
Abstract
Background:
We aimed to evaluate the effects of lncRNA PTV1 on colon cancer proliferation and migration via the Wnt6/β-catenin2 pathway.
Materials and Methods:
A total of 117 colon cancer and normal adjacent tissue samples were collected. LncRNA PVT1 and miR-1207-5p expressions in these samples and colon cancer cell lines were detected by Quantitative reverse transcription-polymerase chain reaction (qRT-PCR). LncRNA PVT1-silencing cells and miR-1207-5p-overexpressing Caco-2-siPVT1 cells were constructed, respectively. The effects of lncRNA PVT1 silencing on cell proliferation were assessed by MTT and colony formation assays. The effects on invasion and migration were tested by Transwell and scratch assays respectively. The targeting regulatory relationship between miR-1207-5p and Wnt6 was analyzed by a dual-luciferase reporter assay. The relationship between lncRNA PVT1 and miR-1207-5p was studied by RNA-binding protein immunoprecipitation and RNA pull-down assays. The expressions of proteins in the Wnt6/β-catenin2 pathway were detected by Western blotting.
Results:
The lncRNA PVT1 mRNA expression in colon cancer tissue was significantly higher than that in normal adjacent tissue (p < 0.05). The expression in lncRNA PVT1-silencing cells was significantly down-regulated (p < 0.05). The colonies of Caco-2-siPVT1 cells decreased, accompanied by a reduced number of cells penetrating Matrigel and migration (p < 0.05). Compared with siPVT1 + NC group, the number of colonies and migration of siPVT1 + miR-1207-5p-overexpressing group increased significantly (p < 0.05). There was a targeting relationship between miR-1207-5p and PVT1. MiR-1207-5p had a targeted binding site with Wnt6. The protein expressions of Wnt6/β-catenin2 in Caco-2-siPVT1 group were significantly lower than those of control and Caco-2-siNC groups (p < 0.05).
Conclusion:
LncRNA PVT1 was highly expressed in colon cancer. It may enhance the proliferation and migration of colon cancer cells by up-regulating miR-1207-5p level and enhancing the Wnt6/β-catenin2 pathway.
Introduction
Colon cancer is one of the common malignant tumors of the digestive tract, with the incidence rate rising annually (Wang et al., 2017). Due to atypical symptoms in the early stage, most patients have entered the advanced stage upon diagnosis and the mortality rate is thus high. With continuous improvement of diagnosis and treatment methods, the outcomes of colon cancer therapy have been evidently improved, but the overall 5-year survival rate is still low (Chida et al., 2017; Nishida et al., 2017). Tumor invasion and metastasis are the main causes for death. At present, the mechanisms for colon cancer invasion and metastasis remain unclear.
LncRNA is a class of functional RNA molecules that do not encode proteins, with the transcription length of over 200 nt (Xu et al., 2019). LncRNA plays crucial regulatory roles in tumor cell proliferation, invasion, and metastasis (Munschauer et al., 2018). LncRNA PVT1 is specifically and highly expressed in tumor cells. Abnormal expression of PVT1 is associated with the onset of a variety of cancers and has been proved to promote tumor cell proliferation, migration, and invasion (Tang et al., 2018).
The distribution analysis of miRNAs in the human genome shows that they are usually produced in unstable gene groups, being closely related to the onset and progression of diseases (Zhu et al., 2017). Huppi et al. (2008) analyzed the desert region of 8q24 gene with frequent chromosome breakpoints. Through bioinformatics analysis and quantitative polymerase chain reaction (qPCR), they found that miR-1204, miR-1205, miR-1206, miR-1207-3p, miR-1207-5p, and miR-1208 were transcribed by PVT1 in this region. Besides, these six miRNAs were located in the intron of PVT1 gene. Currently, the role of PVT1 in colon cancer has seldom been reported. Therefore, we herein studied whether PVT1 regulated the expression of miR-1207-5p in colon cancer tissues and cell lines to participate in the onset and progression of this cancer. The results elucidate the regulatory relationship between lncRNA PVT1 and miR-1207-5p in its locus region and provide valuable evidence for targeted colon cancer therapy.
Materials and Methods
Clinical samples
This study has been approved by the Ethics Committee of our hospital and written informed consent has been obtained from all patients. The cancer tissues of 117 patients with colon cancer who underwent radical surgery in our hospital from July 2016 to July 2018 and normal tissues at >2 cm away from them were collected. All patients did not receive chemotherapy or radiotherapy before surgery.
Cells, reagents, and apparatus
Human normal colonic epithelial cells NCM460 and human colon cancer cell lines Caco-2, SW480, KM12, and HCT-8 were purchased from the American Type Culture Collection. PVT1 knockdown plasmid, lentiviral packaging plasmid, PVT1 primers, miR-1207-5p reverse transcription primers, real-time qPCR primers, and miR-1207-5p overexpression plasmid were all designed by Shanghai GenePharma Co., Ltd. (China). Dulbecco's modified Eagle's medium (D777) was bought from Applied Biosystems. TRIzol reagent and Transwell chambers were obtained from Thermo Fisher Scientific. MTT kit was provided by MsKBio (Germany). Quantitative reverse transcription-polymerase chain reaction (qRT-PCR) system was purchased from Applied Biosystems. Inverted fluorescence microscope was bought from MicroDemo.
Cell transfection
Caco-2 cells were inoculated into a 6-well plate at the concentration of 1 × 106/well, cultured overnight, and transfected with 100 nM siPVT1 and negative control using Lipofectamine 3000 transfection reagent, giving Caco-2-siPVT1 and Caco-2-siNC cells, respectively.
Detection of LncRNA PVT1 expression by qRT-PCR
The total mRNA of each group was extracted by the TRIzol method and reverse-transcribed into cDNA by using specific primers. The reaction conditions were set according to the instructions of SYBR® Premix Ex Taq kit: predenaturation at 95°C for 10 min, denaturation at 95°C for 30 s, annealing at 60°C for 10 s, and extension at 72°C for 10 s, 40 cycles in total; reaction at 95°C for 5 s, at 72°C for 1 min, and at 95°C for 30 s. LncRNA PVT1: upstream primer, 5′-GTCTCCCTATGGAATGTAAG-3′; downstream primer, 5′-AGTGTCCTGGCAGTAAAAG-3′. MiR-1207-5p: upstream primer, 5′-ACTGTCTCCCAACCCTTGTA-3′; downstream primer, 5′-GTGCAGGGTCCGAGGT-3′. The Ct value of each sample was calculated by software, and the relative expression of miRNA was represented as 2−ΔΔCT. Each experiment was repeated three times independently.
Detection of cell proliferation by MTT and colony formation assays
Caco-2-siPVT1, Caco-2-siNC, and NC cell lines were seeded into a 96-well plate at the density of 2000 per well, mixed and cultured in a 37°C incubator with 5% CO2. Then they were incubated with 20 μL of MTT solution each well at 37°C for 4 h in dark, and the supernatant was discarded. Subsequently, the cells were rapidly shaken with 100 μL of dimethyl sulfoxide for 15 min at 37°C to fully dissolve crystals. The optical density (OD) at 492 nm was measured by a microplate reader and recorded as OD492. Caco-2-siPVT1 and NC cells were cultured in a 6-well plate at the density of 500 per well, and the culture medium was refreshed every other week. After 2-3 weeks, the cells were stained with Coomassie blue, and the colonies were counted under a microscope.
Detection of cell invasion by Transwell assay
The cells were digested and washed with 10% phosphate-buffered saline (PBS) twice and added complete medium to prepare a single-cell suspension at the concentration of 1000 cells/mL. Afterward, 200-250 μL of the cell suspension was added to Transwell chambers. After incubation for 24 h, the cells were fixed in ethanol for 20 min, stained with 500 μL of 0.5% crystal violet staining solution at room temperature for 10 min in dark, rinsed with PBS, inverted and air-dried, and observed under an inverted fluorescence microscope for photographing and counting. Each experiment was performed in triplicate independently.
Detection of cell migration by scratch assay
Caco-2-siPVT1, Caco-2-siNC, and NC cells were seeded into a 6-well plate at 5 × 105/well and cultured overnight at 37°C with 5% CO2. Then scratches were made with a 10 μL pipette tip, and the cells were washed three times with PBS to rinse off the debris and added serum-free medium. The cells were thereafter photographed under the microscope and labeled, and the culture was continued. The cell migration was observed under the inverted microscope at 0, 24, 48, and 72 h, and photographed.
Verification of targeting regulatory relationship between miR-1207-5p and Wnt6 by dual-luciferase reporter assay
The regulatory relationship between them was studied by dual-luciferase reporter assay. Caco-2 cells were seeded onto 36-well plates and cultured at 37°C with 5% CO2 for 24 h. Then the cells were transfected with miR-1207-5p mimic and mimic control using Wnt6 wild-type plasmid and mutant plasmid, respectively, and cultured at 37°C with 5% CO2 for 24 h. Afterward, the cells were collected, and the luciferase activity was detected according to the instructions of dual-luciferase reporter assay kit (Promega).
Detection of binding of lncRNA PVT1 to miR-1207-5p by RNA pull-down assay
LncRNA PVT1 recombinant plasmid was digested, gel-recovered, mixed with transcription mixture, placed at room temperature for 3 h, added DNase I and left still at room temperature for 15 min. Then RNA was extracted, 4 μg of which was reacted at 90°C for 2 min, added the same volume of RNA-binding buffer and left still for 0.5 h. RNA and cell protein solution were mixed evenly. After 1 h, a small amount of background control was taken as the input sample, added 50 μL of magnetic beads, left still overnight at 4°C, washed three times, and added proteinase K buffer. After 30 min, the supernatant was collected to extract RNA which was then reverse-transcribed into cDNA for qPCR.
RNA-binding protein immunoprecipitation assay
RNA-binding protein immunoprecipitation assay was performed by using Magna RIP™ immunoprecipitation kit (Millipore). Caco-2 cells transfected with wild-type or mutant lncRNA PVT1 were lysed with a lysis solution containing protease and RNase inhibitors. The magnetic beads were incubated with Ago2/IgG antibody at room temperature for 30-60 min, and subjected to immunoprecipitation overnight at 4°C. After RNA-protein complexes were eluted from the magnetic beads, the expression of lncRNA PVT1 and miR-1207-5p was detected by RT-PCR.
Western blotting
Caco-2 cells after 48 h of transfection were added an appropriate amount of lysate, lysed for 30 min, and centrifuged at 12,000 rpm and 4°C for 10 min. The supernatant was collected, mixed with buffer, denatured in a water bath, and then subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Subsequently, the product was transferred onto a polyvinylidene fluoride membrane, incubated with primary antibodies against Wnt6 and β-catenin2 at 4°C overnight, washed with Tris buffered saline with Tween 20, incubated with secondary antibody for 2 h at 37°C, fully washed and developed with electrochemiluminescence reagent. Images were collected and analyzed by an automatic gel imaging system.
Statistical analysis
All data were analyzed by R3.5.1 and are expressed as mean ± standard deviation (
Results
LncRNA PVT1 expression in different tissues and cell lines
qRT-PCR showed that the mRNA expression levels of lncRNA PVT1 and miR-1207-5p in colon cancer tissue were significantly higher than those in normal adjacent tissue (p < 0.05) (Fig. 1A, C). Compared with normal colonic epithelial cell line NCM460, lncRNA PVT1 and miR-1207-5p expression was significantly higher in colon cancer cells, especially in Caco-2 cells (p < 0.05) (Fig. 1B, D). Therefore, Caco-2 cells were selected in subsequent experiments.

LncRNA PVT1 expression in
Verification of silencing results
qRT-PCR exhibited that the lncRNA PVT1 expression in Caco-2-siPVT1 cells was significantly downregulated compared with those in blank control and negative control groups (p < 0.05). The expression in the latter two groups was similar (p > 0.05) (Fig. 2). Therefore, PVT1-silencing and negative control cells had been successfully established.
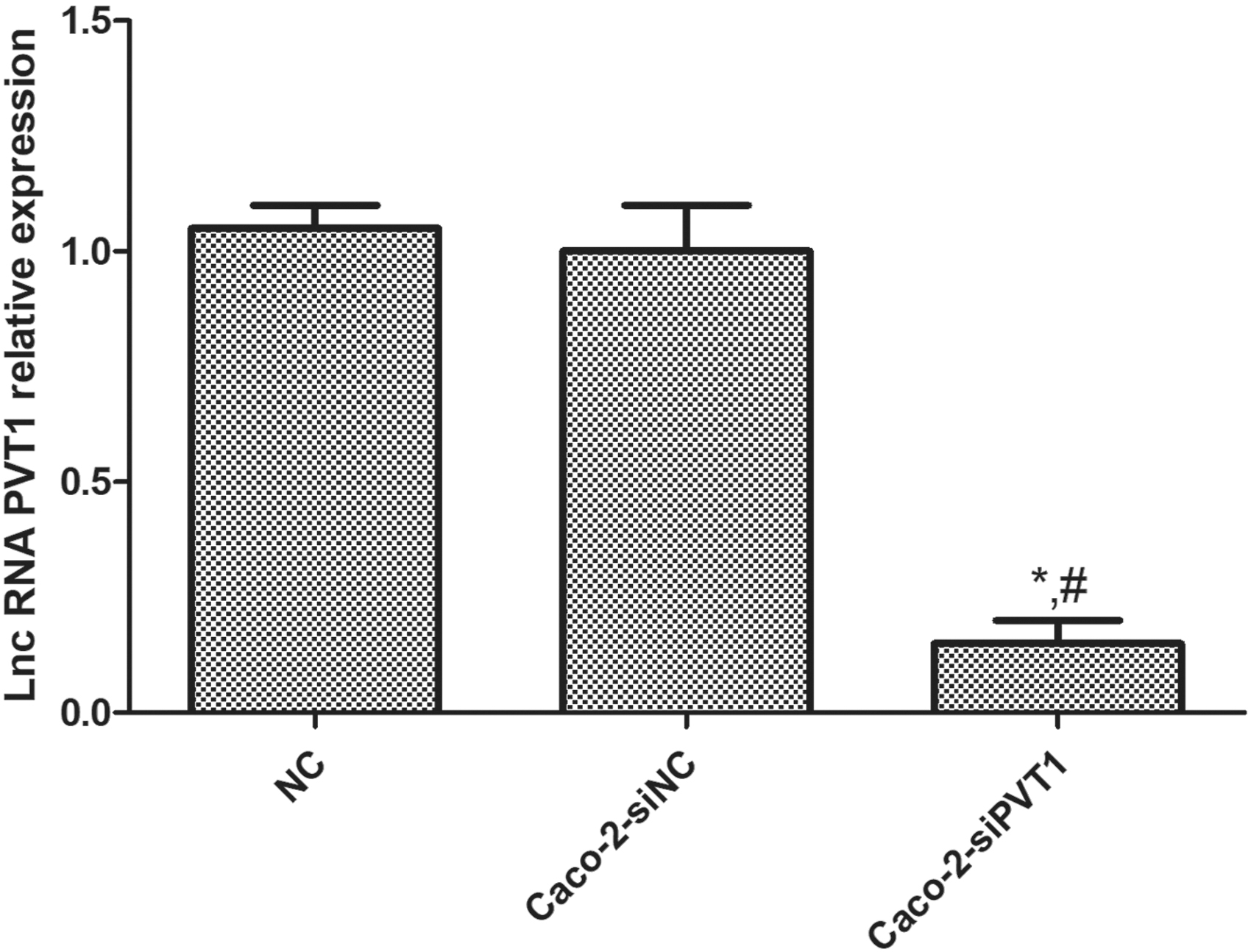
LncRNA PVT1 expression in different cells. qRT-PCR exhibited that the lncRNA PVT1 expression in Caco-2-siPVT1 cells was significantly downregulated compared with those in blank control and negative control groups. The expression in the latter two groups was similar. *Compared with blank control group, p < 0.05; #compared with Caco-2-siNC group, p < 0.05.
Effects of lncRNA PVT1 on Caco-2 cell proliferation
The MTT assay showed that OD492nm of the Caco-2-siPVT1 group was significantly lower than those of blank control and negative control cells (p < 0.05) (Fig. 3A). The colony formation assay exhibited that the colonies of Caco-2-siPVT1 cells significantly decreased (p < 0.05), but the colonies of blank control and negative control groups were similar (p > 0.05) (Fig. 3B). Collectively, the proliferation capacity of Caco-2 cells significantly decreased after lncRNA PVT1 was silenced.
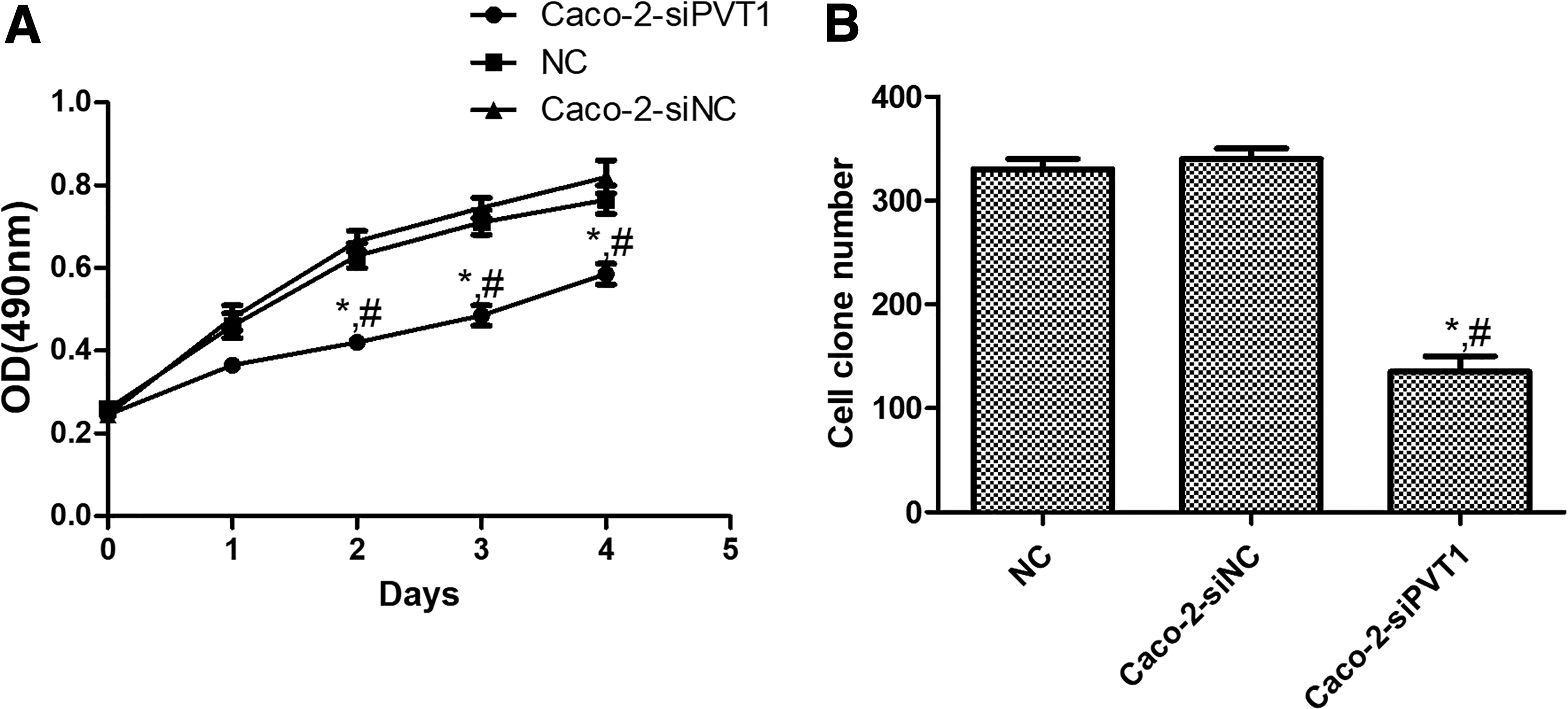
Effects of lncRNA PVT1 on Caco-2 cell proliferation.
Effects of lncRNA PVT1 on Caco-2 cell invasion
The Transwell assay revealed that blank control and negative control groups had similar numbers of cells penetrating Matrigel (p > 0.05) (Fig. 4A). The number of Caco-2-siPVT1 cells penetrating Matrigel significantly reduced (p < 0.05) (Fig. 4B). Thus, the invasion capacity of Caco-2 cells significantly decreased after lncRNA PVT1 expression was suppressed.
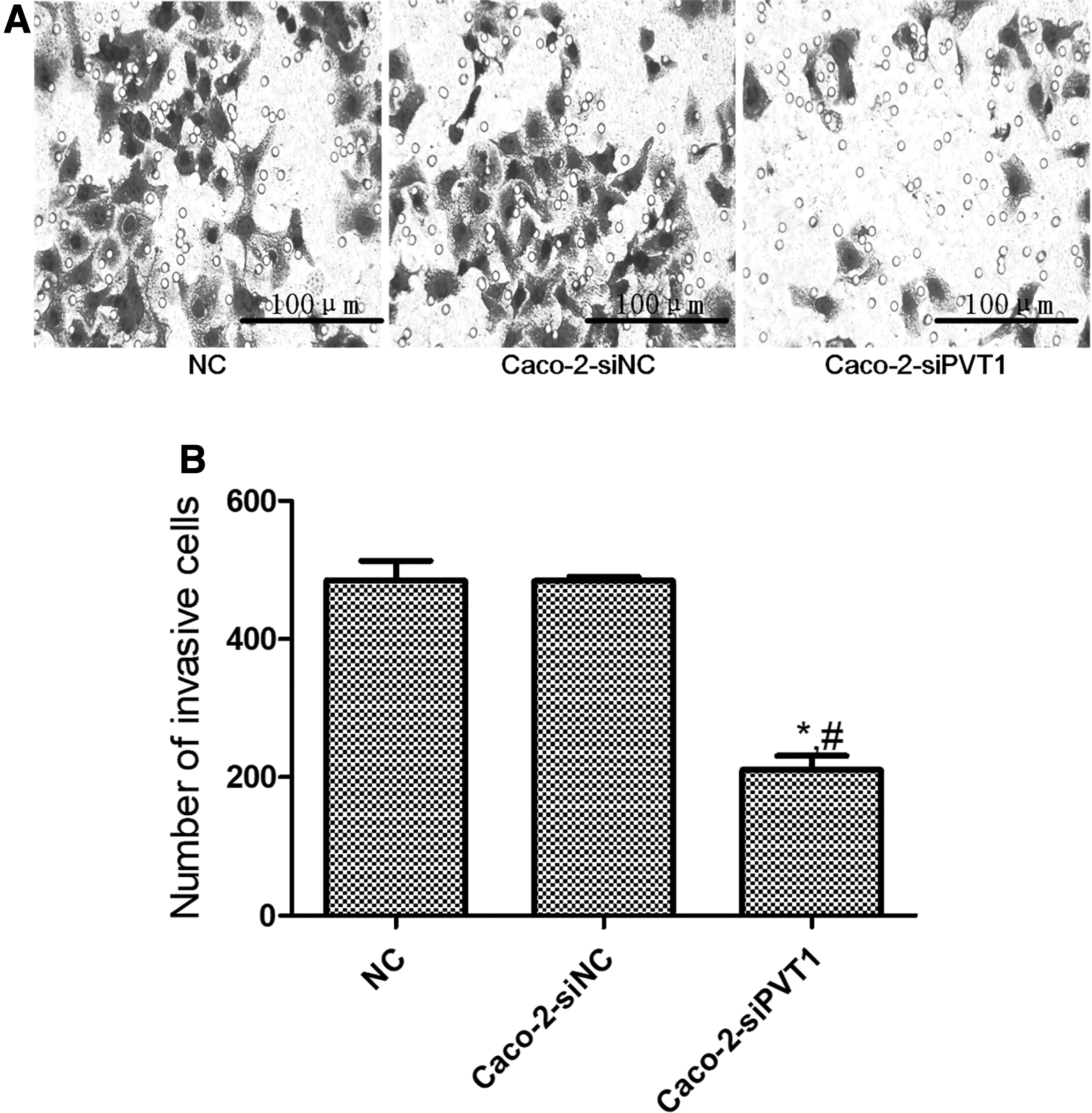
Effects of lncRNA PVT1 on Caco-2 cell invasion. After lncRNA PVT1 was silenced, the cell invasion ability significantly reduced.
Effects of lncRNA PVT1 on Caco-2 cell migration
The scratch assay showed that compared with blank control and Caco-2-siNC groups, the cell wound healing rate (Fig. 5A) and mobility (Fig. 5B) of Caco-2-siPVT1 cells significantly decreased (p < 0.05). There was no significant difference between the former two groups (p > 0.05). Accordingly, the cell migration ability was markedly attenuated after silencing of lncRNA PVT1.
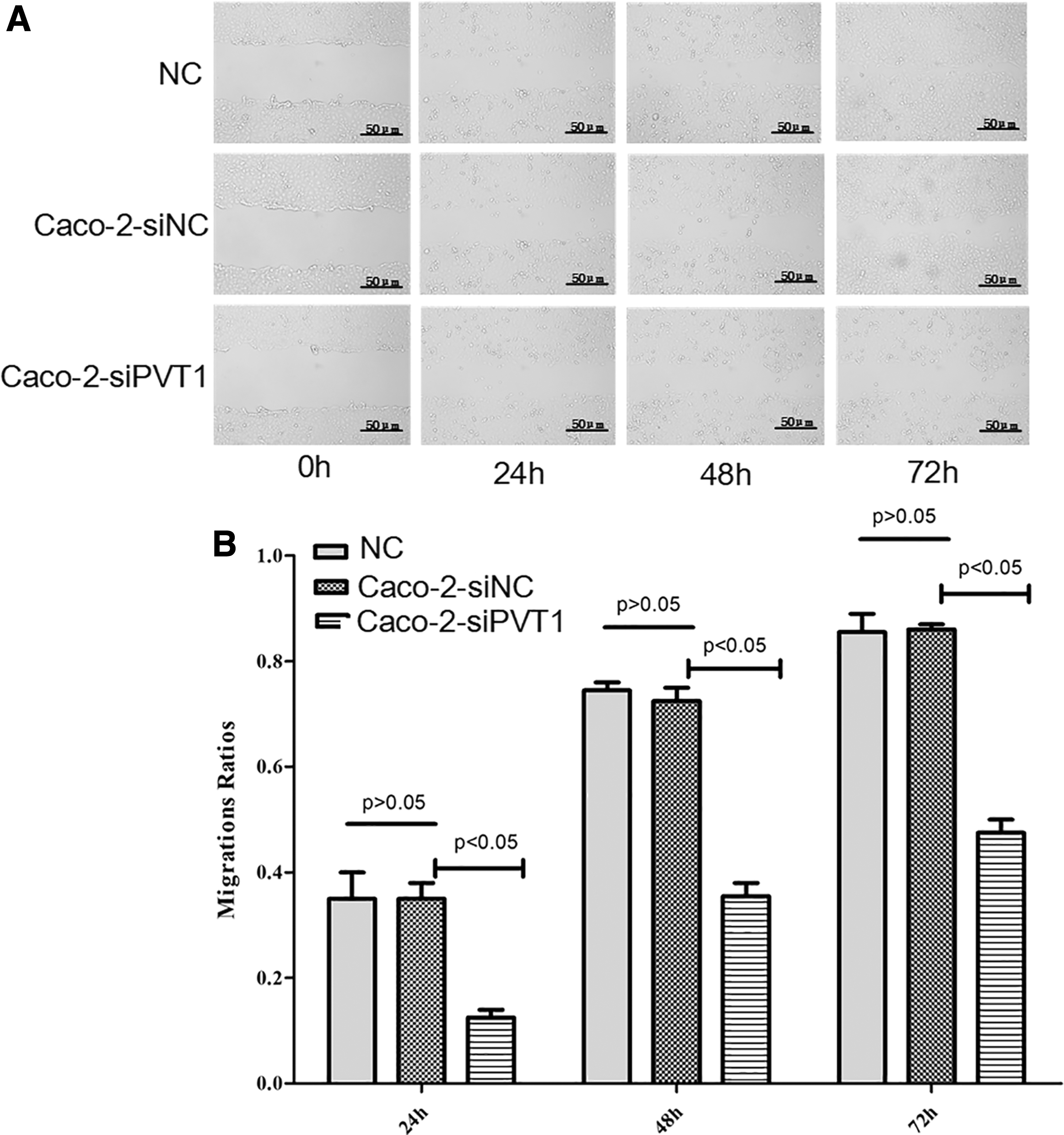
Effects of lncRNA PVT1 on Caco-2 cell migration. After lncRNA PVT1 was silenced, the cell migration ability significantly reduced.
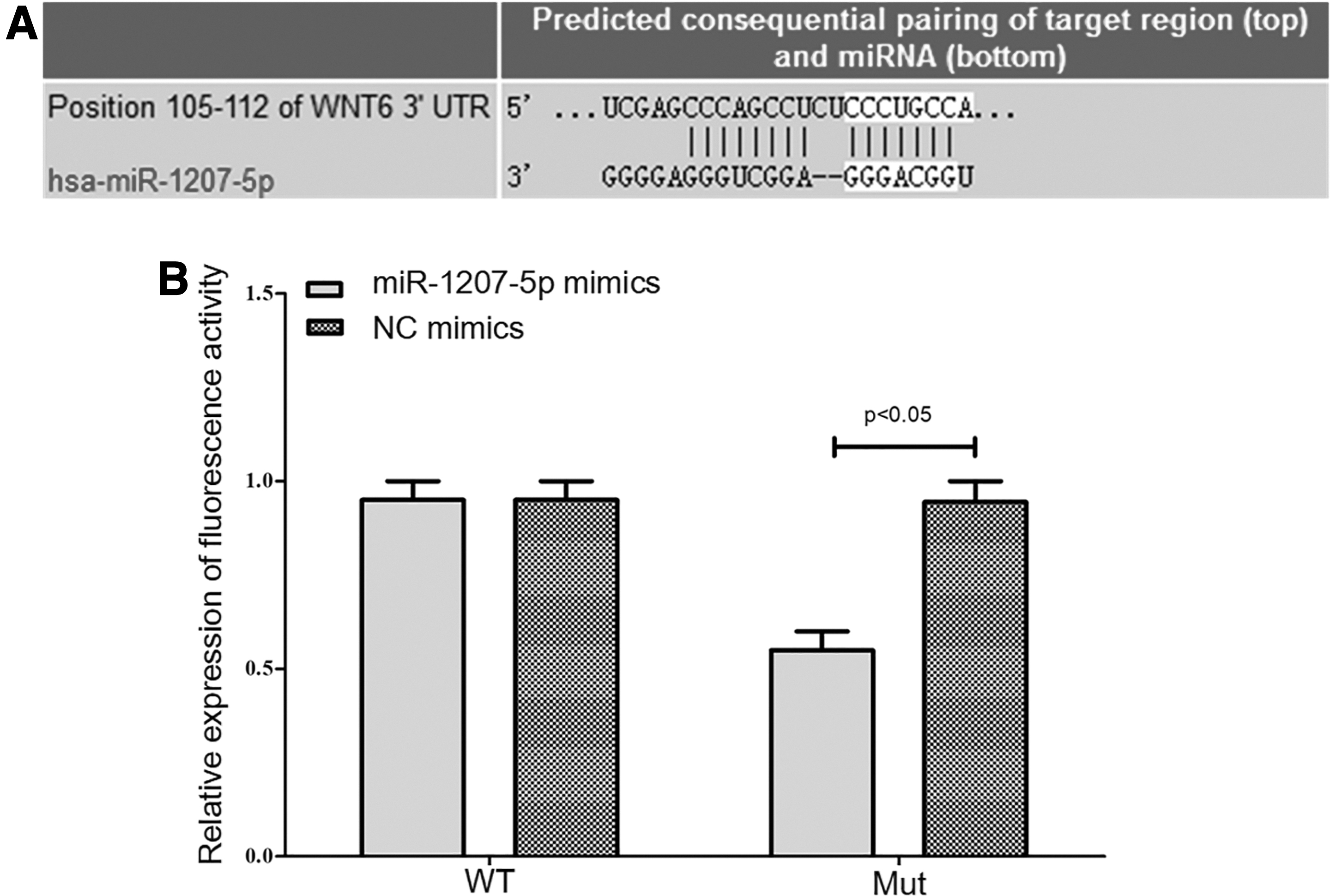
Targeted regulatory effects of miR-1207-5p on Wnt6
TargetScan showed that miR-1207-5p had a targeted binding site with Wnt6. After transfection, the luciferase activity of wild-type miR-1207-5p was significantly inhibited (p < 0.05), but that of mutant miR-1207-5p showed no significant change (p > 0.05) (Fig. 6).
Relationship between PVT1 and miR-1207-5p.
LncRNA PVT1 worked as an endogenous competitor RNA for miR-1207-5p
After adsorbing miRNAs, lncRNAs inhibit their expression and reduce the regulatory effects on target mRNAs. Therefore, we further investigated whether lncRNA PVT1 worked through an endogenous competitor RNA mechanism. The target recognition sequences of miRNAs by lncRNA PVT1, which were predicted by bioinformatics analysis (RegRNA2.0, http://regrna2.mbc.nctu.edu.tw/detection.html; Lncrnadb, www.lncrnadb.org; miRcode, www.mircode.org/), showed that miR-1207-5p had sequences complementary to those of lncRNA PVT1 (Fig. 7A).
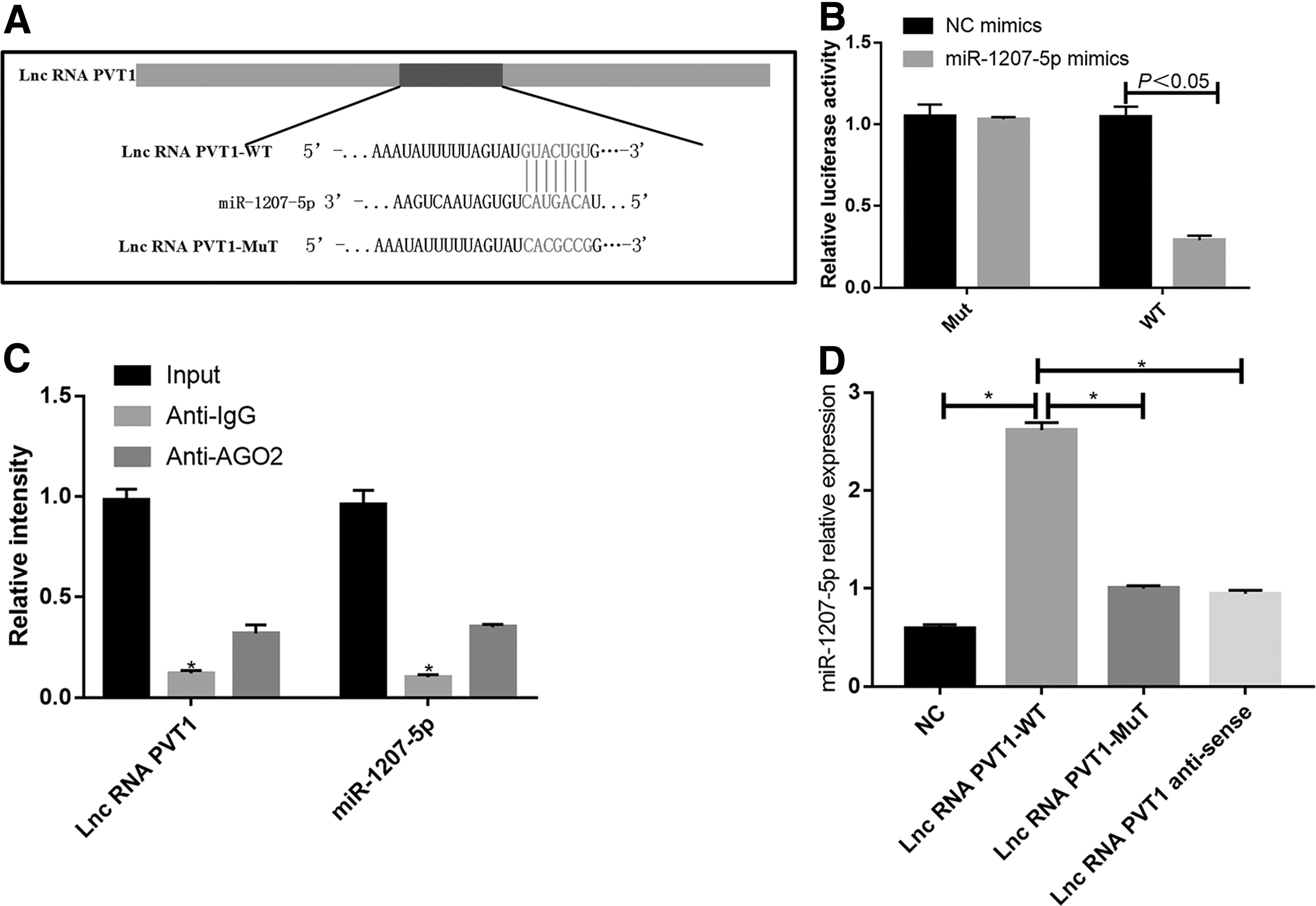
LncRNA PVT1 worked as an endogenous competitor RNA for miR-1207-5p.
To validate the above finding, lncRNA PVT1 cDNA was cloned into luciferase gene (pGL3-wt) and cotransfected with miR-1207-5p or miR-NC. The luciferase activity of the miR-1207-5p group was significantly lower than that of the miR-NC group. Meanwhile, the binding site of miR-1207-5p was mutated to construct a pGL3-Mut vector. However, the mutated vector hardly affected the luciferase activity of the miR-135-5p group (Fig. 7B).
RNA-induced silencing complex, which consists of microRNA-containing ribonucleoprotein complexes (miRNPs), exists in anti-Ago2 immunoprecipitates. Thus, anti-Ago2 immunoprecipitates contain miRNAs and their interacting RNA components. Compared with anti-IgG immunoprecipitate, lncRNAs PVT1 and miR-1207-5p were preferentially enriched in AGO2-containing miRNP (Fig. 7C). The results of the RNA pull-down assay showed that lncRNA PVT1 was more significantly enriched in wild-type miR-1207-5p than in mutant miR-1207-5p at the binding sites (Fig. 7D).
Validation of effects of lncRNA PVT1 on Caco-2 cells by overexpressing miR-1207-5p
After lncRNA PVT1 was silenced, miR-1207-5p was overexpressed to observe the proliferation and migration of Caco-2 cells. Compared with the siPVT1+siNC group, the number of colonies and migration of the siPVT1+miR-1207-5p-overexpressing group increased significantly (p < 0.05) (Fig. 8). Hence, overexpressing miR-1207-5p reversed the influence of lncRNA PVT1 silencing on Caco-2 cells.
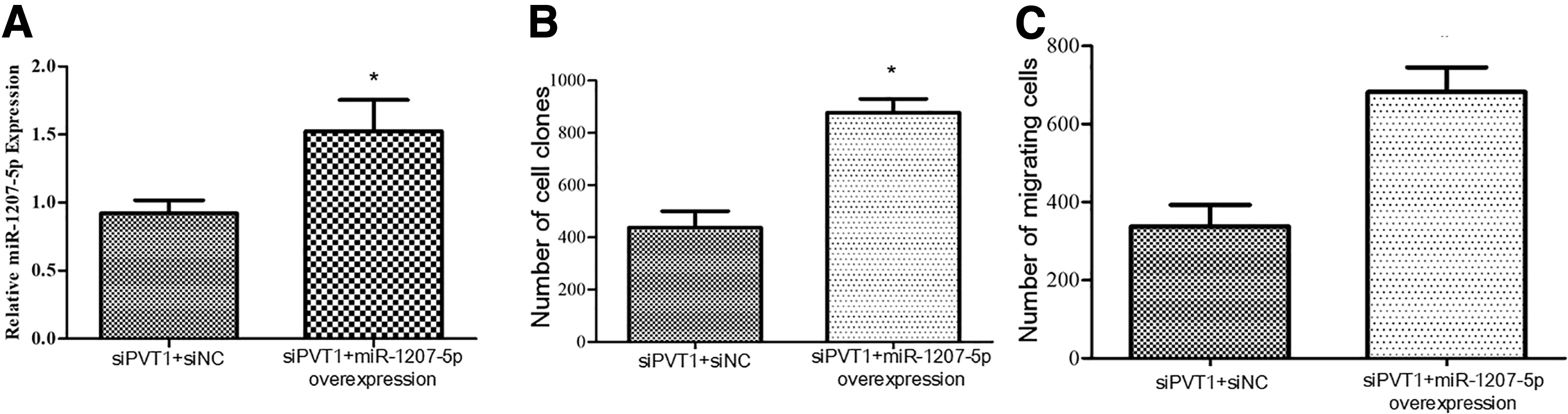
Validation of effects of lncRNA PVT1 on Caco-2 cells by overexpressing miR-1207-5p. Compared with control group, *p < 0.05.
Relationship between lncRNA PVT1 and Wnt6/β-catenin2
Western blot exhibited that the protein expression of Wnt6/β-catenin2 in the Caco-2-siPVT1 group was significantly lower than those of control and Caco-2-siNC groups (p < 0.05), but the former two groups had similar expression (p > 0.05) (Fig. 9). Therefore, Caco-2-siPVT1 promoted protein expression in the Wnt6/β-catenin2 pathway.
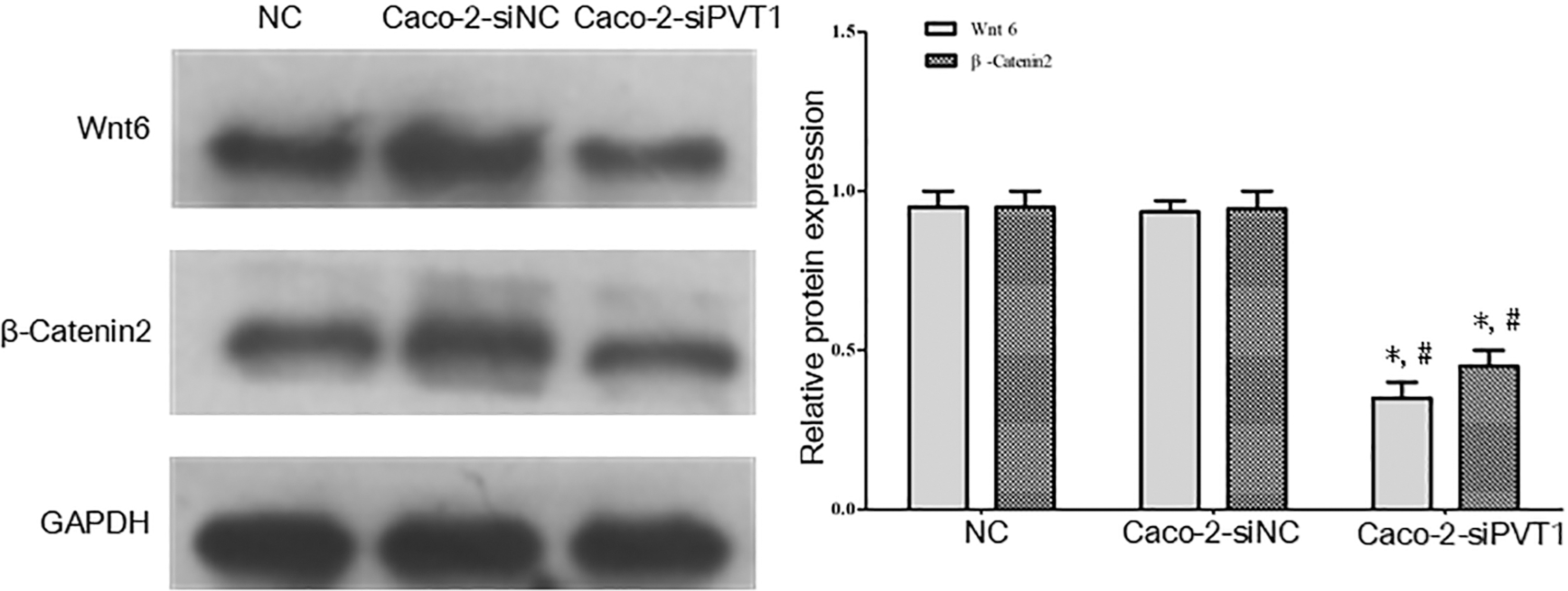
Relationship between lncRNA PVT1 and Wnt6/β-catenin2. Western blot exhibited that the protein expression of Wnt6/β-catenin2 in the Caco-2-siPVT1 group was significantly lower than those of control and Caco-2-siNC groups, but the former two groups had similar expression. *Compared with blank control group, p < 0.05; #compared with Caco-2-siNC group, p < 0.05.
Discussion
Colon cancer is one of the common digestive tract malignancies, with a morbidity rate ranking the third among all the malignant tumors, and the tumor invasion and metastasis are the leading causes of death of patients (Muro, 2017; Shimokawa et al., 2017). Currently, the mechanisms of invasion and metastasis of colon cancer remain unclear. As a category of small noncoding RNAs, miRNAs can inhibit the protein transcription of target genes or degrade target genes. Although the action mechanism and regulatory mechanism of such molecules have not been clarified, yet, there is growing evidence that miRNAs have significant correlations with diseases as well as potential values for the prediction, diagnosis, and treatment of diseases (Li et al., 2017a, 2017b; Shi and Zhang, 2017). Hence, investigating the functions and regulatory mechanisms of abnormally expressed miRNAs in colon cancer can help understand the disease more comprehensively.
LncRNAs play pivotal regulatory roles in the proliferation, invasion, and metastasis of tumor cells, so the research on lncRNAs has been deepened gradually in recent years. LncRNA PVT1 is a type of lncRNA that is specifically highly expressed in tumor cells, which has been proved to be able to influence the proliferation, migration, and invasion of various tumors (Das et al., 2014). Existing studies have demonstrated that PVT1 gene, located on chromosome 8q24 and spanning 300 kD bases on the genome, can produce more than 20 transcripts and 6 clusters of miRNAs (Zhao et al., 2018). Sun et al. (2015) found that lncRNA PVT1 can regulate the expression of miRNAs through common promoters, thus intervening in the proliferative ability of breast cancer cells and further affecting the occurrence and development of the disease. Therefore, lncRNA PVT1 probably promotes the migration and proliferation of ovarian cancer cells.
In this study, the expression of lncRNA PVT1 in colon cancer tissues and paracarcinoma tissues was detected via qRT-PCR, and the results indicated that the expression level of lncRNA PVT1 in colon cancer tissues was remarkably higher than that in normal paracarcinoma tissues, confirming the inference that lncRNA PVT1 may be a crucial player in the occurrence and development of colon cancer. To explore the action mechanism of lncRNA PVT1 in colon cancer, the cell lines with silenced PVT1 were constructed in this study. The results illustrated that the proliferative, invasive, and migratory abilities of colon cancer Caco-2 cells were weakened obviously after lncRNA PVT1 was downregulated.
MiRNAs are endogenous small single-stranded ncRNAs, which can conjugate with target genes by means of incomplete complementary base pairing, so as to influence the occurrence and development of multiple malignant tumors (Song and Yin, 2019). In this study, it was predicted by TargetScan database that miR-1207-5p had a binding site to PVT1, and it was revealed in dual-luciferase reporter assay that there was a targeted regulatory relationship between miR-1207-5p and PVT1. Chen et al. (2014) discovered through research that the expression of miR-1207-5p in breast cancer tissues is higher than that in normal paracarcinoma tissues, the upregulated expression of miR-1207-5p can facilitate the proliferation of breast cancer cells, and the abnormality of miR-1207-5p expression directly affects the expression level of target genes.
Herein, the expression of miR-1207-5p was downregulated after silencing lncRNA PVT1 in Caco-2 cells, suggesting that the role of lncRNA PVT1 in promoting the proliferation and migration of colon cancer Caco-2 cells may be associated with the upregulation of miR-1207-5p. Yan et al. (2017) found that miR-1207-5p acted as an oncogene to accelerate the progression of breast cancer. In this study, overexpressed miR-1207-5p was able to reverse the biological behaviors of Caco-2 cell lines following lncRNA PVT1 silencing, implying that the promoting effect of lncRNA PVT1 on the proliferation of Caco-2 cells may be related to the upregulation of miR-1207-5p. It is well documented that the expression of miRNA is affected by the level of its host gene expression (Gao et al., 2012). The expression of most intronic miRNAs is consistent with their host gene expression, being controlled by the host gene promoters (Lutter et al., 2010).
In addition, E2 can enhance the activity of PVT1 promoter and upregulate its expression by upregulating the expression of MYC protein and by binding E-box in the PVT1 promoter region (Carramusa et al., 2007; Wang et al., 2011). In this study, PVT1 positively regulated miR-1207-5p in colon cancer, so we postulated that PVT1 may regulate the expression of miR-1207-5p through a copromoter, thereby affecting the progression of colon cancer.
Based on the prediction of TargetScan website, miR-1207-5p could complementarily bind to Wnt6/β-catenin2. The Wnt6/β-catenin2 signaling pathway plays an important role in the occurrence and development of tumors, and it transmits pathway signals in cells by phosphorylating transcription factors (Russell and Monga, 2018). Herein, the phosphorylation level of β-catenin in cells declined markedly in Caco-2-siPVT1 group. Thus, lncRNA PVT1 may influence the progression of cancers by modulating the Wnt6/β-catenin2 signaling pathway.
In summary, lncRNA PVT1 is highly expressed in colon cancer, and the highly expressed lncRNA PVT1 may elevate the level of miR-1207-5p to enhance the Wnt6/β-catenin2 signaling pathway, thereby facilitating the proliferation and migration of colon cancer cells. Accordingly, taking lncRNA PVT1 as a new research direction can provide certain theoretical supports for novel biological drugs.
Footnotes
Acknowledgment
We thank all coauthors for their significant contributions.
Authors' Contributions
P.Y.: Conceptualization (supporting), writing—original draft (lead), and formal analysis (lead). J.Z.: Methodology (lead) and formal analysis (supporting). A.Z.: software (lead) and formal analysis (supporting). W.K.: methodology (supporting) and writing—original draft (supporting). X.S.: conceptualization (lead) and writing—review and editing (lead). All authors have approved the submission and publication of this article.
Ethics Committee Approval
This study has been approved by the Ethics Committee of Affiliated Hangzhou First People's Hospital, Zhejiang University School of Medicine.
Informed Consent
Written informed consent has been obtained from all patients.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.