
Abstract
Background:
To investigate the association between long-chain noncoding RNA (lncRNA) plasmacytoma variant translocation 1 (PVT1) gene polymorphisms and the risk and prognosis of colorectal cancer (CRC) in the Han Chinese population.
Methods:
A cohort of 225 Han Chinese CRC patients and 225 healthy controls was analyzed for the PVT1 gene genotypes at the loci rs1221464062, rs1366023633, rs1252200336, and rs1273526412. The levels of PVT1 mRNA, hsa-miR-455-5p, and hsa-miR-455-3p in both cancerous tissues and paracancerous normal tissues of the CRC patients were determined by reverse transcription polymerase chain reaction. The survival rate of CRC patients was recorded after 3 years of follow-up.
Results:
The risk of developing CRC in subjects with the ID genotype of the PVT1 gene locus rs1252200336 was 2.71 times higher compared with the type II genotype; and the D allele was a risk factor for CRC. The survival of CRC patients with the ID/DD genotypes of the PVT1 gene rs1252200336 locus was significantly lower compared with the II genotype. In both the cancerous tissues and the paracancerous normal tissues of CRC patients, the level of lncRNA PVT1 was negatively correlated with the hsa-miR-455-5p and hsa-miR-455-3p levels. In subjects carrying the ID/DD genotypes of the PVT1 gene rs1252200336 locus, the level of the lncRNA PVT1 in cancerous tissues was significantly higher compared with the II genotype, whereas the hsa-miR-455-3p level was significantly lower compared with the II genotype.
Conclusion:
The PVT1 gene rs1252200336 locus polymorphisms are associated with the risk of developing CRC in the Han Chinese population. The rs1252200336 locus deletion mutation (D) may impact the binding of hsa-miR-455-5p to the lncRNA PVT1 and its role in the development and progression of CRC.
Introduction
Colorectal cancer (CRC) is the most common digestive tract cancer in the world (Brenner et al., 2014). It is widely recognized that alterations to one's diet, environment, and other factors can increase the incidence and mortality of CRC (Brenner et al., 2014; Siegel et al., 2017). During the early stages of CRC, symptoms are not obvious. Therefore, most patients are already in the middle or late stage at the time of diagnosis. Clinically, the primary treatments for CRC include surgery, chemotherapy, radiotherapy, and targeted therapy (Navez et al., 2016; Punt et al., 2017; Moridikia et al., 2018). The survival rate, however, is relatively low for patients who are already in an advanced stage at the time of diagnosis (Kim et al., 2012; Xie et al., 2017). It is therefore particularly important to investigate the pathogenesis of CRC and search for biomarkers and targets for earlier diagnoses, which should lead to improved prognoses.
In mammals, very few genes encode proteins; the majority of the genomes encode noncoding RNAs (ncRNAs) (Johnson et al., 2005; Furuno et al., 2006). Recently, a body of evidence has been developed demonstrating that abnormalities in long-chain noncoding RNAs (lncRNAs) are closely correlated with the occurrence and development of CRC (Chen et al., 2017; Yang et al., 2017; Zhang et al., 2017). The plasmacytoma variant translocation 1 (PVT1) gene is located on chromosome 8q24.21 and is highly associated with CRC (Fan et al., 2018). Previously, a high expression level of PVT1 was detected in patients with a deep invasion of gastric cancer cells and patients with late-stage gastric cancer (Kong et al., 2015). In addition, increased expression levels of PVT1 were found in patients with late-stage nonsmall cell lung cancer and lymph node metastases (Yang et al., 2014). Prior studies have also shown that PVT1 expression levels in CRC patients are associated with tumor size, histological grade, metastasis, poor prognosis, and cisplatin resistance (Ping et al., 2018). PVT1 regulates the growth of CRC tumors through action as a competitive endogenous RNA (ceRNA) and negatively regulates miR-455; furthermore, the RUNX2/PVT1/miR-455 regulatory axis plays an important role in CRC tumorigenesis and serves as a therapeutic target for CRC (Chai et al., 2018).
It is still unclear whether PVT1 gene polymorphisms influence the functions regulating the occurrence and development of CRC. As predicted by the starBase database, there are polymorphisms within lncRNA PVT1 at the binding site of the miRNAs hsa-miR-455-5p and hsa-miR-455-3p. The former groups include the s1221464062 and rs1366023633 polymorphic loci, while the latter binding sites include the rs1252200336 and rs1273526412 polymorphic loci. This study was carried out in a case-control manner to investigate the relationship between gene polymorphisms at the predicted binding sites on PVT1 for the hsa-miR-455-5p and hsa-miR-455-3p miRNAs and the risk of developing CRC in Han Chinese patients.
Materials and Methods
Ethical aspects
This study was approved by the corresponding ethics committees of the Second Affiliated Hospital of Zhejiang Chinese Medical University. All of the participants signed the written informed consent form. This study was also conducted in accordance with the World Medical Association Declaration of Helsinki.
Subjects
A total of 225 patients suffering from CRC, who underwent radical resection of their tumors, were consecutively recruited from the Second Affiliated Hospital of Zhejiang Chinese Medical University between August 2014 and February 2016. All the patients were diagnosed with colorectal adenocarcinoma by endoscopy and histological pathology before surgery. Postsurgery, the pathological results were confirmed by two experienced surgical pathologists in the hospital, who were blinded to each other's assessments. The inclusion criteria were as follows: histologically confirmed CRC patients at stage II (T2 and T3, N0, M0) or stage III (any T, N1 and 2, M0). Patients who received chemotherapy and radiotherapy before surgery were excluded. The age of the patients in the CRC group ranged from 41 to 87 years with a mean age of 62.3 ± 11.1 years. All CRC patients were followed for 3 years or until death and their survival time was recorded. Concurrently, 225 healthy controls were recruited from the physical examination center, and ranged in age between 35 and 87 years with a mean of 62.2 ± 10.5 years. The clinical characteristics of all participants were collected and included age, gender, tumor location, clinical stage, smoking, and drinking status.
Genotyping
We collected 5 mL of venous blood from all participants and extracted their DNA using the QIAamp DNA Blood Mini kit (Qiagen, Valencia, CA). Genomic DNA was stored at −20°C until ready for use. The polymerase chain reaction (PCR) was performed using the following primers for the PVT1 gene: 5′-GCG AGC AAA GAT GTG CCC C-3′ (forward) and GAC CGC CAA CAT CCT TTC CG-3′ (reverse). The PCR was conducted in a total volume of 20 μL containing 100 ng of genomic DNA, 2 μL of 10 × PCR buffer, 1.5 μL of 25 mM Mg2+, 1 μL of 10 μM dNTP, 1 μL of 10 pM of each primer, and 0.25 μL of DNA polymerase, and sterile water was added to a final volume of 20 μL. The PCR conditions were as follows: 95°C for 5 min, followed by 94°C for 20 s, 58°C for 30 s, and 72°C for 30 s for 30 cycles; this was then followed by a final extension at 72°C for 5 min before storage at 4°C. After completion of the PCR, Sanger sequencing was performed to analyze the PVT1 gene rs1221464062, rs1366023633, rs1252200336, and rs1273526412 loci.
Real-time quantitative PCR
The total RNA of 225 surgically resected CRC patients was extracted from cancerous tissues and paracancerous normal tissues located ∼2 cm away from the tumor tissue, using TRIzol reagent (Invitrogen, CA) according to the manufacturer's instructions. The first-strand cDNA was synthesized using the RevertAid First Strand cDNA Synthesis kit (Thermo Fisher). The real-time quantitative PCR (qRT-PCR) was conducted on an ABI 7500 system (Applied Biosystems, Foster City, CA) using specific primers (Qiagen, Hilden, Germany) and a SYBR-Green PCR Master Mix kit (Takara). The primer sequences for glyceraldehyde 3-phosphate dehydrogenase (GAPDH, internal reference) were 5′-CCT CGT CTC ATA GAC AAG ATG GT-3′ (forward) and 5′-GGG TAG AGT CAT ACT GGA ACA TG-3′ (reverse). The primer sequences for PVT1 were 5′-TGA GAA CTG TCC TTA CGT GAC C-3′ (forward) and 5′-AGA GCA CCA AGA CTG GCT CT-3′ (reverse). The mRNA expression level of PVT1 was normalized to the mRNA expression level of GAPDH. The primer sequences for U6 were 5′-CTC GCT TCG GCA CA-3′ (forward) and 5′-AAC GCT TCA CGA ATT TGC GT-3′ (reverse). The primer sequences for hsa-miR-455-5p were 5′-ATG TGC CTT TGG ACT ACA TCG AA-3′ (forward) and 5′-TTA CTA CGT CAT GAC TAG TAA-3′ (sequences for hsa- reverse). The primer miR-455-3p were 5′-GCAGTCCATGGGCATATACAC-3′ (forward) and 5′-TAT GGA ACG CTT CAC GAA TTT GC-3′ (reverse). The expression levels of the miRNAs were normalized to the expression level of U6.
Statistical analyses
The statistical analyses were performed using SPSS 20.0 software (SPSS, Inc., Chicago). The continuous parameters and nonparametric data are expressed as (mean ± SD), and the categorical variables are expressed as n (%). Pearson's chi-squared test was utilized to compare the categorical variables, the mean of the normally distributed continuous variables was compared by ANOVA, and the nonparametric statistical analysis between the two groups was compared by Student's t-test. For genotype frequencies, the Hardy-Weinberg equilibrium was performed using the χ2 test, and the odds ratio (OR) together with the 95% confidence interval (CI) was used to analyze the correlation between genotypes and allele frequencies and the risk of developing CRC in subjects. A multivariate logistic regression analysis was performed to correct for age, gender, smoking, drinking, and body mass index (BMI). Pearson's correlation was performed to analyze the association of the lncRNA PVT1 level with the hsa-miR-455-5p and hsa-miR-455-3p levels. The survival curves were plotted using the Kaplan-Meier method, and the log-rank test was used to analyze the survival time difference. A p < 0.05 was considered statistically significant.
Results
Clinical characteristics of the participants
The general clinical characteristics of both the CRC patients and the healthy controls are shown in Table 1. Among the 225 patients suffering from CRC, 22 had tumors in the left colon, 78 in the right colon, and 125 in the sigmoid colon and rectum. There were 12 cases of TNM stage I, 85 cases of stage II, 94 cases of stage III, and 34 cases of stage IV. No significant difference was detected in the general data of age, gender, BMI, smoking status, and drinking status of CRC patients compared to controls (p > 0.05).
The Basic Characteristics of Colorectal Cancer Patients and Healthy Controls
The data are expressed as mean ± SD or n (%).
BMI, body mass index; CRC, colorectal cancer; TNM, tumor node metastasis.
Association of PVT1 gene polymorphisms and the risk of developing CRC
The distributions of the genotypes and the allele frequencies of the PVT1 gene polymorphisms in the CRC group and the control group are shown in Table 2. The frequency distributions of the PVT1 rs1221464062, rs1366023633, rs1252200336, and rs1273526412 genotypes are in agreement with the Hardy-Weinberg equilibrium (p > 0.05). There was no significant difference in the genotypes and allele frequencies between the CRC group and the control group for the PVT1 gene rs1221464062, rs1366023633, and rs1273526412 loci (p > 0.05). However, the risk of developing CRC in patients with the ID genotype of the PVT1 gene rs1252200336 locus was 2.71 times higher compared with subjects with the type II genotype (95% CI: 1.30-5.62, p = 0.01); and the risk of developing CRC in the subjects with D allele was 4.99 times higher compared with the I allele (95% CI: 2.56-9.72, p < 0.001).
The Association of PVT1 Genotypes and Allele Frequencies with the Risk of Developing Colorectal Cancer in Subjects
Multivariate logistic regression analysis to correct factors of age, gender, smoking, drinking, and BMI.
Bold values indicate p < 0.05.
CI, confidence interval; CRC, colorectal cancer; D, delete; HWE, Hardy-Weinberg equilibrium; I, insert; OR, odds ratio (OR >1 indicates a risk factor; OR <1 indicates a protective factor; and OR = 1 indicates that this factor does not contribute to the occurrence of the disease).
Association of PVT1 gene polymorphisms and the prognosis of CRC patients
There was no association between the PVT1 gene rs1221464062, rs1366023633, and rs1273526412 loci polymorphisms and the 3-year survival of CRC patients (Fig. 1A, B, and D, respectively). It is important to note that the survival rate of CRC patients with the PVT1 gene rs1252200336 locus ID/DD genotype was significantly lower compared with patients with the type II genotype (hazard ratio = 2.14, 95% CI: 1.04-4.42, p < 0.001) (Fig. 1C).

Correlation between PVT1 gene polymorphisms and the prognosis and survival of patients with CRC.
Comparison of lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p levels
We further detected lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p levels in both cancerous and paracancerous normal tissues from CRC patients by RT-PCR; the results are shown in Figure 2. The level of the lncRNA PVT1 in cancerous tissues of CRC patients was significantly higher than in paracancerous tissues, while the levels of hsa-miR-455-5p and hsa-miR-455-3p were significantly lower than those in paracancerous tissues (p < 0.05).
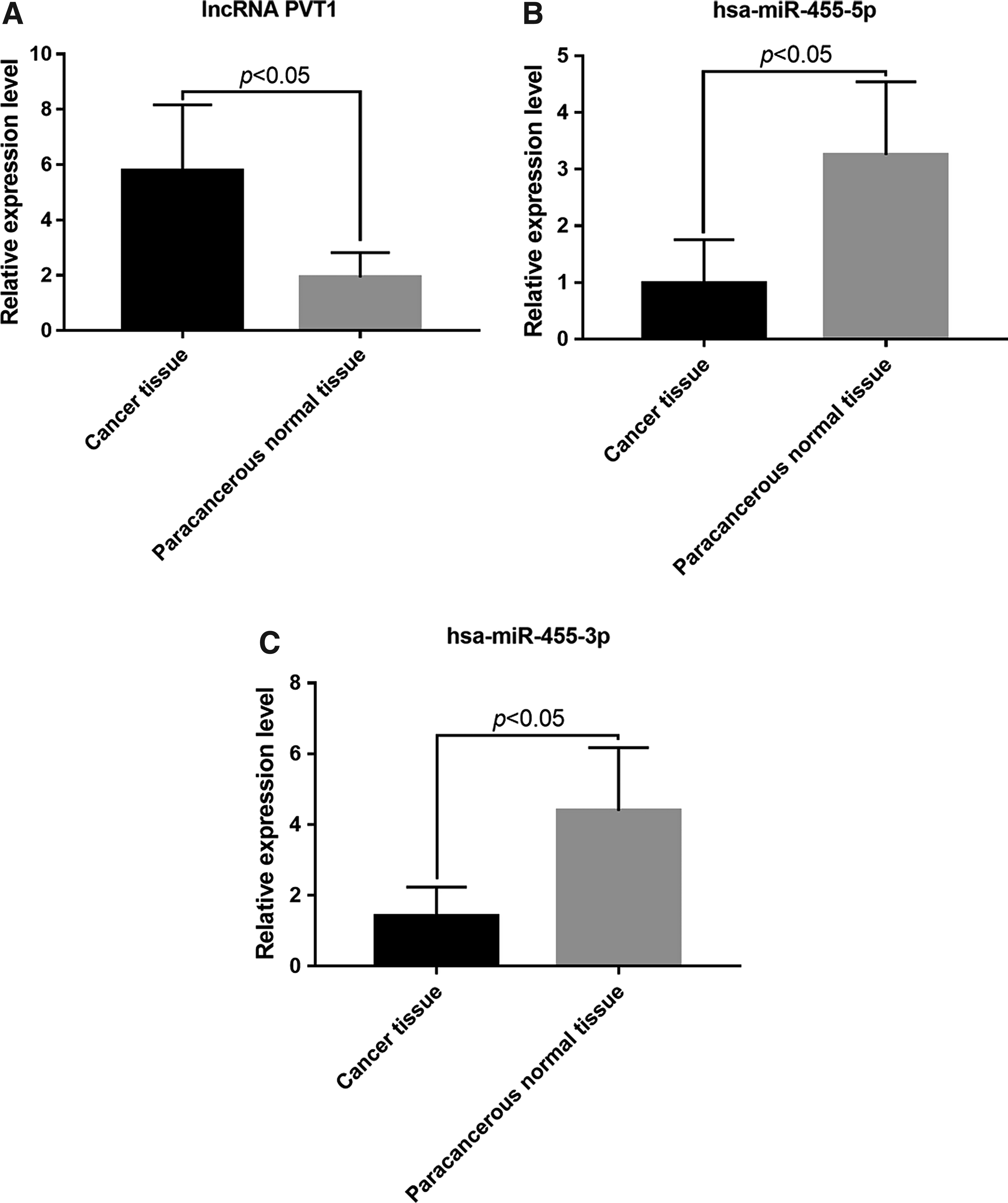
Comparison of lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p levels in the cancerous tissues and paracancerous tissues of CRC patients. The levels of
Correlation of the lncRNA PVT1 level with hsa-miR-455-5p and hsa-miR-455-3p levels
The level of lncRNA PVT1 in cancerous tissues of CRC patients was negatively correlated with the levels of the miRNAs, hsa-miR-455-5p and hsa-miR-455-3p (r = −0.64 and −0.71, respectively) (Fig. 3A, B). In addition, in the paracancerous tissues, the level of lncRNA PVT1 was negatively correlated with the hsa-miR-455-5p and hsa-miR-455-3p levels (r = −0.59 and −0.68, respectively) (Fig. 3C, D).

The level of lncRNA PVT1 was negatively correlated with hsa-miR-455-5p and hsa-miR-455-3p levels.
Correlation between PVT1 gene polymorphisms and the levels of lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p in cancer tissues
We further analyzed the correlation between the PVT1 gene polymorphisms and the levels of the lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p in the cancerous tissues of CRC patients. We found that none of the PVT1 gene rs1221464062, rs1366023633, and rs1273526412 loci polymorphisms significantly correlated with lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p levels in cancer tissues (p > 0.05) (Fig. 4A, B, D). The level of lncRNA PVT1 in the cancerous tissues of CRC patients was significantly higher in subjects with the ID/DD genotype of the PVT1 gene rs1252200336 locus than in the type II genotype. The level of hsa-miR-455-3p in the cancerous tissues of CRC patients with the ID/DD genotype of the rs1252200336 locus was significantly lower than that in the type II genotype (p < 0.05). However, there was no significant difference in hsa-miR-455-5p levels between cancerous tissues and paracancerous tissues (p = 0.45) (Fig. 4C).
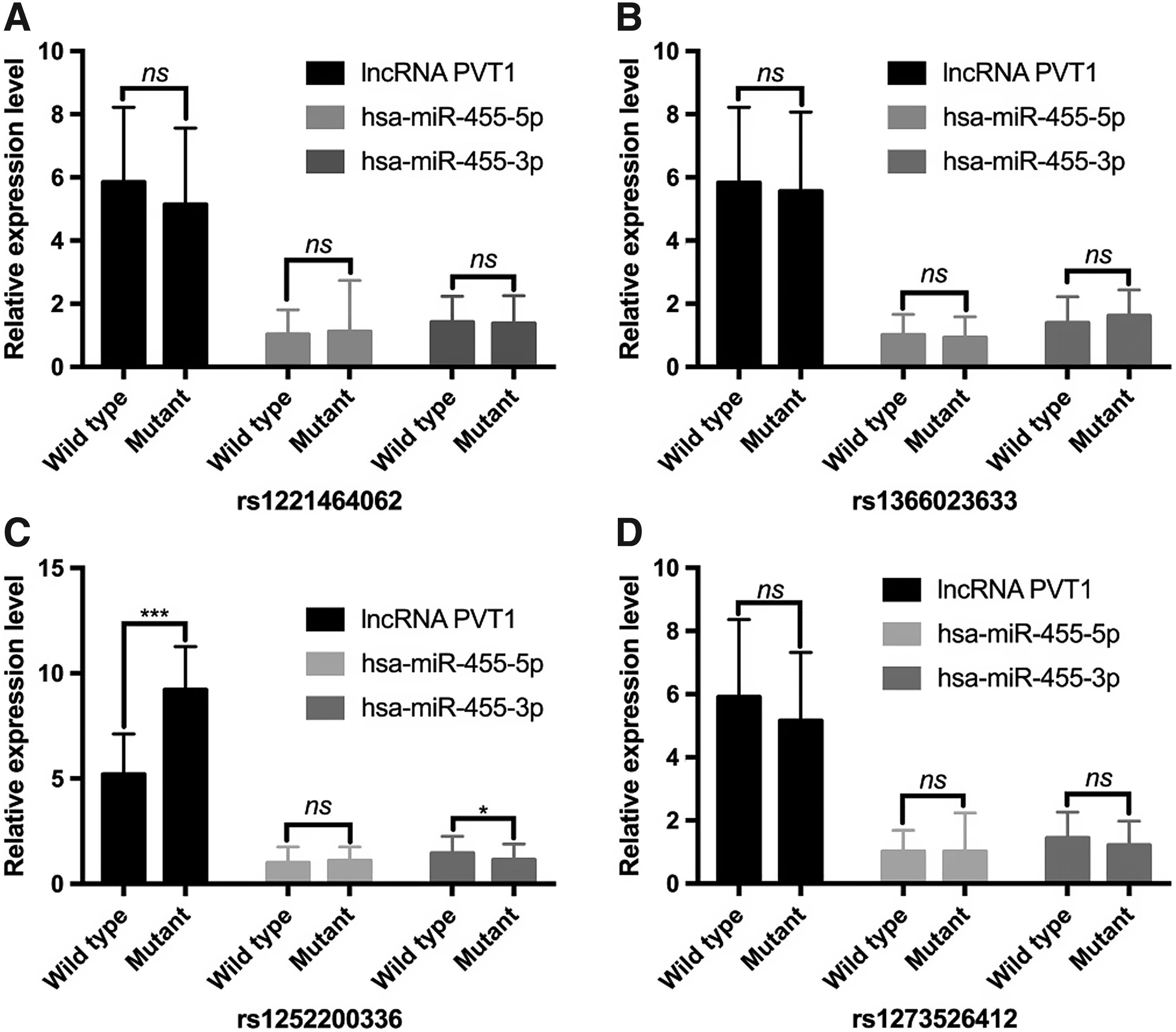
Correlation between PVT1 SNPs and the levels of lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p in the cancerous tissues of CRC patients.
The association of PVT1 gene polymorphisms with the levels of lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p in paracancerous normal tissues
We looked for correlations between the PVT1 gene polymorphisms in paracancerous normal tissues of CRC patients and the levels of lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p. No association was detected between the PVT1 gene rs1221464062, rs1366023633, and rs1273526412 loci polymorphisms and the levels of lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p in paracancerous normal tissues (p > 0.05) (Fig. 5A, B, D). The level of lncRNA in paracancerous normal tissues was significantly higher in patients carrying the ID genotype of the PVT1 gene rs1252200336 locus compared to those of the type II genotype (p < 0.05). The level of hsa-miR-455-3p in paracancerous normal tissues was significantly lower in subjects with the ID genotype of the rs1252200336 locus compared to those of the type II genotype (p < 0.05); there was, however, no significant difference in hsa-miR-455-5p levels in paracancerous normal tissues (p = 0.06) (Fig. 5C).
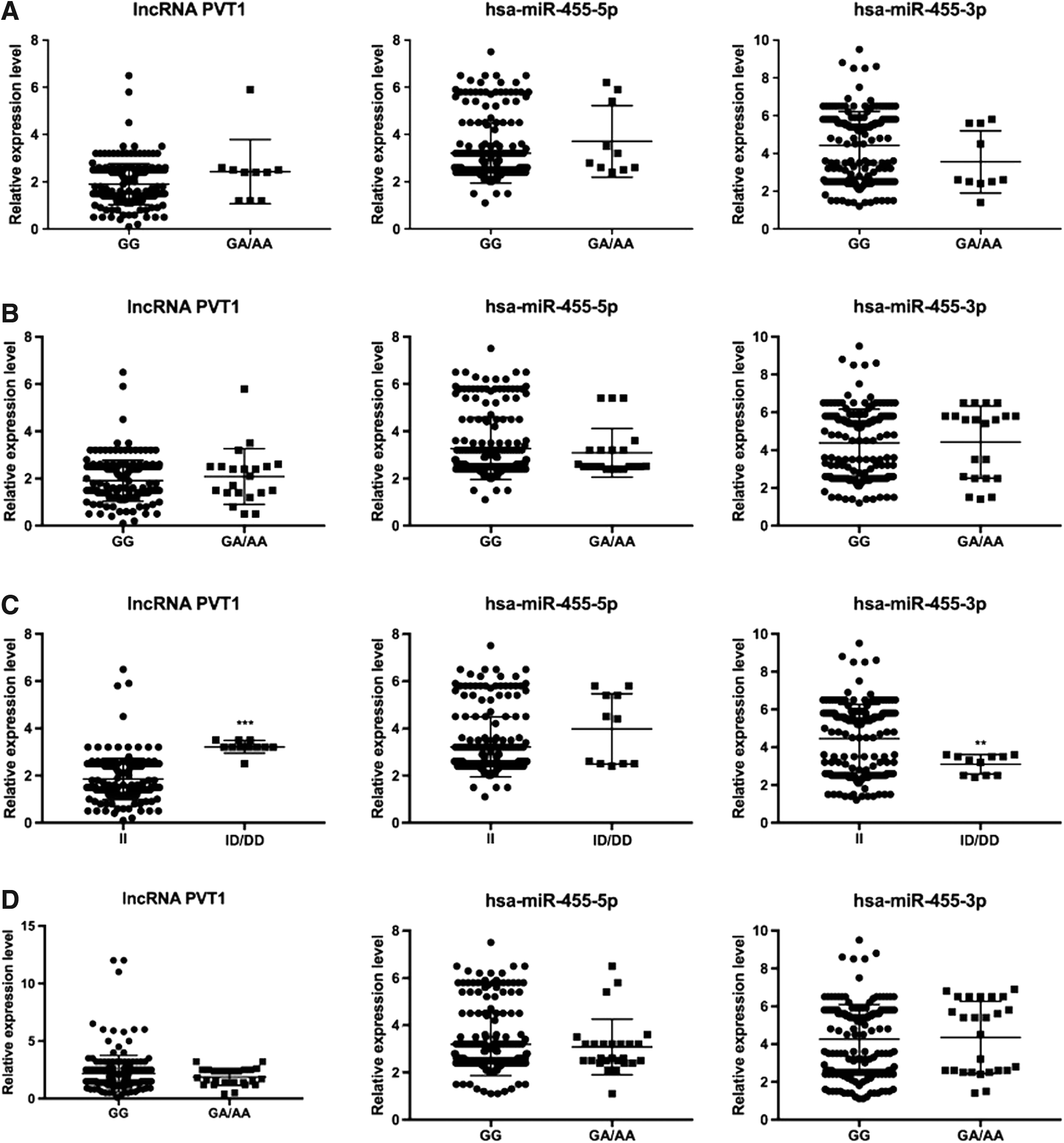
Correlation between the PVT1 SNPs and the levels of lncRNA PVT1, hsa-miR-455-5p, and hsa-miR-455-3p in paracancerous normal tissues.
Discussion
CRC is one of the most common malignant tumors in the world (Brenner et al., 2014) and seriously threatens human health. According to a survey, the incidence of CRC exceeds that of breast cancer and esophageal cancer, and is worldwide ranked third, while it ranks fourth in mortality in China (Chen et al., 2015). With the advances in genetic investigation and molecular mechanisms, our understanding of CRC is gradually increasing. In previous studies that aimed to search for early diagnostic biomarkers of CRC, it was shown that during early stages, manipulation of the expression levels of susceptibility genes and their signaling pathways could be used toward the treatment of CRC (Mahasneh et al., 2017; Zhang et al., 2019).
To date, diverse ncRNAs have been shown to play a core role in the development, progression, and metastasis of cancer (Whitehead et al., 2009). lncRNAs are ncRNAs that are ≥200 nucleotides in length and that regulate gene expression (Khalil et al., 2009; Guttman and Rinn, 2012); some have been shown to be involved in the incidence and development of tumors (Yang et al., 2011; Enfield et al., 2012; Zhao et al., 2014; Yue et al., 2016). According to the ceRNA hypothesis, lncRNAs are able to communicate with other RNA transcripts through miRNA responsive elements, which serve as one of the letters of the new RNA language (Zhang et al., 2018). A large body of evidence has been developed that shows the interaction between certain lncRNAs and miRNAs plays an important role in cancer progression (Yuan et al., 2017, 2018; Li et al., 2018). Therefore, it is imperative to investigate the interaction between lncRNAs and miRNAs to help understand the development and trajectory of CRC. The lncRNA PVT1 is a widely studied lncRNA in cancer research and is located near the MYC gene on human chromosome 8q24 (Beck-Engeser et al., 2008). It is likely that the lncRNA PVT1 interacts directly with the miRNAs hsa-miR-455-5p and hsa-miR-455-3p (Fig. 6). Consistent with a study by Chai et al. (2018), we showed that there were negative associations between the lncRNA PVT1 level and hsa-miR-455-5p and hsa-miR-455-3p levels. This indicates that the lncRNA PVT1 might act as a ceRNA and negatively regulate the expression of hsa-miR-455-5p and hsa-miR-455-3p, and thereby promote the occurrence and development of CRC.
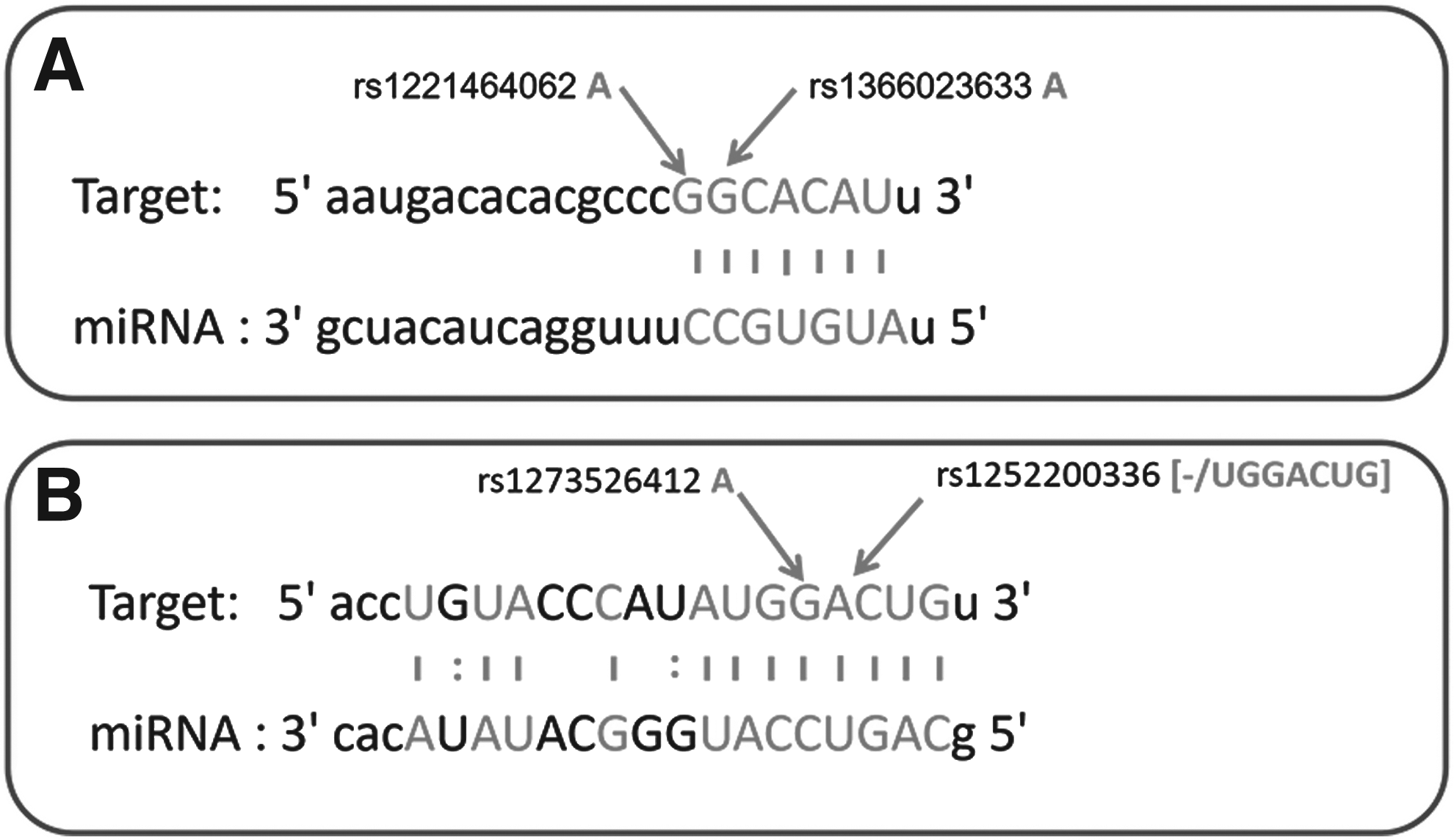
A prediction of binding sites for miRNA to lncRNA.
The interaction of lncRNA PVT1 with miRNAs is mainly through the complementary pairing of bases, which depends on the strength and efficiency of base pairing. In this study, the PVT1 gene single-nucleotide polymorphisms rs1221464062 and rs1366023633, which are located at the binding site of hsa-miR-455-5p, were evaluated to determine if the different allelic forms resulted in decreased binding efficiency. As a result, we found that the PVT1 gene rs1252200336 locus D polymorphism likely leads to the failure of hsa-miR-455-3p binding to lncRNA PVT1 as we found that this SNP was associated with a greatly increased risk (4.99-fold) of developing CRC. The survival period also was significantly decreased in CRC patients carrying this PVT1 gene rs1252200336 deletion. Further analyses showed that the level of lncRNA PVT1 was significantly increased in both cancerous tissues and paracancerous normal tissues of CRC patients with the D allele, while the level of hsa-miR-455-3p was significantly decreased. There was, however, no significant difference in the level of the hsa-miR-455-5p miRNA. It is therefore likely that for subjects with D allele, the binding of has-miR-455-3p to the lncRNA PVT1 is affected, which in turn influences its role in negative regulation and leads to the occurrence and development of CRC. Further studies are needed to verify the detailed mechanisms.
This study is not without its limitations. First, the negative regulation of the lncRNA PVT1 on hsa-miR-455-5p and hsa-miR-455-3p has not been verified in an in vitro model, and thus, in vitro cellular and murine models are required for further verification. In addition, there are additional miRNAs that target lncRNA PVT1. Therefore, it is also possible that other lncRNAs and miRNAs participate in the development and progression of CRC. In addition, when we collected the paracancerous tissues, it is possible that the tissues also contain a few cancer cells; thus, we cannot completely rule out the influence of tumor tissues, which could also have affected the objective of this study.
Conclusion
The PVT1 rs1252200336 polymorphism is associated with the risk and prognosis of CRC in the Han Chinese population. The rs1252200336 locus deletion mutation might affect the binding efficiency and negative regulation of hsa-miR-455-5p to lncRNA PVT1, and eventually result in the occurrence and development of CRC. However, further studies are warranted to verify this mechanism.
Footnotes
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by grants from Traditional Chinese Medical Science and Technology Plan of Zhejiang Province (2015ZB069).