
Abstract
Background:
E2F5 is a transcription factor that is overexpressed in the early stages of ovarian cancer and has been suggested as a potential biomarker for early detection. In this study, we aimed to examine the role of E2F5 in invasion and proliferation of ovarian cancer cells.
Materials and Methods:
We performed cell viability, colony formation, and invasion assays using ovarian cancer cells treated with siRNA to knock down the E2F5 gene. The regulatory effects of E2F5 on proteins involved in the apoptotic, Wnt, Hippo, and retinoblastoma signaling pathways were evaluated by western blotting following E2F5 repression. In addition, we analyzed data available on Gene Expression Profiling Interactive Analysis for correlations between E2F5 and YAP, β-catenin, cyclin D1, cdk4, and caspase-9.
Results:
E2F5 was highly expressed in ovarian cancer cell lines and samples when compared to the nonmalignant tissues. Downregulation of E2F5 inhibited cell viability and invasion and promoted the phosphorylation of YAP, GSK-3-β, β-catenin, and retinoblastoma. However, cyclin D1, cdk4, and caspase-9 were downregulated when compared to control.
Conclusion:
Overall, E2F5 promotes ovarian carcinogenesis via the regulation of Hippo and Wnt pathways
Introduction
Epithelial Ovarian Cancer (EOC) is a lethal gynecological malignancy (Chang et al., 2018) representing 90% of all types of ovarian cancer (Sankaranarayanan and Ferlay, 2006) and is considered the 8th leading cause of cancer-related deaths among women worldwide (Siegel et al., 2018). The majority (70-80%) of the EOC patients are diagnosed at advanced stages (III and IV) (Jelovac and Armstrong, 2011), which is primarily due to the asymptomatic nature of disease progression and absence of reliable biological markers to detect EOC at early stages (Zhou et al., 2019).
Transcription factors have a crucial role in regulating gene expression, and a thorough understanding of the mechanism involved in gene regulation is essential for identifying the genes that can be used as biomarkers. The transcriptional repressor, E2F5, is a member of the E2F family of transcription factors, and a regulator of G0/G1 phase of the cell cycle (Johnson et al., 2016) and its transcriptional activity is regulated through its binding to the retinoblastoma protein family (Rb, p130, and p107 pocket proteins) (Xanthoulis, 2013). Retinoblastoma proteins act as negative regulators of cell proliferation by forming a complex with E2F proteins and hence inhibiting the cell cycle progression (Xanthoulis, 2013). In G0 and early G1 phases, E2F4 and E2F5 are bound to p130 and have inhibitory effects on gene regulation (Tsantoulis and Gorgoulis, 2005).
Rb/E2F pathway is commonly deregulated in several types of cancers and plays a crucial role in normal cell proliferation (Nevins, 2001). Few studies have examined the role of E2F5 in cancer progression. In hepatocellular (Jiang, 2011), breast (Umemura et al., 2009), colorectal (Lassmann et al., 2006), and ovarian (Kothandaraman et al., 2010) cancers, E2F5 amplification was reported. E2F5 was found to be overexpressed in both early and advanced stages of ovarian cancer cell lines and tumor samples (Shridhar et al., 2001; Reimer et al., 2006), following the downregulation of a microRNA (miRNA-132) known to be involved in cancer development (Tian et al., 2016).
Serous and endometrioid ovarian carcinoma showed higher E2F5 expression when compared to other types of EOC, while no expression was observed in the normal or benign ovarian samples. Similarly, other studies showed a fivefold increase in E2F5 expression in both the early and late stages of EOC. Hence, E2F5, along with CA125, was found to increase the specificity of EOC detection to 72.5%, thereby serving as a potential biomarker (Kothandaraman et al., 2010).
In this study, we examined the cross-talk between E2F5 and the Hippo/Wnt-β-catenin pathways and its effect on the cell cycle.
Materials and Methods
Cell culture
The human ovarian cancer cell lines: A2780 cisplatin sensitive (cps) (ECACC catalog no. 93112519) and A2780 cisplatin-resistant (cpr) cell lines (ECACC catalog no. 93112517) were a generous gift from Dr. Benjamin Tsang, University of Ottawa, Canada, while Prof. Aikou Okamoto kindly provided MCAS (JCRB cell bank, Osaka, Japan, catalog no. JCRB0240) and OVSAHO (JCRB cell bank, Osaka, Japan, catalog no. JCRB1046) cell lines from Jikei University School of Medicine, Japan. HOSE 6-3 cells were kindly provided by Prof. GSW Tsao from the School of Biomedical Sciences, The University of Hong Kong. OVSAHO was cultured in RPMI-1640 media, while MCAS, A2780 cpr, and A2780 cps cells were propagated in Dulbecco's modified Eagle's medium (DMEM) (Gibco, NY) supplemented with 10% fetal bovine serum (FBS) (Gibco) and 1% penicillin-streptomycin antibiotic (Gibco) in a humidified incubator at 37°C and 5% CO2.
RNA extraction and real-time-quantitative polymerase chain reaction
Total RNA was extracted from the ovarian cancer cell lines using the PureLink RNA mini kit (Invitrogen, CA) according to the manufacturer's protocol, and subsequently reverse transcribed to complementary DNA using high-capacity reverse transcription kit (Invitrogen). Briefly, TaqMan Expression Master Mix (10 μL) was added to 100 ng of cDNA and 1 μL of TaqMan Gene Expression Assay containing preoptimized primers and probes (E2F5: catalog no. Hs00231092_m1, GAPDH: catalog no. 402869; Thermo Scientific Fisher). Water was added to reach a total volume of 20 μL, and then qPCR was carried out in ABI 7500 Fast real-time PCR machine (Applied Biosystems, Austin, TX). The following program was applied: uracil N-glycosylase (UNG) incubation at 50°C for 2 min, then 20 s at 95°C for polymerase activation followed by 40 PCR cycles. In each cycle, cDNA was denatured for 3 s at 95°C, followed by 30 s incubation at 60°C for annealing/elongation. All the messenger RNA data were normalized to GAPDH expression, and relative expression was computed by the ΔΔCq method (Livak and Schmittgen, 2001).
Cell transfection with siRNA
MCAS and OVSAHO (1.5 × 105 to 3 × 105 cells) were seeded in a six-well plate and incubated until they reached 60% confluency. Then it was transfected with commercially available predesigned E2F5 siRNA (catalog no. 4392420, Thermofisher, Inc.), which is a pool of 3 target-specific 19-25 nucleotide siRNAs designed to act on different positions along the mRNA to avoid off-target effects. Lipofectamine RNAiMAX reagent (Invitrogen; Thermofisher Scientific) and Opti-MEM medium (Invitrogen) were used according to the manufacturers' protocol. The cells were then incubated for 36 h at 37°C and 5% CO2 atmosphere before use for RNA and protein extraction.
In vitro cell proliferation assay
Alamar blue reagent (Invitrogen) was used to measure cell viability. MCAS cells were seeded in 96-well culture plates at a density of 104 cells per well and transfected one day later with E2F5 siRNA and incubated for 24-96 h at 37°C and 5% CO2. Cell proliferation was assessed by adding 10 μL Alamar blue dye to the culture medium and incubating for 4 h at 37°C. The absorbance was measured at 570 nm using the Multiscan spectrum spectrophotometer (Thermo Fisher Scientific, Inc.).
Soft agar colony-forming assay
For soft agar colony formation assay, 1 × 105 MCAS and OVSAHO cells per well were treated with E2F5 siRNA (36 h post-transfection) and resuspended in 0.3% soft agar in culture medium and layered onto 0.6% solidified agar in a six-well plate. After 2 weeks of incubation, colonies were stained with crystal violet and counted in three random fields at 40 × magnification.
Cell invasion assay
The invasive potential of MCAS cells following the knockdown of E2F5 was evaluated using QCM™ Collagen-cell-invasion-assay kit 24-well 8 μm, colorimetric (Sigma Aldrich) following the manufacturer's protocol. Briefly, 1.25 × 105 E2F5-siRNA-transfected cells were suspended in 250 μl of serum-free media and loaded on the upper chamber of the insert. About 500 μl of the medium supplemented with 10% FBS was added to the lower chamber. After a prolonged incubation of 48 h, noninvading cells on the upper surface were removed with a cotton swab, and cells invading the lower chamber were stained with crystal violet and treated commercially provided extraction solution (QCM Collagen cell invasion assay kit 24-well 8 μm, colorimetric, Sigma Aldrich). The absorbance was measured at 560 nm using the Multiscan spectrophotometer (Thermofisher Scientific, Inc.).
Western blotting
E2F5 knocked-down MCAS cells (36 h post siRNA transfection) were subjected to protein extraction using RIPA lysis buffer (Santa Cruz Biotechnology, Inc.) supplemented with proteases and phosphatases inhibitor (Thermofisher). The lysate was centrifuged, and the supernatant was recovered and stored at −80°C until use. After protein quantification, ∼40 μg of proteins were resolved on 8% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and electro-transferred onto a nitrocellulose membrane, which was incubated in a blocking reagent (5% bovine serum albumin in tris-buffered saline/Tween-20 buffer [TBST]) for 1 h at room temperature. Then incubated with the primary antibodies (polyclonal, rabbit anti-human antibodies) directed against the following proteins Rb (sc-50, Santa Cruz Biotechnology), phospho-retinoblastoma (pRb; PA5-37715; Invitrogen, Thermofisher), actin (sc-1616, Santa Cruz Biotechnology), cdk 4 (sc-260, Santa Cruz Biotechnology), E2F5 (sc-999, Santa Cruz Biotechnology); caspase 9 (PA5-22252; Invitrogen, Thermofisher), β-catenin (PA5-77934; Invitrogen, Thermofisher), phospho-β-catenin (pβ-catenin; PA5-67504; Invitrogen, Thermofisher), E cadherin (PA5-32178; Invitrogen, Thermofisher), vimentin (PA5-27231; Invitrogen, Thermofisher), GSK-3-β (#9315, Cell Signaling Technology), phospho-GSK-3-β (pGSK-3-β; #5558, Cell Signaling Technology), and cyclin D1 (ab134175; Abcam) at a dilution of 1:200. Antibody against beta-actin was used as a loading control. After washing 3 × 5 min in TBST, membranes were incubated with horseradish peroxidase-conjugated secondary antibody (dilution 1:5000) for 2 h at room temperature and revealed by using enhanced chemiluminescence western blotting detection reagent (Pierce Biotechnology, IL).
Bioinformatics and statistical analysis
We explored Gene Expression Profiling Interactive Analysis (GEPIA, accessed on July 14, 2019), available public database to examine the expression of E2F5 at the RNA level in 423 human ovarian cancer and 88 healthy ovarian samples. A Pearson correlation analysis in 200 patients' samples was performed using the GEPIA database to determine the correlation between E2F5 and YAP, cdk4, cyclin D1, and β-catenin.
Data represent mean and standard deviation from at least three independent experiments performed in triplicates. Student's t-test was used for the comparison in the dataset between two groups, while one-way analysis of variance (ANOVA) (post hoc test: Dunnett's test) was used for the dataset containing more than two groups using SPSS software (version 23, SPSS, Inc., Chicago, IL) and in GraphPad Prism (version 8.1.2 GraphPad Software, Inc., San Diego, CA). Statistical significance was set at p < 0.05 (p < 0.05 [*], p < 0.01 [**], p < 0.001 [***], and p < 0.0001 [****]).
Results
E2F5 expression at proteins and mRNA levels
We evaluated the expression of E2F5 in ovarian cancer cell lines (MCAS, OVSAHO, A2780-cps, A2780-cpr), and in normal ovarian cell line (HOSE 6-3), using real-time-quantitative polymerase chain reaction and western blotting. E2F5 was significantly upregulated at the mRNA level in A2780-cps and A2780-cpr cell lines (p = 0.0029 and p = 0.0031, respectively). However, the MCAS line displayed lower expression of E2F5 (p = 0.0414), and no significant difference in expression was found in OVSAHO when compared to normal cells (HOSE 6-3). Similar results were obtained by western blot analysis of E2F5 protein expression in ovarian cancer cells. E2F5 was highly expressed in the A2780 cps cell line, followed by OVSAHO, A2780 cpr, MCAS (p < 0.0001), and HOSE 6-3 cell lines (Fig. 1a). Analysis of E2F5 expression in 426 ovarian tumors and 88 nonmalignant samples from the GEPIA public database revealed the high mRNA expression of E2F5 in ovarian tumors compared to healthy tissues (Fig. 1b), which is consistent with our findings in ovarian cell lines.

Relative expression of E2F5 at the mRNA and the protein levels in ovarian cancer cell lines.
E2F5 downregulation inhibited proliferation and colony formation
To evaluate the role of E2F5 on ovarian cancer cell proliferation, we knocked down E2F5 in MCAS and OVSAHO lines, using siRNA (Fig. 1c and d). The effect of silencing E2F5 on cell survival was assessed by Alamar blue assay. Results showed that the transient knockdown of E2F5 significantly inhibited cell proliferation in MCAS cells (p = 0.021) (Fig. 2a). Furthermore, anchorage-independent growth in E2F5-siRNA treated MCAS (p = 0.0369), and OVSAHO (p = 0.022) cells was lower when compared to control (Fig. 2b).
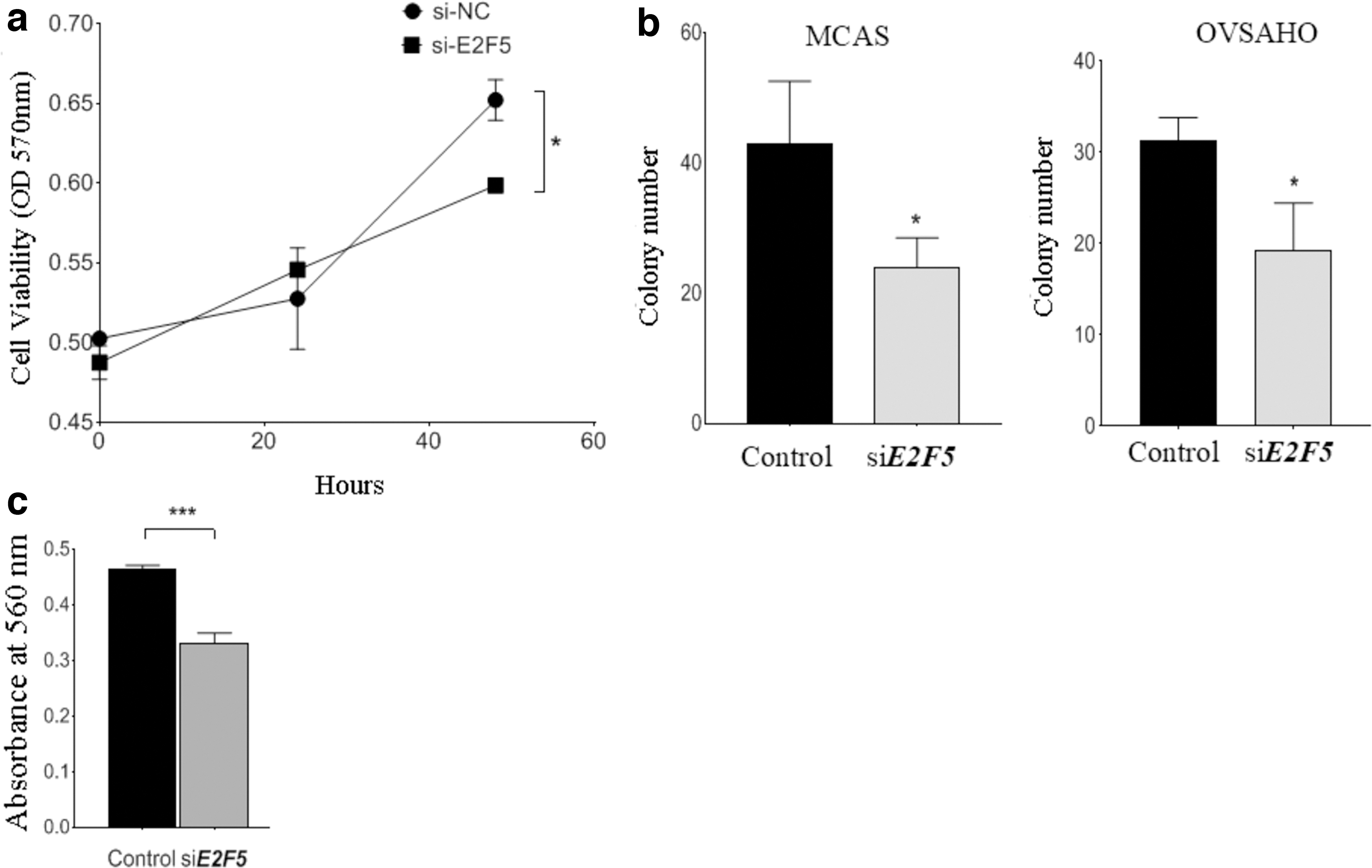
Functional role of E2F5 on cell proliferation, colony formation, and invasive ability.
E2F5 repression antagonizes invasion
We performed cell invasion assay to investigate whether E2F5 regulates invasiveness in ovarian cancer cell lines. Results showed that the invasive ability of MCAS ovarian cancer cells reduced significantly following the E2F5 knockdown (p = 0.0003, respectively), suggesting its oncogenic ability to promote invasion in vitro (Fig. 2c).
Effects of E2F5 on epithelial-mesenchymal transition, Hippo, apoptotic, cell cycle, and Wnt β-catenin signaling pathways
We examined the effect that E2F5 can exert on the tumor characteristics of ovarian cancer cells and regulation of proteins involved in epithelial-mesenchymal transition, Hippo, apoptotic, retinoblastoma, and Wnt-β-catenin pathways by western blotting. Relative expression variation and the protein bands in control, and E2F5 knocked MCAS cells are shown in Figure 3a and b, respectively, while the ratio of phosphorylated to total proteins are shown in Figure 3c. We observed no significant difference in the expression of the epithelial marker, E-cadherin in cells treated with E2F5-siRNA. In contrast, the expression of the mesenchymal marker, vimentin, was higher in E2F5-siRNA treated cells than in nontreated cells (Fig. 3a and b). In E2F5-siRNA treated cells, the level of phosphorylated YAP (p = 0.0138), GSK-3-β (p = 0.0004), β-catenin (p = 0.0285), phosphorylated β-catenin (p = 0.0001), and retinoblastoma (p = 0.0338) was higher with respect to control. On the other hand, the expression of cyclin D1 (p = 0.0002), cdk4 (p = 0.0082), and caspase-9 (p = 0.0033) were lower compared to the control (Fig. 3).
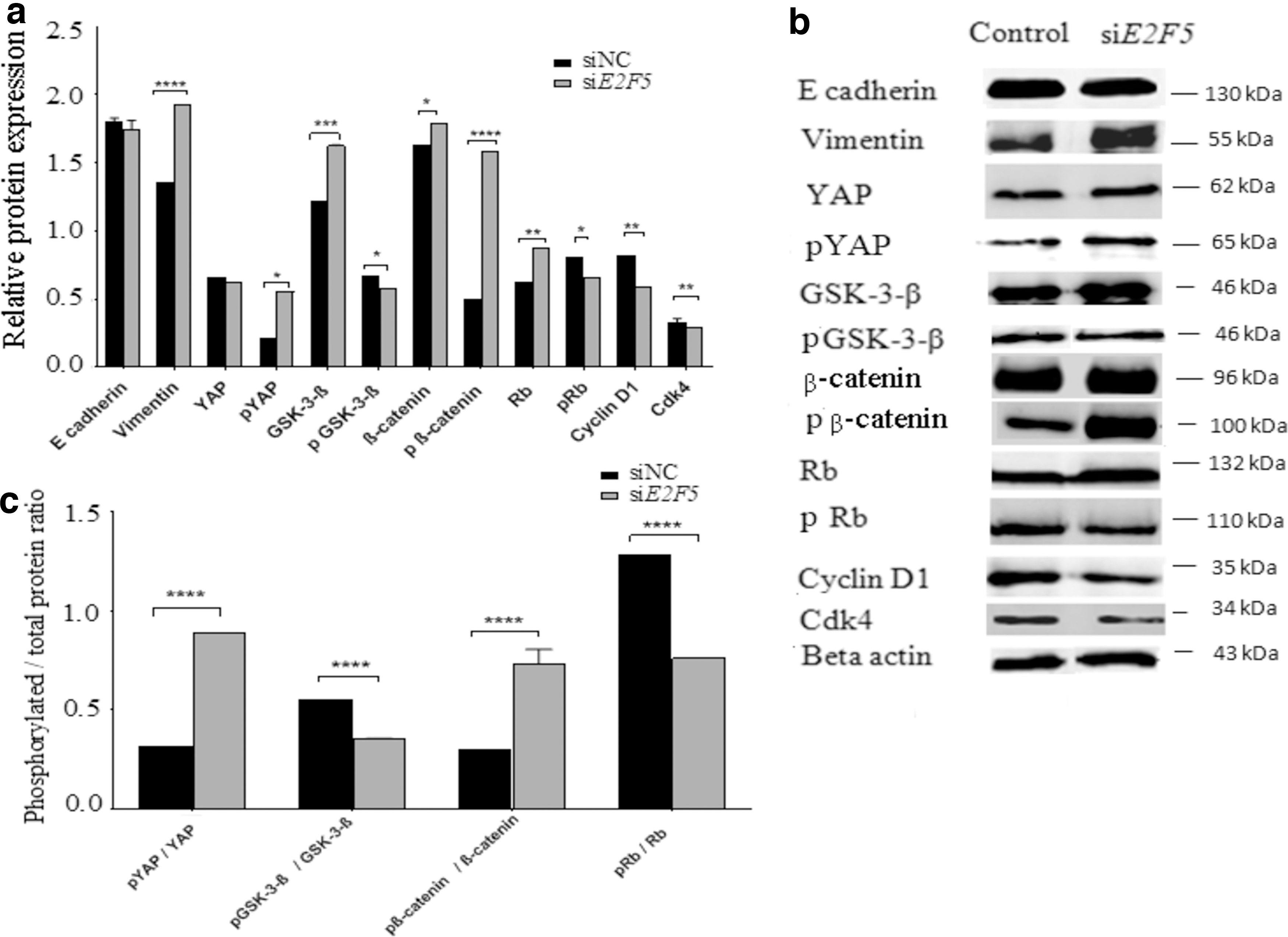
Regulatory effects of E2F5 on the expression of proteins involved in EMT, Hippo, Wnt-β-catenin, apoptotic, and retinoblastoma pathways by western blotting in MCAS cells.
Consistent with the western blotting results, Pearson's correlation showed a negative correlation between E2F5 and caspase 9 (p = 6.2e−9, r = −0.25); while a positive correlation was observed between E2F5 and YAP1 (p = 2.2e−9, r = 0.26); Cyclin D1 (p = 8.9e−16, r = 0.35); and cdk4 (p < 0.05, r = 0.4) and β-catenin (p = 2.2e−6, r = 0.21) (Fig. 4).
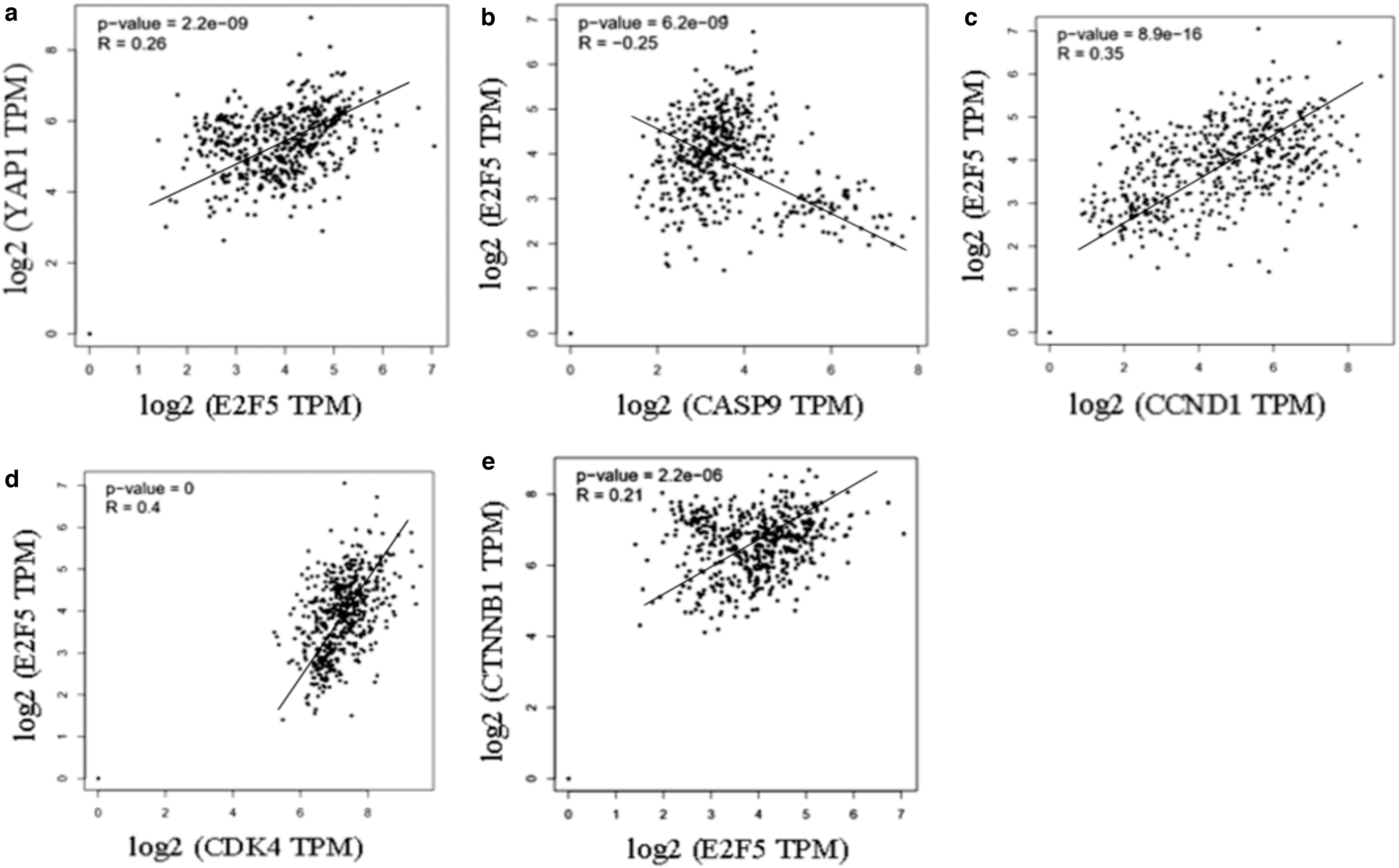
Pearson's correlation between different gene expressions using samples from GEPIA database.
Discussion
In this study, we examined the role and associated signaling mechanisms of the E2F5 gene in EOC tumorigenesis and invasion. E2F5 expression was higher in ovarian cancer cell lines compared to normal ovarian cell line (HOSE 6-3), with the highest expression in A2780-cps and A2780-cpr, followed by OVSAHO and MCAS ovarian cancer cell lines (Fig. 1a). In our hands, the most suitable model for the knockdown study would be the cell lines displaying a moderate expression of the desired gene. Hence, we selected MCAS and OVSAHO as it showed moderate expression of E2F5, unlike A2780-cps and A2780-cpr, which overexpressed E2F5. Cells highly expressing genes usually have active promoters allowing a high turnover of the related messengers and therefore minimizing the effect of siRNA knockdown.
Furthermore, bioinformatics analysis of human samples revealed similar results, showing upregulation of E2F5 expression in human ovarian cancer samples when compared to normal tissues (Fig. 1b). Our results were consistent with previous findings, with E2F5 overexpression in ovarian cancer samples when compared to healthy or benign ovarian tissues (Kothandaraman et al., 2010; Zhou et al., 2019).
To gain further insight into the functional role of E2F5 on cell survival, colony formation ability, and invasiveness, we transfected MCAS and OVSAHO with E2F5-siRNA (Fig. 1c and d) and assessed the proliferation, colony formation, and invasiveness of the cells. We observed that the transient knockdown of E2F5 significantly inhibited cell growth (Fig. 2a), decreased the anchorage-independent growth (Fig. 2b), and the invasive ability of cells (Fig. 2c). Consistent with our findings, E2F5 knockdown inhibited cell proliferation, migration, and invasion in SKOV3 ovarian cancer cell lines (Tian et al., 2016) hepatocellular carcinoma (Jiang, 2011), and prostate cancer (Zheng et al., 2017). Furthermore, previous findings determined the impact of E2F5 repression on cell proliferation using flow cytometry, which revealed an accumulation of cells in the G0/G1 phase and reduction in the number of cells in the S-phase of the cell cycle (Jiang, 2011).
Cell growth, proliferation, and apoptosis are regulated by the Hippo pathway, a kinase signaling cascade (Bae and Luo, 2018). The downstream effector (YAP) in this pathway is a crucial ovarian cancer oncogene promoting increased cell proliferation, invasion, and anchorage-independent growth (Xia et al., 2014). Our western blotting results implied a higher level of inactive, phosphorylated cytoplasmic YAP in E2F5 siRNA treated cells, unable to transcribe cell growth and antiapoptotic genes, while the levels of active unphosphorylated β-catenin; a protein involved in Wnt-β-catenin pathway was lower. In contrast, the level of total GSK-3-β was higher, and phosphorylated form was reduced in E2F5 siRNA treated cells compared to control, suggesting increased levels of active GSK-3-β, which can inactivate β-catenin by phosphorylation (Fig. 3a-c).
During the G1 phase of the cell cycle, Rb is phosphorylated by cyclin-dependent kinases (cdks), causing it to dissociate from E2F transcription factors resulting in the transcription of genes responsible for promoting entry into S-phase (Dyson, 1998). In this study, E2F5 repression reduced cyclin D1, cdk4, and phosphorylated Rb expression inhibiting progression into the G1/S phase of the cell cycle (Fig. 3a-c). Similarly, in a previous study, E2F5 knockdown resulted in G1 cell cycle arrest of prostate cancer cells (Zheng et al., 2017), and regulated the expression of cdks in neuroblastoma cells (Liu et al., 2019) with a downregulation of cdk4 in E2F5 silenced prostate cancer cells (Li et al., 2018).
Consistent with the western blotting results, Pearson's correlation showed a positive correlation between E2F5 and YAP1; Cyclin D1; cdk4; and β-catenin, while a negative correlation between E2F5 and caspase 9 was observed (Fig. 4).
We had previously performed ChIP analysis on MCAS and OVSAHO cell lines to identify the genes regulated by E2F5, a transcription factor overexpressed during the early stages of EOC. This analysis revealed several downstream genes regulated by E2F5, which themselves might have potential roles in EOC pathogenesis (unpublished data). Among the downstream genes regulated by E2F5, FAT4, a key component of the Hippo pathway was identified. Knocking down FAT4 was found to upregulate the expression of E2F5 (Malgundkar et al., 2020), suggesting the tight relationship between these two genes. Since YAP is a downstream effector of the Hippo pathway, we speculated that E2F5 regulates the expression of YAP, via FAT4, and several other proteins known to interact with YAP, such as β-catenin. Hence, we performed western blotting in E2F5 knocked cells to suggest the putative mechanism involved in EOC pathogenesis.
A suggested mechanism through which E2F5 regulates cell growth and invasion, via the regulation of components involved in Hippo, Wnt-β-catenin, and the cell cycle pathways is proposed based on our western blotting results (Fig. 5). E2F5 modulates YAP activity through the regulation of its phosphorylation, as demonstrated by the accumulation of inactive and phosphorylated YAP upon E2F5 repression. Hence, E2F5 upregulates YAP nuclear localization in ovarian cancer, which in turn promotes the nuclear localization of nonphosphorylated active β-catenin. Furthermore, nuclear translocation of β-catenin leads to transcription of cyclin D1, c-myc, and mesenchymal markers such as twist, and vimentin, promoting cell growth and invasion.
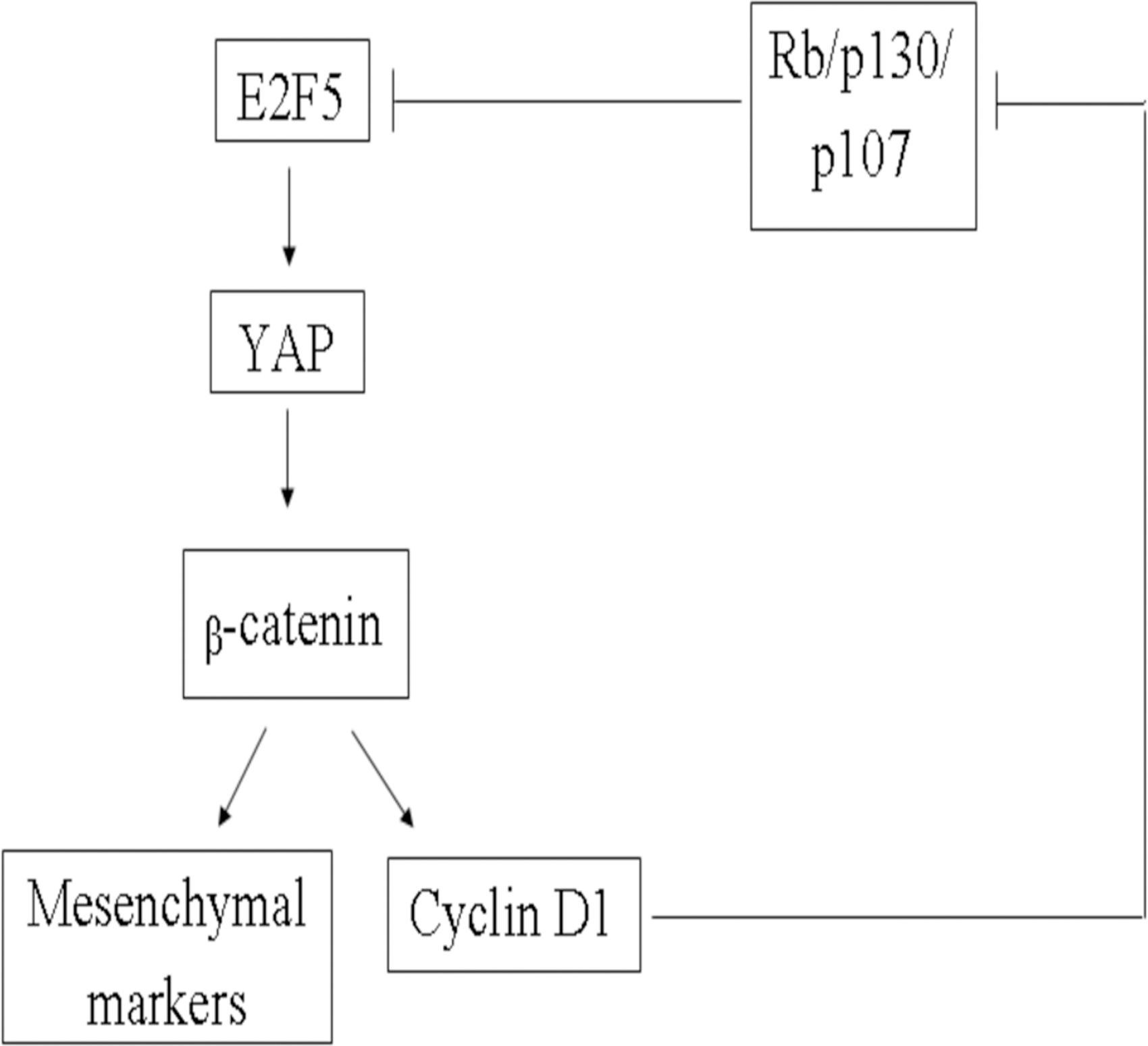
A suggested mechanism of growth and cell cycle regulation by E2F5 based on western blot-obtained data. Inhibition of E2F5 regulates YAP1 and β-catenin, thereby inhibiting cell cycle progression via the suggested pathway.
Furthermore, a transcriptional target of β-catenin, cyclin D1, is known to phosphorylate the retinoblastoma protein family (Rb, p130, and p107), resulting in its inability to associate with E2F5, thus increasing the level of active E2F5 protein.
Our results demonstrated the involvement of E2F5 in proliferation and invasion via the regulation of YAP and β-catenin levels. This study contributes to a better understanding of the molecular mechanism involving E2F5 in EOC. In conclusion, although our in vitro experiments and analysis of expression data from publicly available databases are supportive of our hypothesis, additional in vivo experiments need to be conducted for confirmation.
Footnotes
Acknowledgment
The authors would like to thank Professor Gunnar Norstedt for proofreading the article.
Availability of Data and Materials
The data sets generated during and/or analyzed during this study are included in this published article, and other datasets will be available from the corresponding author on reasonable request.
Authors' Contributions
M.A.M. conceived the idea of initiating the work. Y.T. designed the project and the experimental setup. S.H.M. performed the experiments, interpreted the data, and wrote the article under the supervision of Y.T.. A.O., I.B., M.A.K., and R.L. and contributed to constructive critiques and discussion related to the clinical section of the project. They also contributed to the article reviewing. All the authors approved the final version of this article.
Ethics Approval and Consent to Participate
The cell lines used in this study do not require ethics approval for their use.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by His Majesty Trust Fund (SR/MED/BIOC/14/01) grant in SQU.