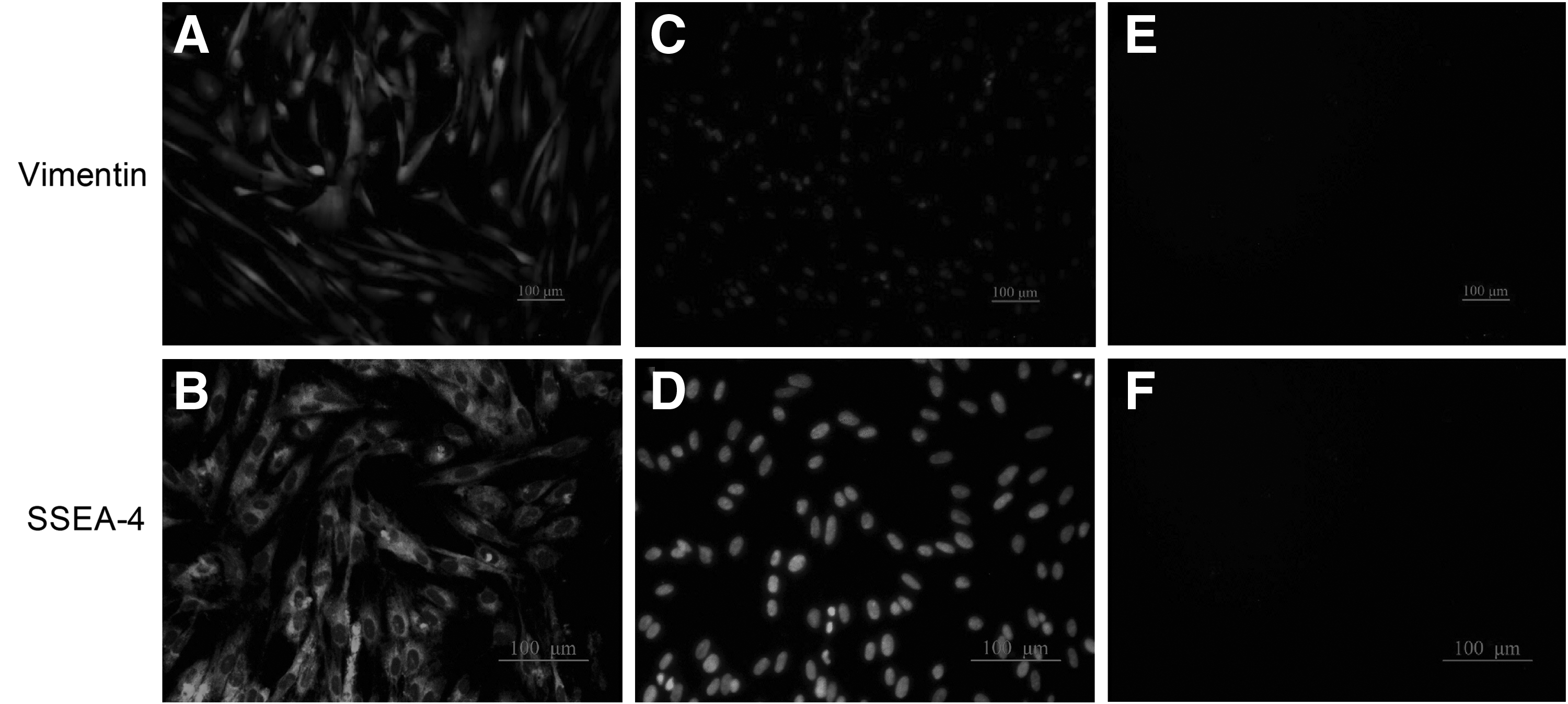
Expression of human antigens in hAM-MSCs (P3). Immunofluorescent staining was used to determine the expression of vimentin and SSEA-4. Representative images showed that vimentin (magnification, × 100) and SSEA-4 (magnification, × 200) were positive in the hAM-MSCs. All the scale bars represent 100 μm.
Upregulation and downregulation of miR-21 in transfected hAM-MSCs
To investigate the effects of miR-21 on the biological function of hAM-MSCs, gene deletion and acquisition experiments were undertaken. hAM-MSCs infected with lentiviruses carrying LV-anti-miR-21 or LV-miR-21, respectively, and the infected hAM-MSCs showed green fluorescence under a fluorescence microscope for the majority (Fig. 4A). qRT-PCR analyses showed that the levels of miR-21 were significantly increased in the hAM-MSCs-miR-21 cells, when compared with hAM-MSCs-LV-Con1. There was a corresponding obvious reduction in the level of miR-21 in hAM-MSCs-anti-miR-21 cells compared with uninfected hAM-MSCs-LV-Con2 at both 2 and 5 days posttransfection (Fig. 4B). We assessed cell surface marker expression after transfection and did not find an obvious alteration for any of the assayed markers (CD29, CD44, CD73, CD90, CD34, and CD45) nor human antigens (vimentin and SSEA-4) (data not shown). These data suggested that we obtained stable hAM-MSCs-miR-21 and hAM-MSCs-anti-miR-21 cell lines.

Upregulation and downregulation of miR-21 in transfected hAM-MSCs.
Upregulation of miR-21 promoted proliferation and suppressed apoptosis of hAM-MSCs
In this study, we explored the potential effects of miR-21 on proliferation and apoptosis of hAM-MSCs via CCK-8 assays and flow cytometry analyses. As shown in Figure 5A, the miR-21 overexpression led to a significant increase in proliferation of hAM-MSCs-miR-21 cells in comparison with control hAM-MSCs. The miR-21 downregulation obviously suppressed cell proliferation. In addition, the results from the flow cytometric analyses revealed that, compared to the hAM-MSCs control group, the number of hAM-MSCs-miR-21 cells at S-phase was significantly increased, and the number of S-phase hAM-MSCs-anti-miR-21 cells was significantly decreased (Fig. 5B). Flow cytometry results showed that, compared to the control group, upregulation of miR-21 led to a marked decrease in apoptosis in hAM-MSCs-miR-21, whereas knockdown of miR-21 significantly increased apoptosis in hAM-MSCs-anti-miR-21 cells (Fig. 5C). These data suggested that upregulation of miR-21 could promote proliferation and suppress apoptosis of hAM-MSCs.

Upregulation of miR-21 promoted proliferation and suppressed apoptosis of hAM-MSCs.
Upregulation of miR-21 promoted immunoregulatory effect of hAM-MSCs on PBMC
Previous studies have shown that hAM-MSCs have low immunogenicity and anti-inflammatory functions (Chao et al., 2009; Wolbank et al., 2009). Nevertheless, the potential mechanisms of the immunoregulatory functions of hAM-MSCs and whether miR-21 is involved remain unclear. Expressions of the inflammatory cytokines, TNF-α, MCP-1, and IL-10 in PBMC cocultured with hAM-MSCs were detected via ELISA. As shown in Figure 6A-C, the results demonstrated that, compared to PBMC cultured alone, hAM-MSCs significantly inhibited the secretion of TNF-α and MCP-1, as well as promoted the secretion of IL-10 in PBMC cocultured with hAM-MSCs. This suggested that hAM-MSCs have immunosuppressive properties. Compared with PBMC cocultured with hAM-MSCs, miR-21 overexpression enhanced the inhibitory effect of hAM-MSCs on the secretion of TNF-α and MCP-1 and the promotive effect on the secretion of IL-10 in PBMC cocultured with miR-21-hAM-MSCs. miR-21 downregulation by coculture of anti-miR-21-hAM-MSCs with PBMC reduced the inhibitory effect of hAM-MSCs on the secretion of TNF-α, MCP-1, and the promotive effect on the secretion of IL-10.
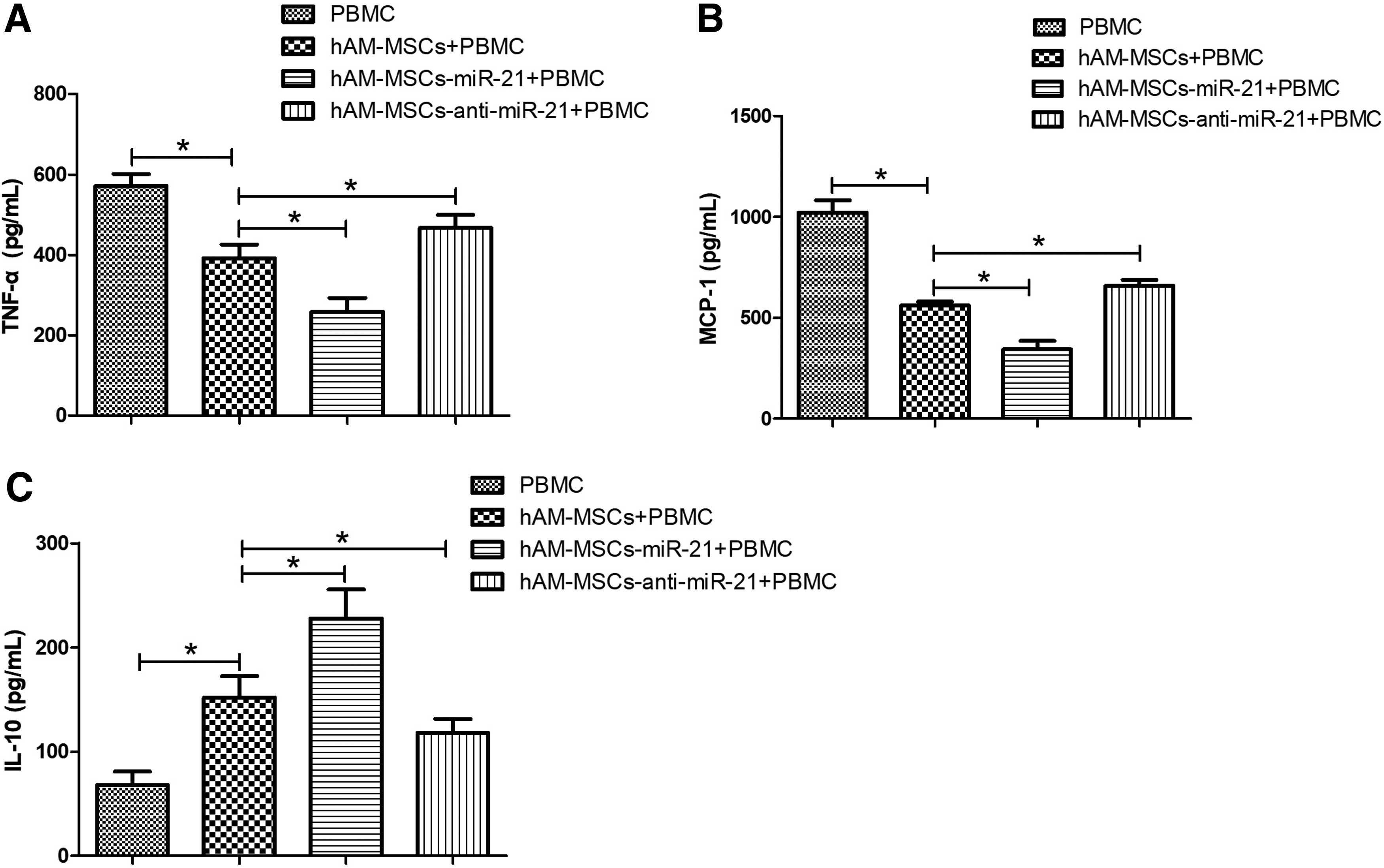
Upregulation of miR-21 promoted the immunoregulatory effect of hAM-MSCs on PBMC. To assess the effects of hAM-MSCs on cytokine secretion, PBMC were treated with PHA (1 μg/μL) for 24 h and then the cytokine quantification was performed.
Discussion
miRNAs are considered crucial gene regulators; they function by negatively regulating gene expression at the posttranscriptional level through translation repression and/or mRNA degradation (Matysiak et al., 2013). There is increasing evidence that has shown that miRNAs are involved in many biological processes and thus play important roles in many disorders, such as cancers, cardiovascular diseases, and multiple autoimmune diseases (Kataoka and Wang, 2014; Murugaiyan et al., 2015; Loginov et al., 2018). With regard to immunoregulation, miRNAs have been reported to be involved in the regulation of some immune responses in both innate and acquired immunity (Matysiak et al., 2013). Murugaiyan et al. (2015) have reported that miR-21 promotes T-helper 17 differentiation by targeting and depleting mothers against decapentaplegic homolog 7, a negative regulator of transforming growth factor beta signaling and mediates experimental autoimmune encephalomyelitis.
Multiple studies have revealed that the transplantation of MSCs for the treatment of neuronal and spinal disorders is becoming increasingly popular (Alviano et al., 2007; Heile et al., 2009). hAM-MSCs are considered a new and promising source of stem cells for cell replacement-based therapy (Yan et al., 2014). It has been reported that successful human amniotic membrane-derived-mesenchymal stem cells orthotropic transplantation into regions of the brain that has been damaged may be an effective treatment for brain injury (Wang et al., 2008). One study has revealed that miRNAs act as regulatory signals for preserving stemness and determining the fate of stem cells, including hMSCs (Trohatou et al., 2014). In this study, we successfully isolated hAM-MSCs that expressed the cell surface markers CD29, CD44, CD73, and CD90, as well as vimentin and SSEA-4 in hAM-MSCs, which is consistent with the previous findings (Zeng et al., 2011). Accordingly, we found that upregulation of miR-21 promoted proliferation and suppressed apoptosis of hAM-MSCs. Interestingly, our results revealed that hAM-MSCs significantly inhibited the secretion of TNF-α and MCP-1 and promoted the secretion of IL-10 in cocultured PBMC. These findings suggest that hAM-MSCs have immunosuppressive properties.
Many researches have reported that cytokines are involved in numerous diseases that have an inflammatory etiology. The expression of proinflammatory cytokines and chemokines, such as TNF, ILs, and C-C Motif Chemokine Ligand 2 (CCL2), may promote disease progression (Amor et al., 2010). Proinflammatory TNF-α, MCP-1, and anti-inflammatory IL-10 were chosen to evaluate the properties of certain immune responses (Deutsch et al., 2017; Walker et al., 2017). One such study disclosed that TNF can increase inflammation and contribute to the release of reactive oxygen species from microglia, which promotes neurodegeneration in the nervous tissue (Figiel, 2008). It has been shown that anti-inflammatory IL-10, in part, represses inflammation and negatively modulates immune responses (Liu et al., 2016a, 2016b). Another study has shown that exogenous IL-10 inhibits cisplatin nephrotoxicity, which is concerned with inflammatory reactions (So et al., 2007). MCP-1 is an influential chemotactic factor for blood monocytes and is secreted by diverse array of inflammatory cells (Lee et al., 2014). MCP-1 can recruit specific monocytes/macrophages and lymphocytes, which are involved in disease progression (Russo et al., 2011; Barnes, 2016). Woo et al. (2015) has demonstrated that IL-10/HMOX1 signaling is protective against cochlear inflammation by repressing MCP-1/CCL2 expression. TNF-α can induce the secretion of IL-6 and MCP-1 and exert anti-inflammatory effects on fibroblast-like synoviocytes (Li et al., 2015).
Aberrant expression of miRNAs has been reported to be involved in many human pathologies, including inflammatory diseases (O'Connell et al., 2010; Zare-Shahabadi et al., 2013). We therefore analyzed the expression of miR-21 in hAM-MSCs to correlate their immunoregulatory potential. PBMC contain predominantly (>80%) lymphocytes. Monocytes constitute less than <10%, while a population of granulocytes, erythrocytes, and platelets constitute less than 5% (Sharma et al., 2013). In many studies, PHA has been widely used to stimulate the activation of lymphocytes in PBMC, and cytokine secretion of lymphocytes can be assessed to study immune regulatory effects (Upratya et al., 2010; Sharma et al., 2013; Diedrichs-Mohring et al., 2018). Cytokine secretion is not only attributable to lymphocytes, since the other cell populations present in PBMC can secrete cytokines (Morrell et al., 2014; Radjabova et al., 2015; Zhang et al., 2016). In searching for whether miR-21 might be of significance in the regulation of the immunoregulatory activity of hAM-MSCs, we further investigated that the secretion of TNF-α, MCP-1, and IL-10 in PBMC with PHA stimulation cocultured with the hAM-MSCs transfected with miR-21 overexpressing and repressing lentiviral vectors. Our data disclosed that miR-21 overexpression enhanced the immunoregulatory effect of hAM-MSCs in PBMC cocultured with miR-21-hAM-MSCs. In contrast, miR-21 downregulation reduced the immunoregulatory effect of hAM-MSCs on the secretion of TNF-α, MCP-1, and IL-10 in PBMC cocultured with anti-hAM-MSCs. These results concluded that hAM-MSCs were identified as cells with a powerful immunoregulatory capacity and that miR-21 overexpression promoted immunoregulatory effect of hAM-MSCs on PBMC.
Conclusion
This study demonstrated that miR-21 promoted proliferation and inhibited apoptosis of hAM-MSCs and played a vital role in the control of the immunoregulatory capacity of hAM-MSCs on PBMC. The hAM-MSCs in this study were obtained from a single donor; thus additional studies using hAM-MSCs from multiple donors will need to be conducted to verify the broader applicability of these findings. Whether miR-21-mediated hAM-MSCs therapy can provide an applicable and safe strategy for immune-related disorders in the clinical setting also requires further investigation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.