
Abstract
Objective:
Expression of circular RNAs (circRNAs) in the peripheral blood of individuals with latent autoimmune diabetes in adults (LADA) and type 2 diabetes mellitus (T2DM) were quantified to identify dysregulated circRNAs compared with control individuals.
Methods:
circRNAs were obtained from the peripheral blood serum of 12 healthy adults and 12 individuals with LADA and 12 type 2 diabetics. The circRNA expression profiles were analyzed by high-throughput RNA sequencing. The most highly dysregulated circular RNAs were validated by quantitative real-time polymerase chain reaction. A circular RNA-microRNA (miRNA) network diagram predicted the interactions of circular RNAs, miRNAs, and coding genes.
Results:
A total of 2334 differentially expressed circRNAs were detected among the three groups, with 277 circRNAs in the Group DM versus Group NG; 992 circRNAs in the Group LADA versus Group NG and 1065 circRNAs in the Group DM versus Group LADA. Six circRNAs were identified as the most distinctive differentially expressed targets (p < 0.05). The proposed molecular functions of these differentially expressed circRNAS included the tumor necrosis factor signaling pathway, the FoxO signaling pathway, cellular senescence, and long-term potentiation (all false discovery rate p < 0.05) which may contribute to T2DM and LADA.
Conclusion:
circRNAs are aberrantly expressed in the peripheral blood of patients with T2DM and LADA and may interact with miRNA and circRNA-derived peptides in the development of diabetes. Further investigations may illustrate the partial pathogenesis of diabetes mellitus.
Clinical Trial Registration number: ChiCTR1900020644.
Introduction
Diabetes mellitus is a metabolic disease characterized by chronic hyperglycemia that occurs due to a combination of genetic and epigenetic risk factors (Benincasa et al., 2020; Napoli et al., 2020; Schiano et al., 2020; Lei et al., 2021; Pepin et al., 2021; Wang et al., 2021). However, the etiology and pathogenesis of diabetes has not yet been fully clarified (Chamberlain et al., 2016). So far, comprehensive diabetes classification is based on an understanding of the clinical manifestations, pathophysiology, and etiology of diabetes mellitus. Types of diabetes mellitus include type 1 and type 2 diabetes, gestational diabetes, genetic defects in β cell function and other special types (Nauck et al., 2021). Individuals that experience a slow start-up and inconspicuous early clinical manifestations could experience a long or short phase of insulin-free treatment.
Early clinical manifestations in these individuals appear as type 2 diabetes mellitus (T2DM), yet are characterized by islet β cell autoimmune damage seen in type 1 diabetes. This is considered as latent autoimmune diabetes in adults (LADA) (N and Subhakumari, 2016). The diagnostic criteria for LADA from the Immunology of Diabetes Society are: (1) adult age of onset >30 years, (2) positive autoantibody to any islet cell, and (3) absence of insulin requirement for at least 6 months after diagnosis (Fourlanos et al., 2005).
Circular RNA (circRNA) is a kind of noncoding RNA, which is derived from exonic circRNAs, intronic circRNAs, or a combination of the two known as exon-intron circRNAs. Some circRNAs can be used as “microRNA (miRNA) sponges,” playing a role in posttranscriptional regulation by engaging in competitive combination with miRNA (Lei et al., 2020). circRNA has a circular structure, is more stable in vivo, and is more abundant in tissues, which renders them to be potential diagnostic molecular markers for various diseases (Zaiou, 2019). There are many studies showing that circRNAs are closely related to the occurrence and development of diabetes.
A study investigated the expression profiles of circRNAs in the peripheral blood of patients with T2DM and validated the utility of hsa_circ_0054633 as a diagnostic biomarker for prediabetes and T2DM (Zhao et al., 2017). A retrospective case-control study found that hsa_circRNA_0054633 was highly expressed in the blood during the second and third trimesters and was highly correlated with GHBA1 and GHBA1c levels in maternal blood samples at various stages of the GDM group (Wu et al., 2019). No research on the relationship between LADA and circRNA is currently available.
In this study, we investigated the characteristic expression of circRNAs in the peripheral blood of individuals with LADA and T2DM. The circRNA expression profile in individuals with LADA and T2DM was systematically explored by comparison with matched control participants through RNA sequencing. Our findings provide important insights into the roles of circRNAs in individuals with diabetes.
Materials and Methods
Study design
This study was approved and supervised by the Ethics Committee of The Second Hospital of Jilin University, Changchun, China (No. [2018]137). All experiments were performed in accordance with the Code of Ethics of the World Medical Association. All participants provided written, informed consent. This study conformed to the ethical guidelines of the 1975 Declaration of Helsinki, is registered with the Chinese Clinical Trial Registry, and was closed to accrual.
Study participants were categorized into three groups, including LADA participants (Group LADA), T2DM participants (Group DM), and healthy participants (Group NG). Experimental group participants were enrolled from the Department of Endocrinology outpatients and inpatients, The Second Hospital of Jilin University from February to May of 2019 and control group participants were included from the population who underwent physical examination in the hospital during the same period. Each participant was tested using the American Diabetes Association diagnostic criteria to determine whether an individual was healthy or had diabetes. Table 1 shows the characteristics of the Group LADA, Group DM, and Group NG.
The Clinical Characteristics of the Study Population
p < 0.05; **p < 0.01; ***p < 0.001; compared with GROUP NG.
p < 0.05; ###p < 0.001; compared with GROUP DM.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; BMI, body mass index; GLU, glucose; HbA1c, hemoglobin A1c; HDL, high-density lipoprotein cholesterol; LADA, latent autoimmune diabetes in adults; TC, total cholesterol; TG, triglyceride.
LADA definition and whole blood sample collection
The LADA group contained participants who were 30-55 years at the time of diabetes diagnosis, who did not require insulin treatment for at least 6 months after diagnosis, and who had glutamic acid decarboxylase autoantibody or other islet cell autoantibody (islet cell antibody-IgG, zinc transporter-8 antibody, insulin autoantibody, and protein tyrosine phosphatase-like IA-2 autoantibody) positivity. Exclusion criteria included other special types of diabetes or gestational diabetes, malignancies, serious liver and kidney dysfunction, any other acute or chronic inflammatory diseases, untreated essential hypertension (≥160/100 mmHg), any endocrine diseases other than diabetes, any acute and chronic diabetic complications, and pregnant or lactating women. Details of antibody positivity in the LADA group were provided in Supplementary Table S1.
RNA library sequencing and subsequent bioinformatics analysis
Venous blood samples were collected from three groups. Total RNA of each specimen was quantified, and quality assurance was confirmed using NanoDrop ND-1000 spectrophotometer (NanoDrop, Wilmington, DE, USA). Standard denaturing agarose gel electrophoresis was performed to assess RNA integrity. For each sample we pooled random 3 RNA samples in each group, to limit variation due to differences in vector injection. Cloud-Seq Biotech (Shanghai, China) performed transcriptome high-throughput sequencing and subsequent bioinformatics analysis.
Briefly, total RNA quality was assessed using agarose gel electrophoresis and quantified using a NanoDrop spectrophotometer (NanoDrop). The Ribo-Zero™ Magnetic Kit (Epicentre Technologies, Madison, WI, USA) was applied to remove ribosomal RNA. Sequencing library was then prepared using the TruSeq RNA Library Preparation Kit (Illumina, San Diego, CA, USA) in accordance with the Illumina protocol and then evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA). RNA libraries were sequenced with 150 bp paired-end read lengths on an Illumina HiSeq 4000 platform (Illumina).
Sequence quality was assessed using FastQC software v0.10.1 (www.bioinformatics.babraham.ac.uk/projects/fastqc/). Trim Galore (www.bioinformatics.babraham.ac.uk/projects/trim_galore/) was used to trim low-quality bases and adaptors. The expression patterns of circRNAs were identified using CIRI software (Gao et al., 2015), followed by matching with data from the circBase database (http://circbase.org/) (Glažar et al., 2014). circRNA expression was normalized using Back Spliced Reads per million mapped reads. Hierarchical clustering maps were used to evaluate circRNA expression variation.
Differential circRNA identification using DEGSEQ software (Wang et al., 2010), with standardized read numbers, was utilized to calculate the statistically significant differential expression of circRNAs among different groups of samples (fold change ≥2.0 and p ≤ 0.05). Molecular data were integrated from genomics, transcription, proteomics, and metabolomics to infer the pathways and biological functions involved in differentially expressed circRNA using KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway analysis. Differential circRNA-miRNA interactions were predicted to explore the functional role of circRNAs used as “miRNA sponges.” The circRNA-miRNA network was constructed and displayed using Cytoscape software.
Quantitative reverse transcription-polymerase chain reaction validation assay for top dysregulated circRNAs
Complementary DNA was produced by reverse transcription using a PrimeScript™ RT Reagent Kit (TaKaRa Bio, Nojihigashi, Kusatsu, Japan) according to the manufacturer's instructions. The circRNA expression in 12 healthy adults, 12 individuals with LADA, and 12 type 2 diabetes were measured by quantitative real-time PCR (qPCR) using SYBR® Green Master Mix (TaKaRa, Tokyo, Japan) in a ViiA™ 7 Real-time PCR System (Applied Biosystems, Inc., Foster City, CA, USA). RNA relative expression was calculated by the 2−ΔΔCt method.
All qPCRs were repeated three times. A single peak in the melting curve indicates the specificity of the PCR product.
Statistical analysis
SPSS Statistics software (version 22.0; IBM, Armonk, NY, USA) was applied for statistical analysis and figures were generated using GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA, USA). Categorical data were analyzed by Chi-square test. Data were presented as mean ± standard deviation of at least three independent experiments. p < 0.05 was considered statistically significant. The most representative circular RNAs were annotated using circBase or Circ2Traits databases (Glažar et al., 2014). These circular RNAs were selected on the basis of differences among groups identified by data analysis and the significance of their corresponding genes.
Results
Baseline characteristics
Baseline characteristics of all participants are summarized in Table 1. No statistically significant differences were found among individuals in three groups in terms of sex, age, percentage of hypertension, and percentage of smoker (all p-value >0.05). Compared with individuals in group NG, group LADA had lower levels of body mass index (BMI), and higher levels of total cholesterol (TC), triglyceride (TG), alanine aminotransferase (ALT), aspartate aminotransferase (AST), hemoglobin A1c (HbA1c), and glucose (GLU) (all p < 0.05); Group DM had higher levels of BMI, TC, TG, ALT, AST, HbA1c, and GLU compared with group NG (all p < 0.05). Group LADA had lower levels of BMI, TG, and higher levels of HbA1c compared with group DM (all p < 0.05).
Differentially expressed circRNA identification and validation
A total of 2334 differentially expressed circRNAs were detected in the three groups, with 277 circRNAs in the Group DM versus Group NG; 992 circRNAs in the Group LADA versus Group NG; and 1065 circRNAs in the Group DM versus Group LADA. These circRNAs were distributed in all chromosomes. Compared with Group NG, 167 circRNAs were significantly upregulated and 110 circRNAs downregulated in the Group DM; Compared with Group DM, 218 circRNAs were significantly upregulated and 847 circRNAs downregulated in the Group LADA. Compared with Group NG, 209 circRNAs were significantly upregulated and 783 circRNAs downregulated in the Group LADA. Principal component analysis (PCA) was first performed to assess the sample correlations using the expression data of all altered circRNAs. PCA clustering show that individuals from Group DG and Group NM cluster together compared with individuals from Group LADA based on circRNA expressions.
The samples of LADA group were well clustered and distant from those of NM and DG groups (Fig. 1). Heat maps depicted the hierarchical clustering of these altered circRNAs in three groups are shown in Figure 2 (|fold change| > 2 and p-value <0.05). Scotty tool (Busby et al., 2013) (http://scotty.genetics.utah.edu) was applied for statistical power analysis, using the gene expression results obtained from the current study. It indicated that with an average of 10 M reads and four replicates per treatment condition, one can identify 80% of the genes with at least 50% maximum power with at least 2 × fold change at a false discovery rate (FDR) of 0.05. We, therefore, conclude that our study, which included four replicates for each treatment and an average read depth of 10 million reads had sufficient statistical power to detect differential gene expression.
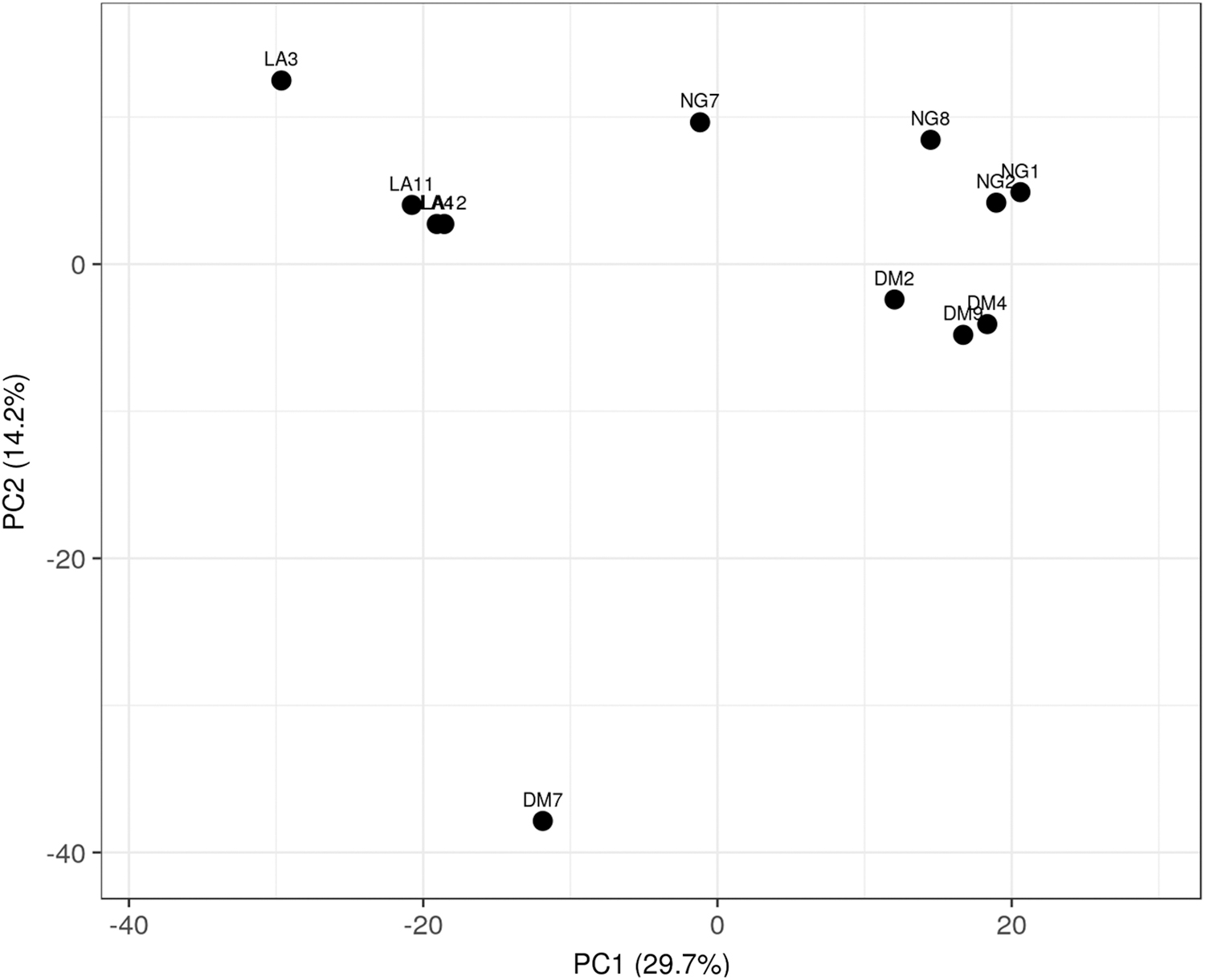
PCA depicts the sample correlations using the expression data of all altered circRNAs. LADA participants (LA), T2DM participants (DM), and healthy participants (NG). circRNA, circular RNA; LADA, latent autoimmune diabetes in adults; PCA, principal component analysis; T2DM, type 2 diabetes mellitus.

The heat map depicts the hierarchical clustering of altered circRNAs in three groups. Red represents upregulation while green represents downregulation.
KEGG pathway enrichment analysis of the circRNA host genes showed an overrepresentation of genes involved several crucial pathways, including tumor necrosis factor (TNF) signaling pathway, FoxO signaling pathway (enriched for host genes of downregulated circRNA in group DM compared with group NG), cellular senescence (enriched for host genes of downregulated circRNA in group LADA compared with group NG) and long-term potentiation (enriched for host genes of upregulated circRNA in group LADA compared with group DM) (all FDR p < 0.05). Detailed information can be found in Table 2.
Kyoto Encyclopedia of Genes and Genomes Pathway Enrichment Analysis of the Circular RNA Host Gene
PathwayID: Pathway identifiers used in KEGG; Definition: definition of the Pathway; Fisher-p-value: enrichment p-value of the PathwayID used Fisher's exact test; GeneRatio: Count of the DE genes' entities directly associated with the listed PathwayID divided by the total number of the DE genes' entities; BgRatio: count of the chosen background population genes' entities associated with the listed PathwayID divided by the total number of chosen background population genes' entities; FDR: the false discovery rate of the PathwayID; GENES: Host genes associated with the PathwayID. Type: the origination of the differentially expressed circRNA.
circRNA, circular RNA; KEGG, Kyoto Encyclopedia of Genes and Genomes; TNF, tumor necrosis factor.
After screening and comparison, six highly dysregulated circRNAs are listed in Table 3, which were selected according to the following conditions: (1) Basic parameters: |fold change| > 2 and p-value <0.01; (2) Exon source for subsequent functional research; (3) circRNAs themselves or derived genes have been reported associated with diabetes, two of which were recognized as unannotated, new circRNAs in the circBase or Circ2Traits database. According to the txStart genome coordinates of the circRNAs, we named them hsa_circ_62540520 and has_circ_100670302. The expression of these circRNAs have been validated in the whole cohort by quantitative reverse transcription-polymerase chain reaction (qRT-PCR) (Fig. 3). Detailed information of primer sequences can be found in Supplementary Table S2.
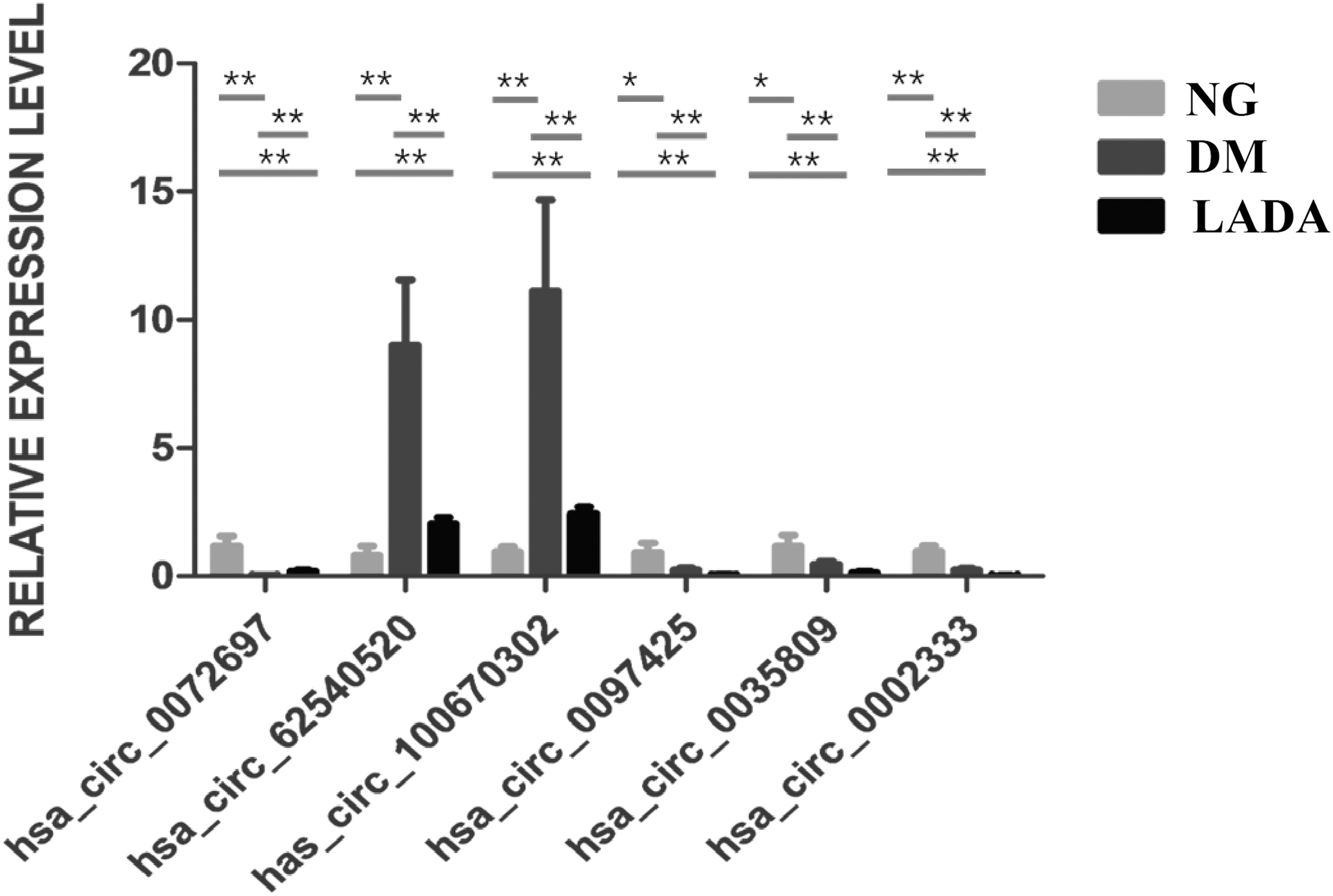
Validation of the top 6 dysregulated circRNAs by qRT-PCR in the three groups. The results show the mean ± SD. *p < 0.05; **p < 0.01. qRT-PCR, quantitative reverse transcription-polymerase chain reaction; SD, standard deviation.
Top 6 Dysregulated Circular RNAs
down_DM_vs_NG: downregulated in DM compared with NG; up_DM_vs_NG: upregulated in DM compared with NG; down_LADA_vs_DM: downregulated in LADA compared with DM; down_LADA_vs_NG: downregulated in LADA compared with NG.
Prediction of circRNA-miRNA interaction and the circRNA network
We predicted the potential target miRNAs and coding genes of hsa_circ_62540520, hsa_circ_100670302, hsa_circ_0072697, hsa_circ_0097425, hsa_circ_0035809, and hsa_circ_0002333 using MiRanda and TargetScan software. The network diagram shows the top 5 miRNAs that may bind to circRNA and several of the most likely target genes for each miRNA, and clearly identifies six potential targets for differentially expressed circRNAs. These findings lay the foundation for studying the specific mechanisms of these circRNAs in different types of diabetes (Fig. 4).
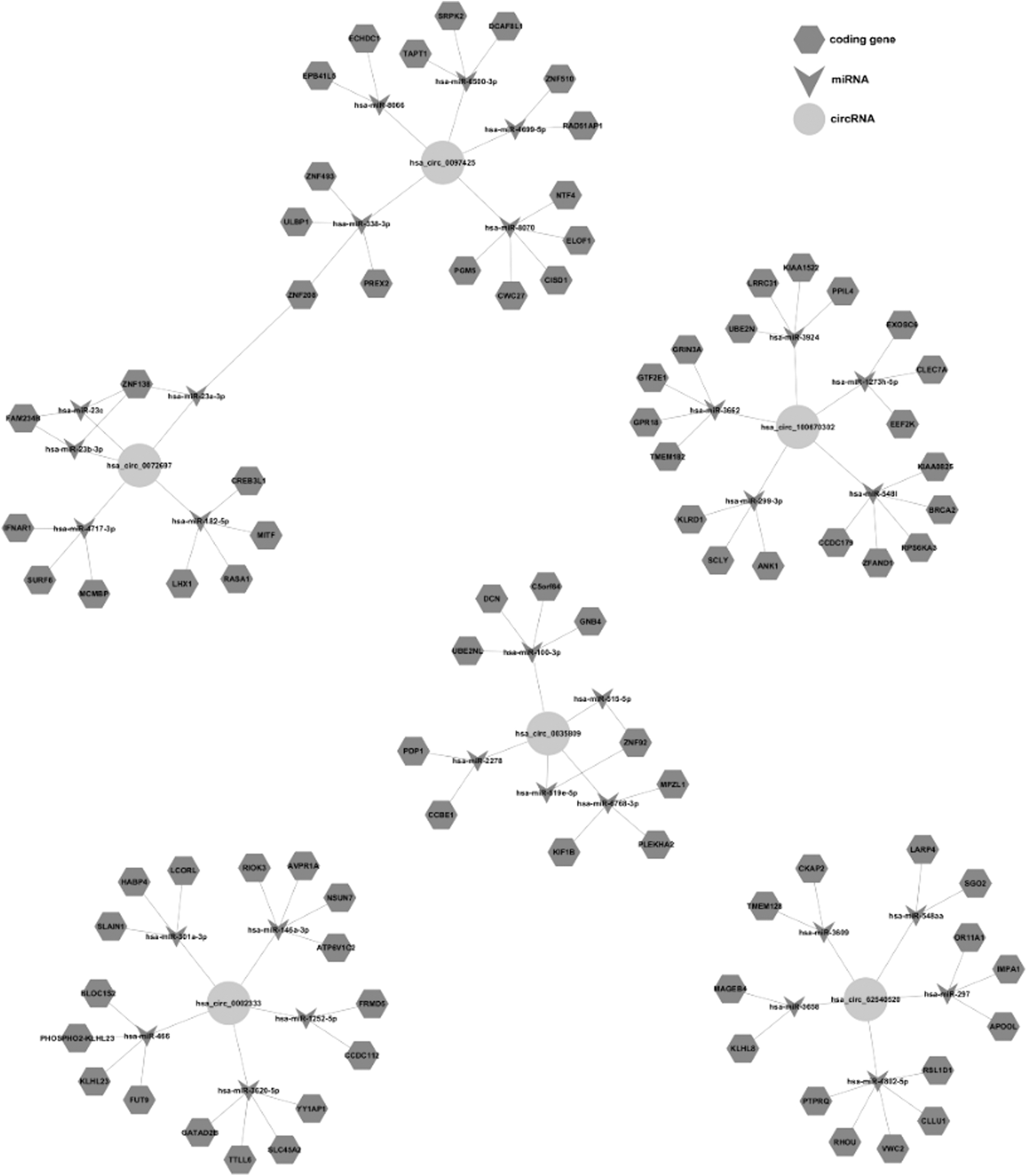
The network diagram of the top 5 miRNAs that may bind to circRNAs and several of the most likely target genes for each miRNA. miRNA, microRNA.
Discussion
According to the Diabetes Atlas of International Diabetes Federation, 10 million people are newly diagnosed with diabetes every year (Rahelić, 2016). A Chinese LADA study found that rate of LADA in individuals over 30 years of age, newly diagnosed with T2DM was 5.9% (Zhou et al., 2013). LADA is currently classified as T1DM, but its clinical manifestations are similar to T2DM. The autoantibodies of the patients with LADA indicate the pathogenesis of autoimmune diseases. However, the autoimmune process of LADA seems to be milder, and the procession of islet β cell failure is slower and consistently shows a higher level of C-peptide as an indicator of insulin secretion, indicating that other mechanisms also play an important role in the pathogenesis of LADA (Hernandez et al., 2015).
Studies have shown that the number of natural killer cells in individuals who were newly diagnosed with LADA is similar to that in individuals with type 1 diabetes, but lower compared with individuals with type 2 diabetes (Rahelić, 2016). The degree of insulin resistance in individuals with LADA is similar to that in individuals with T2DM (Chiu et al., 2007). Another Chinese LADA study found that the prevalence of metabolic syndrome in individuals with LADA is slightly lower in those with T2DM, but higher in those with type 1 diabetes and healthy individuals (Xiang et al., 2011). To summarize, while some of the relationships and differences among individuals with type 1 diabetes, type 2 diabetes, and LADA have been explored, the nature of these connections has not yet been fully elucidated.
Circular RNA was first discovered by scientists using electron microscopy to observe the virus in the 1970s. With the development of scientific and technological advances, especially high-throughput sequencing and bioinformatics, we gradually uncover the mystery of circRNAs. Numerous studies have shown a strong link between circRNA and diabetes. A study investigates the characteristic expression of circRNAs in the peripheral blood of T2DM patients; hsa_circ_0054633 was found to be used as a diagnostic biomarker of prediabetes and T2DM (Zhao et al., 2017).
Functional analysis and prediction of differently expressed circRNA profiles in T2DM patients with or without depression found that hsa-circrna_003251, hsa-circrna_015115, hsa-circrna_100918, and hsa-circrna_005019 may serve as biomarkers and potential therapeutic targets for future intervention studies, diagnosis and treatment of T2DM patients with depression (Jiang et al., 2017).
In addition, with the aging of population and the rapid increase in the number of cardiovascular disease (CAD) deaths, Li et al. (Li et al., 2017) confirmed for the first time that some circRNAs were related to T2DM with CAD, among which, hsa-circRNA11783-2 had the strongest related to T2DM with coronary artery disease, which contributed greatly to the study of T2DM with CAD. These studies show that circRNA is closely related to the occurrence and development of diabetes, which has an excellent research prospect.
This study screened differentially expressed circRNAs from blood samples of participants with LADA, T2DM, and matched healthy controls by high-throughput sequencing. Pair grouping allowed us to identify circRNAs that were aberrantly expressed between two groups. We finally identified the six most distinctly expressed circRNAs, then verified them by qRT-PCR. Host gene of the hsa_circ_100670302 gene, VPS13B, is associated with insulin resistance.
According to literature (Limoge et al., 2015), in VPS13B-deficient cells, adipocyte differentiation is accelerated, fat storage is increased, and insulin response is dysfunctional. Insulin resistance is a critical factor in diabetes pathogenesis, especially in T2DM (Sampath Kumar et al., 2019). Our results showed that hsa_circ_100670302 was obviously upregulated in participants with T2DM relative to the control group. Therefore, we hypothesize that hsa_circ_100670302 affects the occurrence and development of diabetes by regulating the VPS13B gene.
Metabolic syndrome refers to the pathological status of metabolic disorders of proteins, fats, carbohydrates, and other substances in the human body (Hudish et al., 2019). It is a group of complex metabolic disorders, which is a risk factor for dyslipidemia, hyperglycemia, and cerebrovascular diseases. Individuals with metabolic syndrome often show a state of proinflammation. The change in cytokine expression may be one of the mechanisms of low inflammation accompanying disorder of lipid and glucose metabolism (Saklayen, 2018).
Host gene of the hsa_circ_0097425 gene, HECTD4, has been reported to be a pleiotropic gene that regulates metabolic syndrome and inflammation (Kraja et al., 2014). In our results, hsa_circ_0097425 was obviously downregulated in participants with LADA relative to the T2DM group. We speculate that hsa_circ_0097425 regulates metabolic syndrome and inflammation through HECTD4, and acts as a protective circRNA.
Besides β cell destruction, other autoimmune mechanisms, including immune defense may play a role in the pathogenesis of type 1 diabetes mellitus (Lee, 2014). For instance, many studies have shown that LADA is related to T cell-mediated immune response (Tard et al., 2015). The combination of SPRY and PRY to produce B30.2 domains may be related to immune defense (Rhodes et al., 2005). The SPRY domain was subsequently identified in a total of 10 distinct protein families, including human HERC1. SPRY domains were subsequently identified in the protein family of HERC1. Our results showed that hsa_circ_0035809 was clearly downregulated in participants with LADA relative to the normal control group. We inferred that hsa_circ_0035809 may be used as an “miRNA sponge” that regulates posttranscription by combining competitively with miRNA and affects HERC1 to regulate immune defense. Individuals with LADA display insulin resistance (Hjort et al., 2018), although not as pronounced as displayed by those with T2DM, and this is due to differences in BMI (Juhl et al., 2014). This has also linked LADA to several of the aforementioned genes.
Recent studies have shown that circRNAs are derived from the exons or introns of their host genes and may regulate the expression of these host genes (Lasda and Parker, 2014). The results of KEGG analysis revealed that the host genes of significantly dysregulated circRNAs were involved in many important pathways, among which, TNF signaling pathway, FoxO signaling pathway were enriched for host genes of downregulated circRNA in group DM compared with group NG.
TNF-α pathway was recently found to activate early during the development of diabetic nephropathy in Type II diabetes mellitus (Lampropoulou et al., 2020). FoxO proteins are well known for researchers in the field of metabolism disease and can regulate insulin signaling, gluconeogenesis, insulin resistance, immune cell migration, and cell senescence (Gross et al., 2009); circRNAs derived from FoxO signaling pathway genes were very likely to involve in the pathology of T2M through regulating the function of FoxO.
Cellular senescence is a fundamental mechanism of aging and appears to play a crucial role in the onset and prognosis of CAD in the context of both aging and diabetes (Shakeri et al., 2018). Since this pathway was enriched for host genes of downregulated circRNA in group LA compared with group NG and cellular senescence has been known for years to occur in immune cells, it might be a unique pathway for the function of circRNAs in the development of LADA (Vicente et al., 2016).
Long-term potentiation was identified enriched for host genes of upregulated circRNA in group LADA compared with group DM, however, it is not clear whether long-term potentiation reflects the difference between LADA and T2DM. More work is needed to elucidate the function of genes in long-term potentiation pathway.
There are several limitations to our study. First, we used a relatively small sample size, hence there could be a possibility of overestimating the significance of the association and a great chance of missing the true signal. Meanwhile, it is important to mention that the limitation of a small sample size might influence the data interpretation and study conclusion due to individual variability.
An additional limitation of our study represents the inability to generalize study findings to other populations. We will initiate further follow-up studies, recruiting and characterizing more non-Chinese Han patients with DM and LADA, especially more patients with T1D, another type of diabetes phenotype. As functional analysis of the identified circRNAs was not the main scope of the present study, a further limitation of the present study is the difficulty to judge whether the identified circRNAs originate from disease-specific mechanisms or from more unspecific biological processes.
Conclusion
Our study is the first to analyze circRNAs of Chinese participants with LADA and T2DM. Additional research is needed to verify circRNAs as a diagnosis and treatment target for individuals with different types of diabetes in the future. Our findings need to be further validated in larger, more diverse samples.
Ethics Approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the Institutional and/or National Research Committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Informed Consent
Informed consent was obtained from all individual participants included in the study.
Availability of Data and Materials
The statistics data used to support the findings of this study are available from the corresponding author upon request.
Footnotes
Authors' Contributions
L.N. and Y.Y. drafted the initial article. Y.Y. and X.F. analyzed the data; Y.C. and M.L. have interpreted the results; X.Z., H.S., M.Y., and L.W. have contributed to the literature search and bioinformatic analysis; H.C. has conceived the idea of the study and edited the article. All authors read and approved the final article.
Acknowledgments
The authors thank Cloud-Seq Biotech Ltd. Co. (Shanghai, China) for providing circRNA-Seq analysis, and Mr. Tianqi Wang for help during the study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Jilin Province Health and Health Technology Innovation Project (2018J047).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.