
Abstract
Background:
Gastric cancer (GC) is the third leading cause of cancer-related mortality worldwide, and single nucleotide polymorphisms (SNPs) in microRNAs (miRNAs) are believed to affect the occurrence and progression of cancer by altering the expression and biological functions of miRNAs.
Methods:
The present scoping review was designed to evaluate and discuss microRNA SNPs (miR-SNPs) that have been found to be associated with GC in the following two contexts: (1) the biological effects on GC based on SNP localization; and (2) the associations between miRNA-SNPs and clinical factors (susceptibility, tumor size, metastasis, overall survival, and prognosis) of GC.
Results and Conclusions:
Information on miRNAs was collected, including the SNPs, their proven target genes, and the possible impact of the SNPs on GC outcome. Our findings suggest an etiological or modifying role for multiple miRNA SNPs (miR-499, miR-146a, miR-149, miR-148, miR-27a, miR-608, miR-196a-2) in GC and its progression. The findings of this study reinforce the multiple roles of miRNA SNPs in GC.
Introduction
Gastric cancer (GC) is the fifth most frequently diagnosed type of cancer, and the third leading cause of cancer-related mortality worldwide (Bray et al, 2018). The development of GC is a multistep process, involving both genetic and environmental factors. Despite some advances in therapies, the long-term survival of patients with advanced-stage GC remains poor (Alessandrini et al, 2018).
In recent years, many studies have demonstrated that microRNAs (miRNAs) can regulate the expression of multiple genes that are closely associated with the occurrence and development of GC and may therefore be valuable in its diagnosis and treatment. miRNAs are a class of noncoding, endogenous, short, single-stranded molecules ∼18-22 nucleotides in length, which regulate gene expression by binding to the 3′ untranslated region (UTR) of target mRNAs, leading to their degradation or translational repression based on the degree of complementarity (Moazeni-Roodi et al, 2019). In the canonical biogenesis pathway, pri-miRNAs are transcribed from their genes and then processed into pre-miRNAs by the microprocessor complex, consisting of an RNA-binding protein DiGeorge Syndrome Critical Region 8 (DGCR8), and a ribonuclease III enzyme named Drosha.
Then, pre-miRNAs are exported to the cytoplasm by Exportin-5, which creates the microRNAs by the action of Dicer. microRNAs function by forming miRNA-RISC complex, which binds to the 3′UTR of the target mRNA (Fig. 1) (O'Brien et al, 2018). miRNAs are often dysregulated in cancer. Multiple mechanisms can lead to miRNA dysregulation, including deletion, amplification, mutation, transcriptional dysregulation, and epigenetic alterations (Lujambio et al, 2007). Increasing efforts are being made to identify genetic variants of miRNA genes that may be relevant to cancer risk or prognosis.
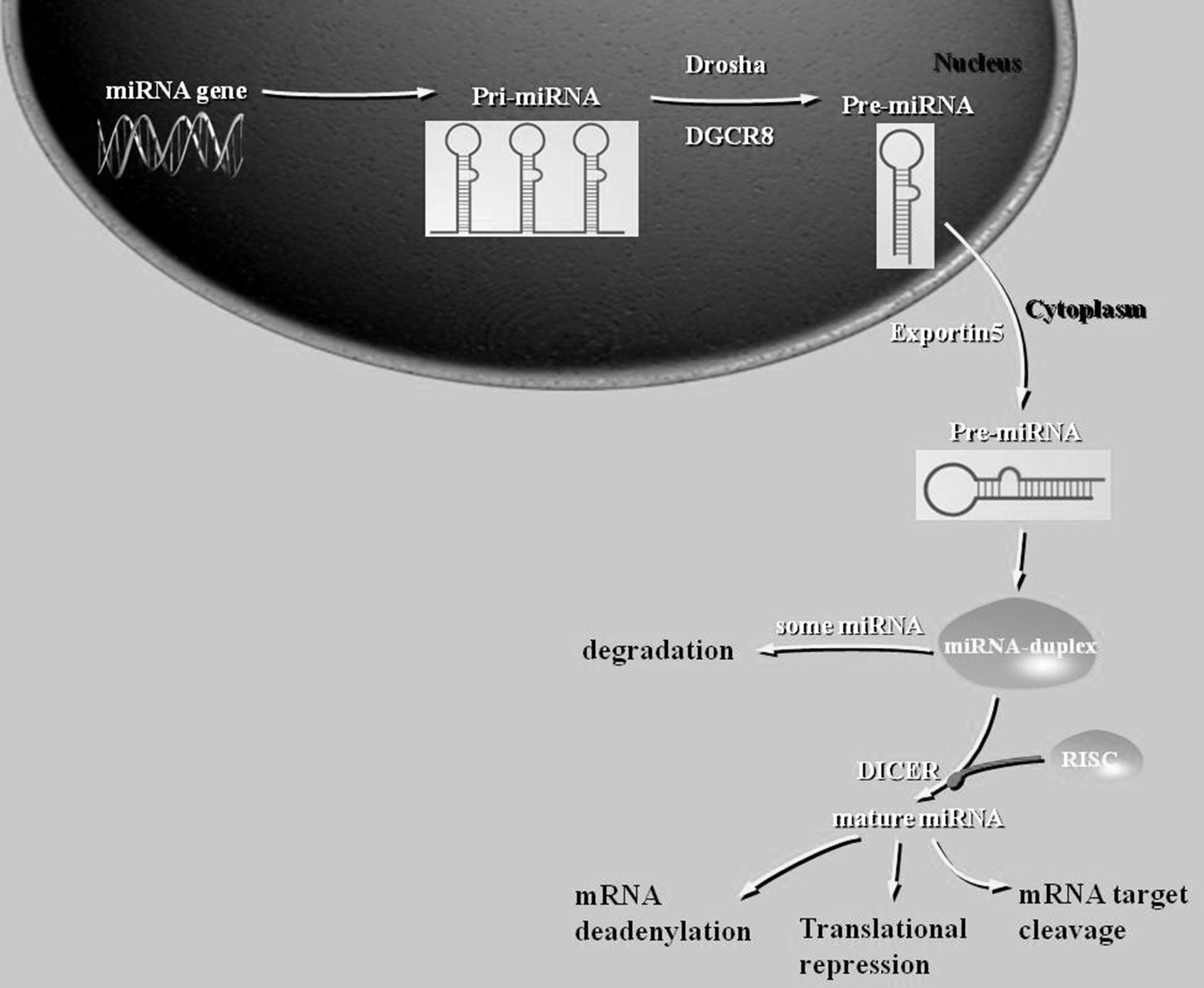
Biogenesis of miRNA:Drosha/DGCR8 complex recognizes the double-stranded RNA structure of the pri-miRNA and specifically cleaves at the base of the stem-loop, releasing the pre-miRNA, which then is exported to the cytoplasm byExportin-5; Dicer complex then creates the mature miRNA duplex by affecting the pre-miRNA. The translation suppressing function of miRNA is achieved when the miRNA-RISC complex is formed and attached to the 3′-UTR of the target mRNA. 3′-UTR, 3′-untranslated region; mRNA, messenger RNA; pri-miRNA, primary microRNA; pre-miRNA, precursor microRNA; RISC, RNA-induced silencing complex.
Single nucleotide polymorphisms (SNPs) are the most common type of genetic variation in the human genome. It has been found that SNPs not only exist in coding genes, but also in noncoding genes, such as miRNA-related SNPs. miRNA-related SNPs include those located in the promoter region of miRNAs, in the sequence of miRNA genes, in the site where miRNAs bind to their target genes, and in the biosynthetic pathways of miRNAs. SNPs can regulate the function of miRNAs in the following three ways: (1) by regulating the transcription of pri-miR, (2) by affecting the processing and maturation of pri-miR to pre-miR, and (3) by affecting the binding of the miRNA to its downstream genes. SNPs thus regulate the transcription, expression, and biological function of the target genes, ultimately affecting the susceptibility of individuals to diseases, as well as the effectiveness of drug therapies.
A growing body of evidence suggests that miRNA-associated SNPs can be used as surrogate markers to predict tumor prognosis (Moriyama et al, 2009; Saetrom et al, 2009; Wang et al, 2019). Studying SNPs is valuable, as the data thus extracted can be used for personalized medicine. The focus of the present review was to investigate miRNA-related SNPs in GC using two criteria: (1) the localization of microRNAs SNPs (miR-SNPs), (2) the published data processing of the relationship between miR-SNPs and genetic susceptibility, risk of disease, pathological characteristics as well as prognosis, and collection of data on functional regions of miRNA genes, effects of miRNA SNPs, other possible miRNA gene subtypes, and their roles in cells. To the best of our knowledge, to date, there is no literature reviewing miR-SNPs in GC.
Materials and Methods
The SNPs discussed in this study were searched through the literature and the DBSNP database (www.ncbi.nlm.nih.gov/snp/). We searched PubMed (https://pubmed.ncbi.nlm.nih.gov/) and GCBI (www.gcbi.com.cn/gclib/html/index) databases using the following terms: “miRNA,” “polymorphism,” “gastric cancer,” “gastric tumour” and “Stomach Neoplasms” (last search update: March 16, 2021).
Results
The localization of miR-SNPs
SNPs in miRNA promoter regions
The mechanism of transcriptional regulation of miRNA genes is similar to that of protein-coding genes. Transcription factors usually bind to the conserved sequences upstream of the genes, and thus regulate their transcription. For example, rs4719839, located in the miR-148a promoter region ∼2966 bp upstream from pre-miR-148a, might affect the posttranscriptional regulation of miR-148 by binding to different transcription factors, which could be the reason behind lymph node metastasis (p = 0.047) and tumor invasion (p = 0.009) in GC in a large sample of the Han population of Northern China (Chen et al, 2014). SNPs in miR-196a2 promoter region may directly trigger the transcription of miR-196a2, and thus affect susceptibility to GC (Xu et al, 2014). The SNP in the promoter region of pri-miR-34b/c (rs4938723) may alter the binding efficiency of transcription factor GATA (Pan et al, 2015).
SNPs in pri-miRNAs and pre-miRNAs
An increasing number of studies have shown that SNPs in pri-miRNA and pre-miRNA genes may interfere with mature processing or degradation of miRNAs by altering the efficiency of pri-miRNA conversion to pre-miRNA or pre-miRNA conversion to mature miRNAs, and thus compromising the secondary structure and causing a reduction in the amount of mature miRNA (Króliczewski et al, 2018). This has been demonstrated by Duan and coworkers in 2007. Furthermore, they hypothesized two possibilities for how the miR-125a SNP might alter the processing of pri-miRNA. One hypothesis was that there might be other structural requirements for efficient processing of pri-miRNA by the Drosha-DGCR8 complex. Another hypothesis was that other proteins might bind to pri-miRNA and regulate the processing of pri-miRNAs. These interactions might contribute to miRNA specificity or differential expression, and the miR-125a SNP may be a candidate regulator of such interactions (Duan et al, 2007).
Most SNPs listed in the Table 1 (rs3746444 in miR-499b; rs2910164 in miR-146a; rs2292832 in miR-149; rs895819, rs11671784 in miR-27a) we collated were located at this position. The rs11614913 C allele could accelerate the process of conversion of pre-has-mir-196a2 to its mature form, although the variant rs11614913 CC genotype was associated with a significant reduction in the risk of GC (adjusted odds ratio [OR] = 0.78, 95% confidence interval [CI] = 0.62-0.99 for the test set, and 0.64, 0.52-0.80 for the validation set), compared to the CT/TT genotypes. Furthermore, the CC genotype was associated with a significantly increased survival in GC, compared to the CT/TT genotypes (adjusted hazard ratio [HR] = 0.72, 95% CI = 0.55-0.95) (Wang et al, 2013b). The underlying mechanism is not yet clear, and further research on this aspect is therefore required.
miR Related Single Nucleotide Polymorphisms Involved in Gastric Cancer and Their Genic Information
SNPs in mature miRNAs
The critical region for miR binding in animals is the “seed sequence” (nucleotides 2-7 from the 5′end of the miR), which most often binds to a target site in the 3′ UTR of the given mRNA. The seed sequence is highly conserved, because of which the probability of SNP in the seed sequence is much lower than that in other parts. Only 1% of the miRNAs had SNPs in the functional seed region (Saunders et al, 2007). The SNP located in the seed sequence of miR-125a could significantly inhibit the processing and production of pri-miRNA and pre-miRNA, thereby inhibiting miRNA expression (Duan et al, 2007).
SNPs in miRNA target gene-binding site
mRNA target selection by miRNA is based on the complementary pairing of a 3′UTR mRNA to the seed sequence. In addition, the effect of G:U base pairs and bumps in the seed sequence and additional mRNA pairing to the 3′ region of miRNA, called supplement seed pairing (3′ supplementary sites and 3′ compensatory sites), also play a crucial role. Thus, the introduction of SNPs into mature miRNAs may have genome-wide effects, altering hundreds of potential mRNA targets and having unpredictable impacts on cellular homeostasis (Króliczewski et al, 2018).
It has been reported that the variants rs2240688 C and rs3130 C > T in the miRNA-binding site of the stem cell marker gene CD133 are associated with GC risk and reduction in overall survival (Wang et al, 2015). The miR-29c-binding site SNP (rs11944405 T>C) in the 3′-UTR of LAMTOR3 decreased the GC risk. (TC vs. TT, OR = 0.79, 95% CI = 0.63-0.99; TC/CC vs. TT, OR = 0.81, 95% CI = 0.65-1.00) (Song et al, 2015). Moreover, Chen et al (2015a) found that rs114673809 modified the binding of hsa-miR-214 to MTHFR, and thus altered MTHFR protein levels in GC patients, showing that subjects carrying the rs114673809 AA genotype had a significantly increased GC risk (OR = 1.667, 95% CI = 1.044-2.660, p = 0.034) compared to the GG genotype.
SNPs in genes associated with miRNA biosynthesis pathways
SNPs in genes involved in the miRNA biosynthesis pathways can affect the expression of miRNAs in general, by affecting gene function (Arroyo et al, 2017). The rs11170877 polymorphism is located in an exon of the host gene COPZ1 of miR-148b, which may contain a missense mutation that changes Met to Val, resulting in the production of shorter than normal peptides, leading to the development of GC. Also, the rs11170877A>G mutation has been identified as a protective factor for GC risk in the northeastern Chinese population (p = 0.027) (Chen et al, 2014). Moreover, miR-SNPs in genes encoding microRNA processing machinery, including the C/C genotype of rs14035 from RAN, the A/A genotype of rs3742330 from Dicer, and the T/T genotype of rs9623117 from TNRC6B, have all been found to be significantly associated with GC risk (Xie et al, 2015).
Analysis of the associations between miR-SNPs and GC
SNPs of miRNAs and susceptibility to GC
As biomarkers of cancer susceptibility, miRNAs have attracted increasing attention. Various studies have linked genetic variations in miRNA-encoding genes and their target genes to GC risk. With respect to GC susceptibility, SNPs in miR-149 rs2292832 (TT vs. TC+CC: OR = 1.15, 95% CI: 1.03-1.30, p(h) = 0.68, p = 0.02) and (TT vs. TC + CC, recessive model: OR and 95% CI, 1.24 [1.04-1.48]; T vs. C, allele model: OR and 95% CI, 1.13 [1.00-1.28]) (Li et al, 2013; Zhang et al, 2018); miR-27a rs895819 (AG+GG vs. AA: OR = 1.43, 95% CI = 1.01-2.02, p = 0.04), and miR-499 rs3746444 (TT vs. TC+CC: OR = 1.491, 95% CI = 1.155-1.923, p = 0.002) (Cai et al, 2015; Song et al, 2014) were associated with increased GC risk among Asians, and Dikeakos Pet al reported similar results in miR-149 rs2292832 (p = 0.009) in the Greek population (Dikeakos et al, 2014).
Qian et al demonstrated that gene-gene interactions produce a greater effect than a single gene in determining cancer susceptibility, and concluded that let-7e rs8111742 and miR-4795 rs1002765 SNPs interacted with H. pylori infection to increase GC risk in a Chinese population (rs8111742: P interaction = 0.024; rs1002765: P interaction = 0.031, respectively) (Xu et al, 2016).
Similarly, there have been several studies on the reduction of susceptibility to GC. Polymorphisms of miR-148b (rs11170877 A>G G allele, p = 0.027 and rs12231393 T > C C allele, p = 0.034) (Chen et al, 2014); miR-25 (rs41274221), and the upregulation of miR-25 with the AA genotype (OR = 0.67, 95% CI = 1.33-1.39, p = 0.001) (Zhou et al, 2016); SNPs in the miR-196a2 promoter rs35010275 G > C were found to be protective factors against susceptibility to GC (OR = 0.85, 95% CI = 0.77-0.94) (Chen et al, 2014). Li et al (2017) suggested that miR-1269a rs73239138 AA genotype (p = 0.045, OR = 0.610, 95% CI = 0.376-0.990) played role in decreasing the susceptibility to GC by downregulating its expression and targeting ZNF70, which promoted apoptosis of GC cells.
Studies by Pan et al (2015) (CT vs. TT: OR = 0.66; 95% CI = 0.45-0.97; and CT/CC vs. TT: OR = 0.67; 95% CI = 0.47-0.97) and Yang et al (2014a) (CC vs. TT: p = 0.006, OR = 0.53, 95% CI = 0.34-0.83; C vs. T: p = 0.005, OR = 0.75, 95% CI = 0.61-0.92) showed that the C allele of the miR-34b/c rs4938723 was associated with a significantly decreased GC risk, suggesting that miR-34b/c rs4938723 might individually and jointly exert a protective effect against GC.
Previous studies have also revealed that SNPs of miR-933 (rs79402775), miR-92a (rs9589207), miR-27a (rs11671784), miR-107 (rs2296616), miR-124-1 (rs531564), and the rs35010275 C allele, were significantly associated with a decreased GC risk, and may therefore be useful as biomarkers for GC prognosis (Chen et al, 2014; Liu et al, 2016; Mirnoori et al, 2018; Moazeni-Roodi and Hashemi, 2018; Soleimani et al, 2016; Song et al, 2014; Wang et al, 2014; Yang et al, 2014b; Zhang et al, 2017).
SNPs of miRNAs and prognosis of GC
Abnormal miRNA expression has been widely recognized as a marker of tumor prognosis. Accumulating evidence suggests that miRNA-associated SNPs could be useful as alternative markers to predict tumor prognosis. Patients with a functional genetic variant of miRNA-148a rs4719839 with the G allele were found to be more likely to have Borrmann type III-IV (AG+GG vs. AA, OR = 1.383, 95% CI = 1.027-1.862, p = 0.034), increased tumor size (AG+GG vs. AA, OR = 1.226, 95% CI = 1.037-1.450, p = 0.020), lymph node metastasis (AG+GG vs. AA, OR = 1.548, 95% CI = 1.062-2.255, p = 0.023), and advanced Tumor Node Metastasis stage (p = 0.009) (Chen et al, 2014). In case of G allele of miR-421, patients with GG genotype had a higher frequency of lymph node metastasis (OR = 1.846; 95% CI = 1.148-2.965).
In survival analysis, GG carriers had shorter survival times than GC carriers, (Jin and Yu, 2016) and C allele of miR-499 rs3746444 might be associated with a larger tumor size (p = 0.015) (Cai et al, 2015). AA genotype of miR-30c rs928508 promoted the maturation of pre-miR-30c, and also facilitated the development of GC with lymph node metastasis in a Chinese patient population (OR = 2.23, 95% CI: 1.07-4.64, p = 0.032) (Mu and Su, 2012). The variant homozygote CC genotype in miR-196a2 displayed a strong association with lymph node metastasis in GC (OR = 2.25, 95% CI = 1.21-4.18, p = 0.011) (Peng et al, 2010).
On the contrary, some SNPs have a positive effect on prognosis. The A > G SNP in 12nt of miR-379 rs61991156 (AA vs. AG: OR = 2.64, 95% CI = 1.02-1.34; AA vs. GG: OR = 0.60, 95% CI = 1.12-1.31; AA vs. G carrier: OR = 3.04, 95% CI = 1.19-1.29) (Cao et al, 2018); the G/A and G/G genotypes of rs11134527 in miR-218 (OR = 0.75, 95% CI = 0.61-0.95, p = 0.016) (Wu et al, 2017b); the C allele of polymorphism in miR-146a rs2910164 (p = 0.041) (Liao et al, 2015); the variant rs11614913 CC genotype of miR-196a2 (HR = 0.72, 95% CI = 0.55-0.95) (Wang et al, 2013b); the rs3746444 A allele carrier in miR-499 in Japan (HR = 0.33, 95% CI = 0.18-0.75) (Tahara et al, 2014); and all the above-mentioned in a Chinese population might be associated with low glycolysis levels, higher cell differentiation, low occurrence, and lower aggressiveness, as well as better prognosis of GC.
In Korea, patients with intestinal-type GC with the miR-146a CG + GG genotypes had significantly higher survival rates than those with the CC genotype, and patients with the miR-499 AA genotype had significantly higher survival rates than those with the AG + GG genotypes. When miR-146a CG + GG and miR-499 AA genotypes were combined, the survival rate of patients with intestinal-type GC increased (Ahn et al, 2013).
Stenholm et al identified three miRNA polymorphisms: rs7372209 in miR-26a1 (HR = 1.307, 95% CI = 1.031-1.656, p = 0.0272), rs895819 in mir-27a (HR = 1.304, 95% CI = 1.031-1.650, p = 0.0270), and rs11614913 in mir-196a2 (HR = 0.791, 95% CI = 0.625-1.000, p = 0.0497) as independent prognostic factors in patients with advanced GC (AGC), and found that rs7372209 and rs895819 were associated with a worse prognosis in AGC, and that the variant T allele of rs11614913 (miR-196a2) was associated with improved overall survival among patients with AGC in a German population (Stenholm et al, 2013).
Controversial results regarding the association between SNPs of miRNAs and GC
Several studies have reported the role of the miR-146a rs2910164 G > C polymorphism as a susceptibility factor for several digestive cancers. In China, Zeng et al (2010) and Xie and Wang (2017) observed that the GC+GG genotype of miR-146a was associated with increased risk compared with the CC genotype. Dikeakos et al (2014) reported that the rs2910164 CTT and CCT haplotypes appear to play a protective role against GC in a Greek population. However, other studies did not support this conclusion, as Wei et al (2015) demonstrated that the only significant association between the miR-146a rs2910164 polymorphism and susceptibility to GC was found in the heterozygous model, and it may only marginally contribute to a decreased risk.
In Japan, Tahara et al (2014) indicated that individuals with the CC genotype had a significantly increased GC risk. Wu et al (2013) and Ma et al (2013) observed that the miR-146a rs2910164 G > C polymorphism was not significantly correlated with GC risk and Kupcinskas et al (2014) demonstrated that the rs2910164 (G/C) polymorphism was not significantly associated with GC in Europe. In addition, Chen et al (2015b) suggested that the miR-146a rs2910164 polymorphism was associated with GC risk in Asians, but not in Caucasians.
The association between SNPs in miR-196a2 (rs11614913) and susceptibility to GC have been inconsistent. Compared with the T allele in Korean women, the C allele of miR-196a2 rs11614913 can significantly increase the risk of cancer (Ahn et al, 2013). Wang et al (2013a) suggested that variant C allele could significantly increase risk of GI cancers in different genetic models (C vs. T: OR = 1.17, 95% CI = 1.07-1.28, p = 0.0008; CT + CC vs. TT: OR = 1.26, 95% CI = 1.08-1.48, p = 0.004; CC vs. CT + TT: OR = 1.23, 95% CI = 1.08-1.39, p = 0.002; CC vs. TT: OR = 1.55, 95% CI = 1.24-1.94, p = 0.0001; CT vs. TT: OR = 1.20, 95% CI = 1.02-1.40, p = 0.03).
However, other researchers reported that the variant rs11614913 CC was associated with a significantly reduced risk compared with the CT/TT genotypes (Wang et al, 2013b). Chen et al (2015b) suggested that there was no distinct correlation between the miR-196a-2 rs11614913 polymorphism and the risk of cancer . Ni et al (2015) demonstrated the same result, but they also proved that it might decrease susceptibility to diffuse-type GC. In Japan, Parlayan et al (2014) found that rs11614913 is generally not a GC risk factor, but might affect the risk in the elderly Japanese population. Likewise, Kupcinskas et al (2014) demonstrated that gene polymorphisms were not associated with the presence of high-risk atrophic gastritis (HRAG), GC, or different histological subtypes of GC, in European subjects.
In addition, the three other SNPs are also controversial. MicroRNA-34b/c gene rs4938723 polymorphism was associated with a decreased GC risk (Hua Li et al, 2017), but other studies did not support an association between the rs4938723 variant and digestive tract tumors, including GC (Hashemi et al, 2019). Sun et al (2010) demonstrated that a common polymorphism (rs895819) modulated miR-27a and ZBTB10 levels, with the variant genotypes (AG + GG) conferring a significantly increased GC risk relative to AA carriers, whereas the miR-608 rs4919510 variant G allele exhibited a significant association with an increased GC risk in a meta-analysis (Wu et al, 2017a); however, in Europe, gene polymorphisms of miR-27a T > C (rs895819) and miR-608 C > G (rs4919510) were not associated with the presence of HRAG, GC, or different histological subtypes of GC (Kupcinskas et al, 2014).
The inconsistency of the aforementioned conclusions may be mainly attributed to ethnic and racial differences. In addition, specific GC sites (cardia vs. noncardia), the histological subtypes (Ahn et al, 2013), molecular biological subtypes, the effect of classical gene mutations on these subtypes, such as TP53, HER2, CDH, and RAS, the heterogeneity of the cancer tissue, and the different sample sizes of the study, may also lead to research bias, or even contradictory conclusions. At the same time, different chemotherapy regimens, and the interactions between miRNAs, SNPs, and multiple miRNAs, can also have different effects on tumor prognosis and patient outcomes.
Related miR-SNPs involved in GC and their genetic information
An extensive search was performed through the literature and the dbSNP database, and the SNPs mentioned in this article were selected. The content of our study included functional regions of miRNA genes; other possible isoforms of miRNA genes; the location of SNP and allelic variants as listed in Table 1; the genotype and alelle frequencies of selected miRNA SNPs in different populations as listed in Table 2; and the selected miRNA genes with their target genes and suggested roles in GC cells as listed in Table 3.
Genotype Distribution of Investigated miRNA Single Nucleotide Polymorphisms in Gastric Cancer Patients and Controls in Different Populations
miR Involved in Gastric Cancer and Their Target Genes
Conclusion
miRNA-SNPs have stable inheritance, high abundance, and diversity within and among populations, and these factors play an important role in the transcription of pre-miRNA, miRNA processing, miRNA maturation, and mRNA interactions, all of which are prospective biomarker candidates for detecting disease. In this review, we not only focused on the biological function of GC based on SNP localization but also on susceptibility, tumor size, metastasis, overall survival, and prognosis, and uncovered the contradictions of previous studies. In addition, we selected some important and well-studied miRNA-SNPs associated with GC, for readers to consult in Table 3. We hope that this article will provide valuable information to researchers in this field.
However, the mRNA level of gene expression is often inconsistent with the protein levels, and the majority of these studies did not investigate the target at protein/functional levels. The different ethnicities and sample sizes, among other factors, may partly explain these discrepancies. Moreover, miRNA research lacks macroanalysis, the potential mechanism of specific regulation of miRNAs through different signaling pathways remains to be elucidated. Hence, a comprehensive screening is required for finding more evidence-based and disease-associated miRNA polymorphisms, to provide a new direction for the early monitoring of GC, the evaluation of therapeutic efficacy and prognosis, and the guidance of clinical individualization of patients.
Authors' Contribution
Concept; Design; Supervision - G.L.; Writing Article - L.P.; Data Collection - Y.S.; and Literature Search -J.Z.
Footnotes
Author Disclosure Statement
All authors declare that they have no competing interests, all authors confirm its accuracy.
Funding Information
No external funding was received for this article.