
Abstract
Aims:
Many studies and researchers have reported on the genetic association between lipoprotein lipase (LPL) gene polymorphisms and myocardial infarction (MI). The results, however, have been inconclusive. Therefore, we assessed the relationship of LPL gene polymorphisms and MI risk by performing a meta-analysis.
Methods:
Literature was retrieved through PubMed, Web of Science, the Cochrane Library, Chinese National Knowledge Infrastructure (CNKI), and Embase databases. Pooled odds ratios (ORs) with 95% confidence intervals (CIs) were used to evaluate the genetic associations between LPL gene polymorphisms and MI risk. A total of nine studies, with 10 individual groups, comprising 2785 cases and 4317 controls were used for this meta-analysis.
Results:
The allelic (p = 0.0003, OR [95% CI] = 0.86 [0.79-0.93]) and dominant models (p = 0.001, OR [95% CI] = 0.83 [0.73-0.93]), but not the recessive model (p > 0.05) for the LPL gene showed that the HindIII variant significantly decreased the risk of MI. In addition, the allelic model (p = 0.04, OR [95% CI] = 0.71 [0.50-0.99]) for the S447X variant showed a significant decrease in the risk of MI. No association was observed between the PvuII variant and MI (p > 0.05). A subgroup analysis based on ethnicity revealed that all of the genetic models (allelic model: p < 0.00001, OR [95% CI] = 0.62 [0.51-0.77]; dominant model: p = 0.003, OR [95% CI] = 0.66 [0.50-0.87]; recessive model (p = 0.02, OR [95% CI] = 0.47 [0.25-0.88]) found an association of the HindIII polymorphism with MI in the Asian, but not in the Caucasian population (p > 0.05). Under the dominant model the HindIII SNP was also shown to be associated with MI risk in the Caucasian population (p = 0.03, OR [95% CI] = 0.87 [0.76-0.99]). In addition, the allelic (p = 0.02, OR [95% CI] = 0.75 [0.59-0.95]) and dominant models (p = 0.02, OR [95% CI] = 0.51 [0.29-0.90]) for S447X demonstrated a significantly decreased MI risk in the Caucasian, but not in the Asian population (p > 0.05).
Conclusions:
LPL HindIII and S447X polymorphisms, but not PvuII might be the protective factors for MI. To confirm these results, case-control studies with larger numbers of subjects need to be conducted.
Introduction
Lipid metabolism disorders and arteriosclerosis, hypertension and obesity, and the occurrence of cardiovascular (CVD) and cerebrovascular diseases are major problems around the world (Paapstel et al., 2016; Holmes et al., 2018). Lipoprotein lipase (LPL) is a key enzyme in lipid metabolism. And its deficiency can lead to a degradation disorder of triglycerides (TGs) in chylomicrons (CM) and very low-density lipoprotein (VLDL) particles, resulting in lipid metabolism disorder (Olivecrona, 2016). LPL defects lead to hypertriglyceridemia and hyperlipidemia, which are associated with early onset atherosclerosis and CVD, including coronary artery disease (CAD) and myocardial infarction (MI) (Kuller, 2007; Majumdar et al., 2010; Shia et al., 2011). LPL plays a particularly important role in the occurrence of MI.
The LPL gene is located at chromosome 8p22 (Yang and Guo, 2009; Klannemark et al., 2010). Multiple polymorphisms, including PvuII (rs285), HindIII (rs320), and S447X (rs328), in the LPL gene have been investigated for their genetic associations with CAD risk (Duman et al., 2004; Sagoo et al., 2008; Rebhi et al., 2012). The HindIII (rs320) polymorphism encodes a T to G transversion at position +495 (Hemimi et al., 2009), which abolishes a restriction site for HindIII. The HindIII H+ allele may lead to high TG levels and low high-density lipoprotein cholesterol (HDL-C) levels in healthy individuals who carry this allele (Heizmann et al., 1991; Gerdes et al., 1995). In addition, the H+H+ genotype of HindIII has been suggested to elevate TG levels and contribute to the risk of MI in Russian and South Indian populations (Malygina et al., 2001; Tanguturi et al., 2013). However, the genetic relationship of this polymorphism and MI has not been shown to be consistent in other populations (Peacock et al., 1992; Tarek et al., 2011). The PvuII (rs285) polymorphism encodes a replacement of C to T in intron 6 (Fisher et al., 1987); and the PvuII (rs285) P+ allele has been shown to influence the TG and HDL-C levels (Chamberlain et al., 1989; Chung et al., 1999), which may indicate that the PvuII (rs285) polymorphism is a susceptibility factor for CVD and cerebrovascular diseases.
The LPL S447X (rs328) polymorphism truncates the LPL protein loss of the two C-terminal amino acids. This polymorphism was shown to be associated with increased LPL protein secretion and plasma post-heparin activity, as well as lower TG levels (Stocks et al., 1992; Lee et al., 2004). This polymorphism has been shown to have a protective effect against MI. The X allele was shown to have a protective effect in the Chinese (Zou et al., 2008) and Egyptian populations (Tarek et al., 2011). This effect, however, was not shown in the Swedish (Peacock et al., 1992) and Costa Rican populations (Yang et al., 2004).
Considering the inconsistent and inconclusive results of the individual studies in this area, we performed this meta-analysis by pooling all eligible studies to investigate the correlation between the LPL HindIII (rs320), PvuII (rs285), and S447X (rs328) variants, and the risk of suffering from MI.
Materials and Methods
Literature search
This meta-analysis followed the Cochrane collaboration definition and PRISMA 2009 guidelines for meta-analyses and systematic review (Boccia, 2009). Publications were searched on PubMed, Embase, Chinese National Knowledge Infrastructure (CNKI), and Cochrane Collaboration Library database. The key words were as follows: “lipoprotein lipase” or “LPL” and “polymorphism” or “variant” or “Single nucleotide polymorphism” or “SNP” and “myocardial infarction” or “MI.” No language and published year were limited. References in relevant publications were also examined manually.
Selection criteria
Inclusion criteria: (1) case-control designed; (2) referred to the association between the LPL polymorphisms (HindIII, S477X, and PvuII) and MI risk; (3) the data of the genotypes were available; (4) the distribution of genotypes in the control group were in Hardy-Weinberg equilibrium (HWE).
Exclusion criteria: (1) abstracts, meeting reports, letters, correspondence, reviews, meta-analysis, and animal studies; (2) duplicated studies; (3) insufficient data for the genotypes; (4) did not refer to the association between the LPL polymorphisms (HindIII, S477X, and PvuII)) and MI risk; (5) the control group did not confirm to HWE.
Data extraction
Two independent authors screened and extracted the data. Disagreements concerning study selection were discussed and resolved by the third investigator. Information from each study was extracted as follows: author's name, year of publication, percentage of males, mean ages, body mass index, systolic blood pressure, diastolic blood pressure, low-density lipoprotein cholesterol (LDL-C), HDL-C, TG, and total cholesterol in both cases and controls, percentage of individuals who smoke and/or have hypertension, and number of cases and controls. All of the information is summarized in Table 1.
The Characters of Included Studies
BMI, body mass index; DBP, diastolic blood pressure; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; NA, not available; NOS, Newcastle-Ottawa scale; SBP, systolic blood pressure; TC, total cholesterol; TG, triglyceride.
Quality assessment
The Newcastle-Ottawa scale (NOS) was applied in the quality assessment (Han et al., 2017). The quality assessment comprised three parameters of quality, including selection, comparability, and exposure assessment. The NOS uses a “star” rating system to judge quality. A study with a total of six stars or higher was included in this analysis.
Statistical analyses
The statistical analyses were performed using STATA 12.0 software (StataCorp., College Station, TX) and Revman 5 (Cochrane Collaboration, London, UK). Genetic associations between the genetic models (dominant, recessive, and allelic models) of LPL HindIII, PvuII, and S477X, and MI risk were evaluated by odds ratios (ORs) with 95% confidence intervals (CIs). The summary OR was determined by the Z test. The heterogeneity among studies was evaluated by Cochrane's Q-statistic and I2 statistics. The random model was used in case of significant heterogeneity (I2 > 50%, or p < 0.05). Otherwise, the fixed model was utilized. Subgroup analyses, based on ethnicity (Asians and Caucasians), were applied to detect sources of heterogeneity. The stability of the individual studies was evaluated by sensitivity analysis. In addition, a funnel plot based on Egger's test and Begg's tests was used to assess potential publication bias. A p < 0.05 indicates significant publication bias.
Results
Study characteristics
As shown in Figure 1, 347 publications were initially identified and screened. These studies were then carefully analyzed according to the selection criteria. Finally, nine case-control studies were selected for this meta-analysis (Peacock et al., 1992; Holmer et al., 2000; Malygina et al., 2001; Yang et al., 2004; Baum et al., 2006; Zou et al., 2008; Tarek et al., 2011; Tanguturi et al., 2013; Bahrami, 2015). The study conducted by Holmer et al. (2000) included two separate groups, which we treated as two individual studies. Therefore, a total of 10 studies, including 2785 cases and 4317 controls, were utilized for this meta-analysis. Among the included studies, five were conducted among Asian populations (Baum et al., 2006; Zou et al., 2008; Tarek et al., 2011; Tanguturi et al., 2013; Bahrami, 2015), and five among Caucasian populations (Peacock et al., 1992; Holmer et al., 2000; Malygina et al., 2001; Yang et al., 2004). For the HindIII polymorphism, eight studies with 2468 cases and 3914 controls were available (Peacock et al., 1992; Holmer et al., 2000; Malygina et al., 2001; Yang et al., 2004; Baum et al., 2006; Tarek et al., 2011; Tanguturi et al., 2013; Bahrami, 2015); for the Ser447X polymorphism, five studies with 1925 cases and 1917 controls were available (Peacock et al., 1992; Yang et al., 2004; Baum et al., 2006; Zou et al., 2008; Tarek et al., 2011); and for the PvuII polymorphism, three studies with 790 cases and 816 controls were available (Peacock et al., 1992; Yang et al., 2004; Bahrami, 2015). The distributions of genotypes of LPL HindIII, Ser447X, and PvuII polymorphisms were in HWE for the control groups. The detailed characteristics of the eligible publications are shown in Table 1. The NOS quality assessment is provided in Supplementary Table S1.
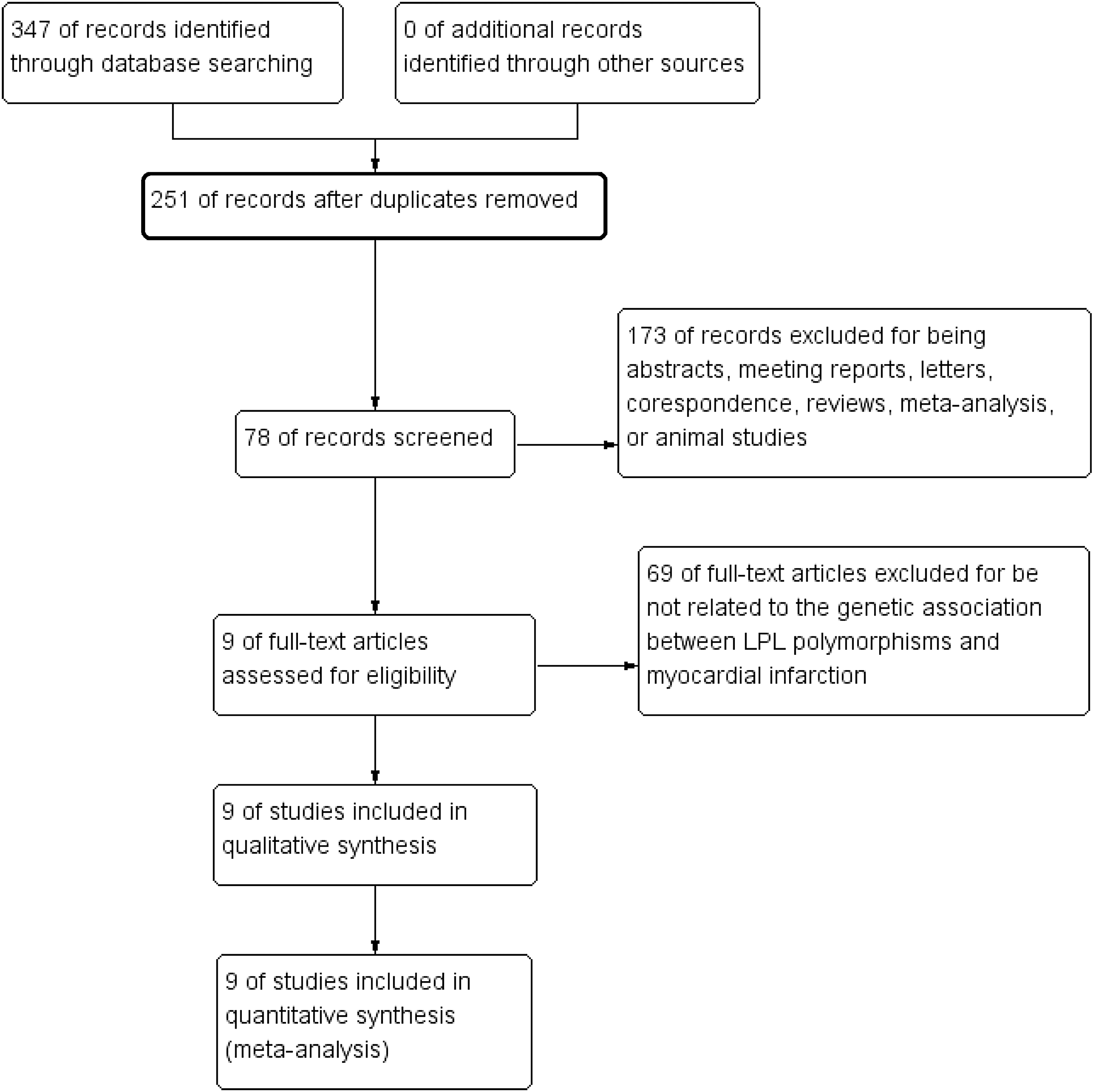
PRISMA flowchart of studies inclusion and exclusion.
Results of meta-analysis
The overall data showed that the allelic (p = 0.0003, OR [95% CI] = 0.86 [0.79-0.93]) and dominant models (p = 0.001, OR [95% CI] = 0.83 [0.73-0.93]), but not the recessive model (p > 0.05) of HindIII significantly decreased the risk of MI (Fig. 2 and Table 2). In addition, the allelic model (p = 0.04, OR [95% CI] = 0.71 [0.50-0.99]) of S447X can significantly decrease the risk of MI (Fig. 3 and Table 2). A random-effects model was used for the significant heterogeneity among the studies (except for the recessive model of S447X, p < 0.05, or I2% > 50%). Furthermore, subgroup analyses based on ethnicity indicated that the allelic (p < 0.00001, OR [95% CI] = 0.62 [0.51-0.77]) and recessive models (p = 0.02, OR [95% CI] = 0.47 [0.25-0.88]) of HindIII can significantly decrease the risk of MI in the Asian, but not in the Caucasian populations (p > 0.05). Whereas the dominant model of HindIII was associated with MI risk both in Asian (p = 0.003, OR [95% CI] = 0.66 [0.50-0.87]) and Caucasian populations (p = 0.03, OR [95% CI] = 0.87 [0.76-0.99]). For S447X, the allelic (p = 0.02, OR [95% CI] = 0.75 [0.59-0.95]) and dominant models (p = 0.02, OR [95% CI] = 0.51 [0.29-0.90]) were shown to be associated with MI risk in the Caucasian, but not in the Asian population (p > 0.05). No association was detected between the genetic models of the PvuII polymorphism and MI risk (p > 0.05) (Fig. 4 and Table 2).
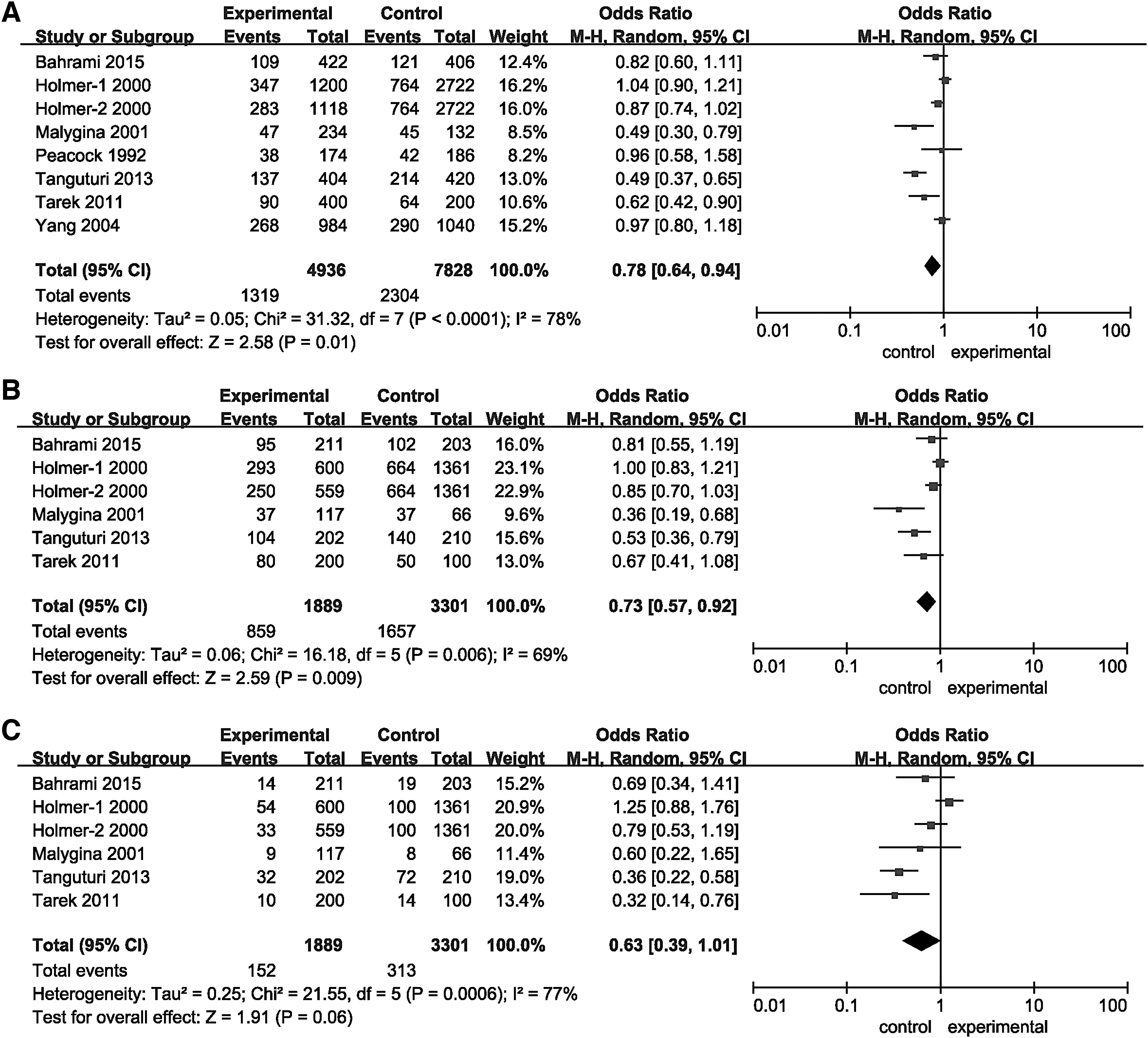
Forest plots of ORs for the association between LPL HindIII and MI.

Forest plots of ORs for the association between LPL S447X and MI.
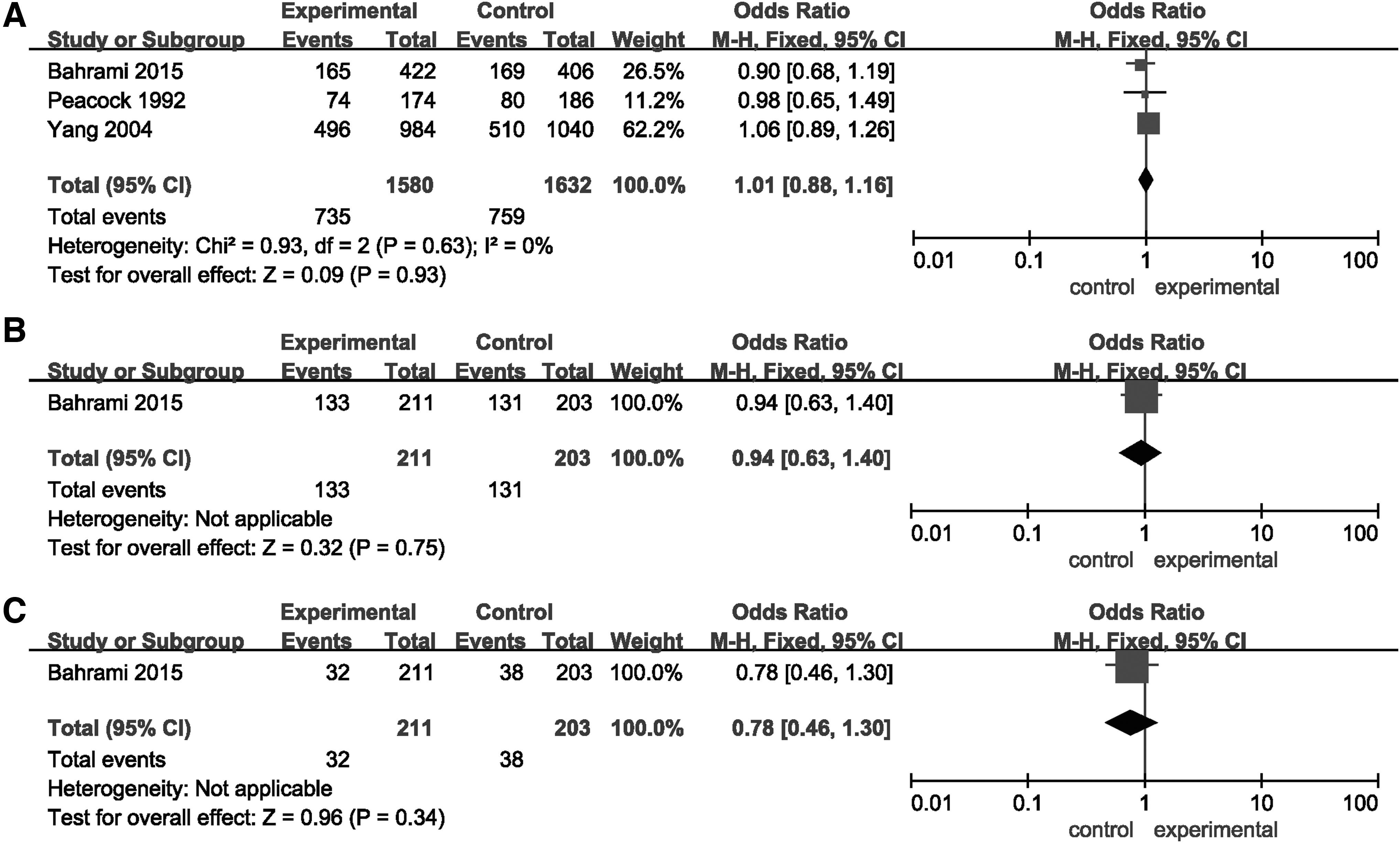
Forest plots of ORs for the association between LPL PvuII and MI.
The Results of Meta-Analysis for Lipoprotein Lipase HindIII, S447X, and PvuII, and Myocardial Infarction
R, random model; F, fixed model; —, not available
CI, confidence interval; OR, odds ratio.
Heterogeneity and sensitivity analyses
The heterogeneity within each study in each comparison is shown in Table 2. The results indicate that there was no significant alteration for the pooled OR after deleting one study at a time (Fig. 5), which supports the stability of the individual studies included in this meta-analysis.
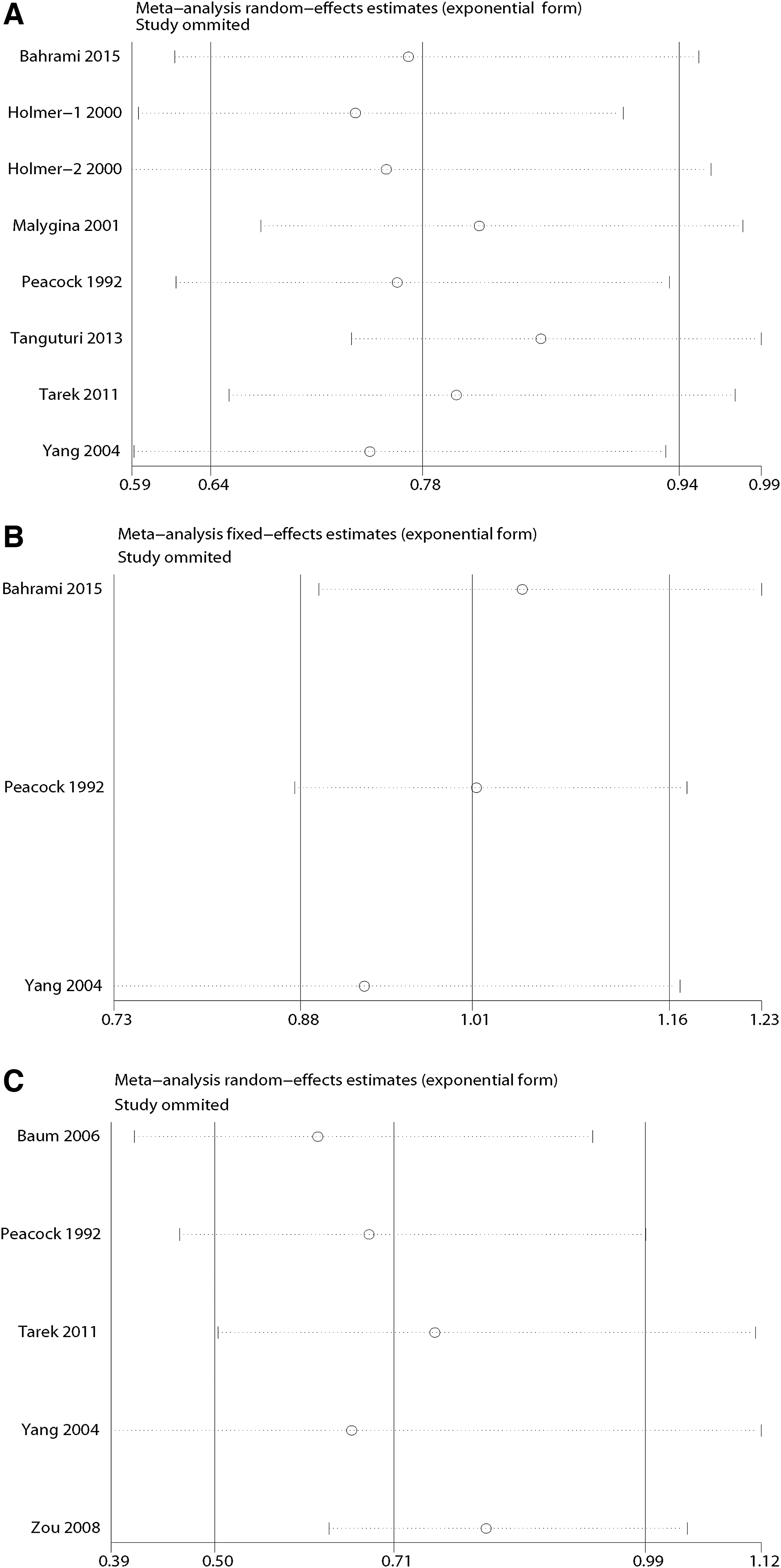
Sensitivity analyses between LPL HindIII, S447X, and PvuII, and MI.
Publication bias
Begg's funnel plot and Egger's test were used to evaluate the publication bias of this meta-analysis. The funnel plot shapes were symmetrical for alleles of the LPL HindIII, PvuII, and S447X polymorphisms. The results of Egger's regression test are also presented under the allelic models (Fig. 6). Therefore, there was no publication bias among the included studies.
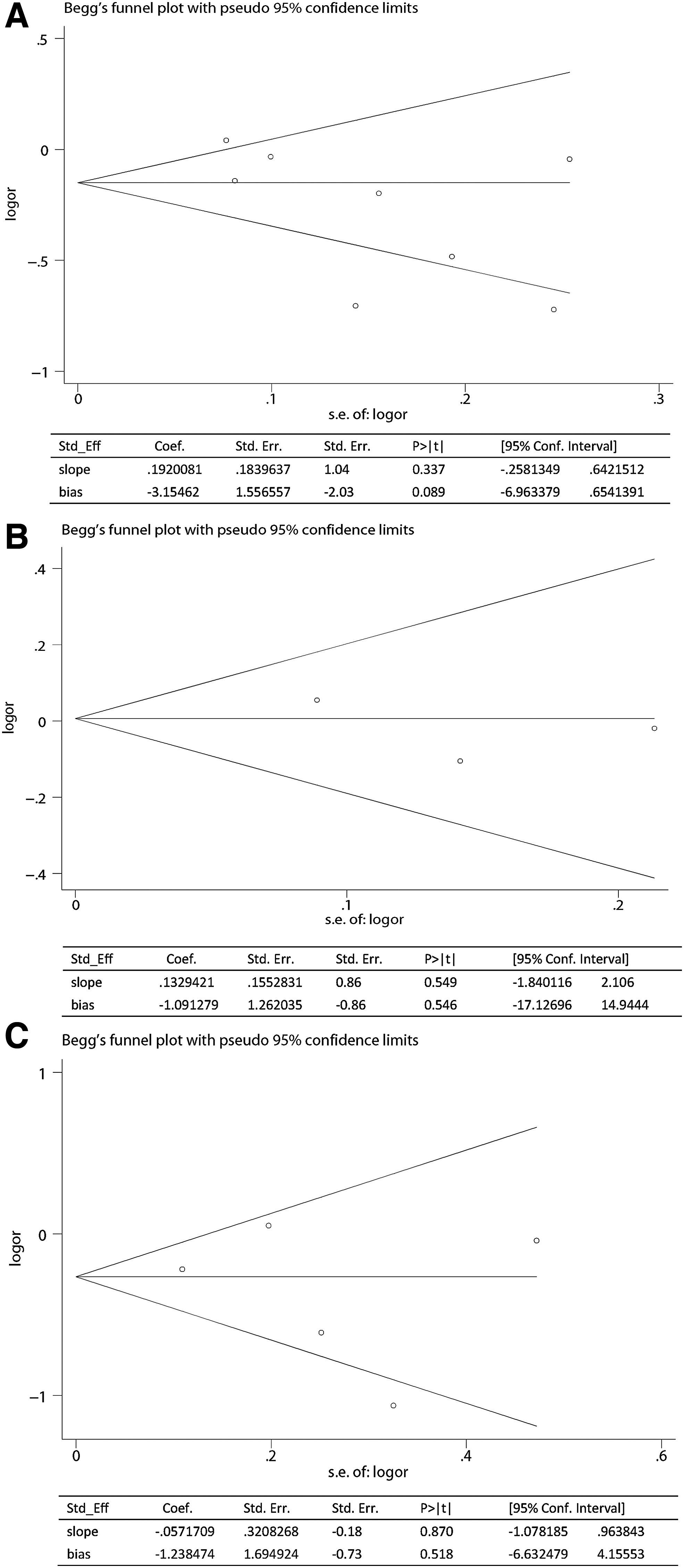
Publication bias of literatures for allelic model of LPL HindIII, S447X, and PvuII were tested by Begg's funnel plot and Egger's test.
Discussion
LPL is a glycoprotein composed of 448 amino acids with a molecular weight of 65 kDa, which is primarily synthesized and secreted by fat cells, myocardium, skeletal muscle, and other extracellular parenchymal cells (Goldberg and Merkel, 2001). LPL can hydrolyze TGs in the blood. The glycerol and fatty acids generated by LPL can be oxidized and decomposed, which plays a key role in the catabolism of CM and VLDL (Hata et al., 1990). LPL defects lead to hypertriglyceridemia and hyperlipidemia, which contributes to a myriad of pathogenic conditions, including MI (Watson et al., 2010; Surendran et al., 2012).
The relationships of the LPL HindIII polymorphism and plasma TG, TC, HDL-C levels have been investigated with inconclusive results (Merkel et al., 2002; Ma et al., 2003). And, the HindIII H+ allele has been shown to be associated with the alteration of TG, HDL-C, and severity of atherosclerosis, as well as the risk of CAD and MI (Ye et al., 1996). An increased risk of MI was detected for the HindIII H+ allele in the study conducted by Malygina et al. (2001). Similar results were observed in Egyptian and Indian populations (Tarek et al., 2011; Tanguturi et al., 2013). Whereas other studies conducted by Peacock et al. (1992), Holmer et al. (2000), and Bahrami (2015) reported no association between the LPL HindIII polymorphism and MI risk both in Asian and Caucasian populations. In this study, we detected the genetic association between the LPL Hind III polymorphism and MI susceptibility by using a meta-analysis. The allelic and dominant models of the LPL HindIII polymorphism were found to significantly decrease the MI risk, indicating protective effects of these genetic models of LPL HindIII on MI. Furthermore, the genetic association between the allelic and recessive models of LPL HindIII was detected in the Asian group, but not in the Caucasian group. Different gene backgrounds in Asians and Caucasians may lead to this inconsistency.
S447X (rs328) located in exon 9 of LPL gene. It has been shown to encode a Ser to Gly switch at the C terminus of the LPL protein. The alteration may shorten the LPL protein and affect its receptor affinity (Mc-Gladdery et al., 2001). The function of LPL in hydrolyzing TGs might be enhanced by the LPL S447X polymorphism, which may hinder the development of atherosclerosis. Lower serum LDL was detected in patients carrying the S447X mutation, which indicates a role of S447X in plasma lipoprotein metabolism and the development of CAD (Stocks et al., 1992). Therefore, the LPL S447X may be a susceptibility factor for CVD and cerebrovascular diseases (Gigek et al., 2007). According to our results, the allelic and dominant models the LPL S447X polymorphism was significantly associated with decreased risk of MI compared with the control group, indicating a protective effect for this polymorphism on MI. However, this protective effect was only observed in the Caucasian subgroup. These inconsistencies based on ethnicity may implicate the influence of the genetic background in the pathogenesis of MI. To figure out the role of this polymorphism with regard to the pathogenesis of MI, additional mechanistic studies are needed.
Limitations should be considered. First, the number of included studies and subjects was relatively small, especially for the PvuII polymorphism and in the Asian subgroup, which may partly influence the effects of this polymorphism on the genetic risk of MI. To further identify the results in this analysis, more studies with larger numbers of subjects are necessary. Second, both genetic and environmental factors are likely involved in the development of MI. However, we failed to detect associations between environmental factors and MI risk due to lack of sufficient data. Third, the ethnic distribution of the included studies was primarily in Asians and Caucasians. Racial bias may exist, and the conclusions may not be applicable to other races. In addition, some genetic models displayed high heterogeneity, even though subgroup analyses were performed to detect the sources of heterogeneity.
In conclusion, this meta-analysis suggested, the LPL HindIII and S447X polymorphisms might be protective factors for Asians against MI. In addition, the LPL HindIII polymorphism might be protective factor for Caucasians against MI. However, there was no support for a protective or causative role for the PvuII polymorphism associated with MI.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Foundation of the Education Department of Hunan (18B536), the Hunan Key Laboratory of the Research and Development of Novel Pharmaceutical Preparations (2016TP1029), and Clinical medicine professional innovation and entrepreneurship education base (Xiangjiaotong-[2018]-380).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.