
Abstract
Abstract
Zhang, Jiaxing, Haichen Liu, Xiaodan Yan, and Xuchu Weng. Minimal effects on human memory follwing long-term living at moderate altitude. High Alt. Med. Biol. 12:37–43, 2011.—A number of studies describe memory deficits at extremely high altitudes. However, little is known about the effect of long-term living at moderate altitude (MA). The subjects for this study were 52 college students originally from sea level (SL), but studying at a MA of 2260 m over a 7-month period, with a return to SL for 30 days in the middle of the period. Fifty-two matched college students who stayed at SL all the time were the control group. The neuropsychological battery of assessments included the Chinese revised version of Wechsler Memory Scale tests (WMS-CR), verbal and spatial two-back working memory tests, long-term explicit memory (word recall and recognition of words, faces, and pictures) tests, Rey Auditory Verbal Learning test, Rey–Osterrieth Complex Figure (ROCF) test, degraded picture naming test, and the Serial Reaction Time Test. We found that the MA subjects showed significantly poorer performances than SL controls only in short-term visual construction assessed in the visual reproduction test from WMS-CR and in the ROCF immediate test. There were no significant differences in all other tasks between the MA group and SL group. These findings suggest that long-term hypoxic exposure at moderate altitude has minimal effects on human memory.
Introduction
Population density at elevations near 2300 m exceeds that at 200 to 300 m and approaches that found below 100 m (Cohen and Small, 1998). In Mexico (Vazquez, 2000) and western China, many cities are located between 2000 and 2500 m above sea level (SL). Moreover, every year hundreds of thousands of people travel from the lowlands to such moderate altitude (MA) locations to work, study, or train for long periods of time (Levine and Stray-Gundersen, 1997; Cohen and Small, 1998; Vazquez, 2000; Stray-Gundersen et al., 2001). However, up to now, the effect of long-term MA living on cognitive functions remains unclear.
Our previous studies on animals have shown that long-term exposure to a simulated MA of 2000 m had an enduring effect on memory, and this effect was related to the development stage when animals were exposed to hypoxia (Zhang et al., 2005; Zhang et al., 2006). We therefore hypothesized that MA exposure could have either acute or chronic effects on the cognition of human who resided at MA for a long time.
A large number of students come to Qinghai–Tibet Plateau to attend universities every year. The young college students are in their later developmental period; thus whether their cognitive functions would suffer from the MA environment may be of most concern. Therefore, we assessed the cognitive functions of college students at Xining (located on the northeastern part of the Qinghai–Tibetan Plateau (height 2260 m, Pa
Materials and Methods
Subjects
We recruited 52 students from Qinghai Normal University and Qinghai University at Xining for cognitive tests. All were lowlanders, born and living at SL below 300 m,. without any prior exposure to altitude higher than 500 m. They ascended to Xining for college study. Fifty-two students, who were born and raised at SL, were recruited as the SL control group from the China Agriculture University at Beijing (<300 m) and Southwest University for Nationalities at Chengdu (<300 m). Each group consisted of 24 males and 28 females aged 17 to 20 yr. The MA and SL students did not differ on IQ or college enrollment scores, and all were selected from the Han ethnic group to avoid possible racial differences. Subjects were examined physiologically (including respiration, pulse, blood pressure, and hematological measurements) before cognitive assessment and were excluded if they had (1) a documented neurological disorder or (2) a past history of head injury with loss of consciousness.
All subjects were tested after they had attended college for 7 months, with a 1-month vacation in the middle. During the vacation, MA subjects returned to SL, and therefore the MA subjects had hypoxic exposure of two periods of around 80 days, with 30 days in the middle spent at SL. All subjects were tested on their own campus. In both groups, prior to cognitive assessment, we conducted physiological tests and collected information such as body mass index, sleep quality, and mood disorders. The Research Ethics Review Board of the Institute of Psychology, Chinese Academy of Sciences, approved all experimental protocols. We assured that all subjects understood the experimental procedure, and each provided written informed consent before participating in the study.
Neuropsychological tests
All subjects were given the following standardized neuropsychological tests:
The Chinese revised version of Wechsler Memory Scale (WMS-CR) (Gong, 1989) provided measures of new verbal learning and visual and verbal memory functions. Verbal and spatial two-back tasks were used to assess working memory (D'Esposito et al., 1998). Words recalled and recognition of words, faces, and pictures provided measures of long-term explicit memory functions. Degraded Picture Naming was used to measure implicit memory, and a Serial Reaction Time Task (SRTT; Nissen and Bullemer, 1987) was used to measure simple visuomotor implicit procedural learning. The Rey–Osterrieth Complex Figure (ROC; Rey, 1941; Watanabe et al., 2005) assessed the short- and long-term visual memory and visuoconstructional ability. The Rey Auditory Verbal Learning Test (RAVLT) (Schmidt, 1996; Mattos et al., 2003) was used to assess short- and long-term verbal memory.
Experienced psychologists scored all the tests using the scoring rules detailed in the test manuals. The visual stimuli in working memory, words recall, degraded picture naming, and SRTT, as well as words, faces, and pictures recognition tests, were presented to the subjects in black font on a white screen on a computer with a PHILIPS 107S6 17" flat CRT monitor (Philips Inc., Amsterdam, The Netherlands), and subjects were instructed to respond on a standard keyboard.
Verbal and spatial two-back working memory
In the verbal working memory task, the letters A, B, C, D, E, F, G, and H were presented in a programmed sequence for 2 sec. Subjects were instructed to determine whether each presented letter was the same as the one presented two stimuli previously. In the spatial working memory task, a square was presented for 2 sec in one of eight different locations around an imaginary square centered on a fixation cross. Subjects were instructed to decide whether each presented square was in the same position as the square shown two stimuli previously. In each test, there were 84 trials, with a ratio of targets to distracters of approximately 1:4.
Word free recall
Thirty emotionally neutral Chinese nouns (e.g., home, tree, sky) were presented centrally for 2 sec each in the central visual field, followed by a fixation cross for 500 ms. Five minutes after this encoding period, the subjects were asked to recall the words they had seen during the presentation. Words were selected from the Modern Chinese Frequency Dictionary (1986) with a mean frequency of 0.03290% (0.00641% to 0.27836%). Each was unrelated to the others in semantics, phonology, or orthography. The score for this test was the number of correct words counted.
Word, face, and picture recognition
These tests investigated memories for words, faces, or pictures by consecutively presenting 25 target words, faces, or pictures on the screen with the same interval of 2 sec. After the presentation, 25 target words, faces, or pictures were mixed with another 25 new words, faces, or pictures, and the subjects were required to recognize the targets they had seen in a previous session. All 50 words were selected from the Modern Chinese Frequency Dictionary with a mean frequency of 0.03352% (0.00542%-0.29799%). Each was unrelated to the others in semantics, phonology, or orthography. The 50 faces consisted of 24 happy and 26 sad masked faces selected from the International Affective Picture System (Lang, 1995). The 50 pictures were asymmetric, gray-scale, line drawings from a published database (Shu et al., 1989). Pictures (3.2 cm wide by 3.2 cm high) depicted a variety of objects, animals, and people (e.g., ladder, horse, nurse).
Degraded picture recognition
The 50 pictures used in the picture recognition test were degraded with homemade software. Immediately following completion of the picture recognition test, degraded pictures were presented centrally for 2 sec, followed by a 5-sec response interval. This gave the subject ample time to make a discrimination. Pictures were also selected from a database (Shu et al., 1989). The pictures used in the picture recognition test were not used in this test.
SRTT
We employed a version of the Nissen and Bullemer SRTT (1987) based on the paradigm developed by Gomez Beldarrain and colleagues (2002). Subjects were seated in front of a computer screen with their middle and index fingers of their left and right hands resting on a keyboard over keys 1, 2, 3, and 4, respectively. An asterisk was presented in any one of four positions horizontally located on the screen aligned above the response keys. Subjects were instructed to press the key that was spatially aligned with the asterisk as fast as possible with the corresponding finger. The asterisk did not disappear until the correct button was pressed, after which the next stimulus appeared after a delay of 500 ms. The test consisted of six blocks with 120 trials in each block. In blocks 1 and 6, the asterisk positions randomly presented at each of the four positions, in blocks 2 to 5, a fixed 12-trial sequence (3-4-1-4-3-4-2-4-1-3-4-2) was repeated 10 times. Reaction time (RT) was measured for each presentation. None of the subjects detected the sequence.
ROCF
Subjects were presented with a complex figure from the ROCF set (maximum score = 36) on a card measuring 21 by 29.7 cm and asked to copy it. The ROCF was drawn with pens in four colors, which were changed in a fixed order to enable the examiner to track the drawing sequence. Immediately following completion of the copy trial, the figure was removed and subjects were asked to reproduce the figure from memory (testing for immediate recall). Approximately 20 min after the last exposure to the figure, subjects were asked to reproduce the figure from memory again (a test for delayed recall). Two scores were derived from the immediate recall and delayed recall.
RAVLT
This test was based on the one previously developed by Mattos and colleagues (2003). The test consists of two lists (A and B) of 15 words (translated into Chinese), each arranged in a fixed order. List A was read aloud by the examiner (trial 1, T1) and immediately the subjects were asked to recall as many words as they could. The examiner then repeated the same procedure four more times (T2 to T5). A normal performanc showed increasing recall of words during the test, usually referred to as a learning curve. After the fifth recall of list A (T5), list B was read aloud only once, and the subjects were also instructed to recall as much as possible. List B was actually a distracter to prevent rehearsal of the material being held in short-term memory (T6). The examiner then asked the subject to recall list B. Twenty minutes later, another free recall of list A was demanded (T7). Scoring was based on the number of correct responses. Recalling a word not presented in list A or B was counted as an error. In the current study, the RAVLT scores assessed different aspects of memory. T1 measured immediate memory span and attention, T5 was a measure of best learning (in most cases this represents the maximum number of words recalled on a trial), the total score represented the incremental learning of words across the five trials, and delayed recall (T7) assessed storage and retrieval.
Data analysis
All data were analyzed using SPSS (Version 11.0, Chicago, IL, USA) software. The ANOVA statistic identified the differences between the MA and SL groups. Data were presented as mean (±SD). Statistical significance was set at p < 0.05.
Results
Physiological characteristics
Physiological tests (Table 1) were conducted 1 day before cognitive tests. One-way ANOVA showed that hemoglobin levels and circulating red blood cell count in MA males were both significantly higher than those in SL males (both p < 0.001). The two measurements did not show significant differences among females (p > 0.05). Our results from MA males agreed with those of Heinicke and colleagues (2005), who observed increases in red blood cell volume and total hemoglobin mass in athletes trained at 2050 m for 3 weeks. Average body weight of both MA males and females was lower than that of SL control males and females (both p < 0.05).
MA, midde altitude; SL, sea level; HGB, hemoglobin; RBC, red blood cells. Data are represented as mean (SD).
p < 0.001.
p < 0.05; subjects at MA vs. subjects at SL.
Cognitive tests
The test scores of the MA group and SL control group for WMS-CR are shown in Table 2. The scores of long-term explicit memory (words recall and words, faces, and pictures recognition) tests, degraded picture naming test, verbal and spatial two-back working memory tests, and ROCF test are shown in Table 3. One-way ANOVA only detected significantly poorer performances in the visual reproduction task from WMS-CR (p < 0.05) and in the ROCF immediate recall test (p < 0.05) in the MA group than in the SL group. No significant differences were found in the other tests between the MA and SL groups.
SRTT
The RTs across blocks 1 to 6 of the SRTT in theMA and SL groups are shown in Fig. 1. Group (MA vs. SL) was taken as the variable between subjects, while block (2 vs. 3 vs. 4 vs. 5) was taken as the variable within subjects. Two-way ANOVA with repeated measures (four levels, blocks 2 to 5) did not detect a significant difference between MA and SL group RT values over the entire trial of four blocks (p > 0.05), with a significant effect of block within both groups (both p < 0.001). One-way ANOVA revealed a significant difference in RT values between the random block (block 6) and the last sequential block (block 5) in both the SL (p < 0.01) and MA (p < 0.001) groups. However, there was no significant difference between the MA and SL groups in the rebound increase in the RT values between blocks 5 and 6 (96.3 vs. 70.2 ms) (p > 0.05). Therefore, both task-related (the shortening of the RT from blocks 2 to 5) and sequence-specific (the rebound RT increase between blocks 5 and 6) learning occurred in both groups. However, there were no significant group difference between MA and SL.

Response time (RT) across blocks 1—6 of the Serial Reaction TIme Task in altitude and sea level groups. Note the progressive shortening of RT during sequential blocks (1—5) and the rebound increase in RT in block 6 (random). Note also that MA showed no significant difference in rebound from SL, suggesting no changed sequence.
RAVLT
Figure 2 illustrates the words learning curve and free recall. Group (MA vs. SL) was taken as the variable between subjects, while block (II vs. III vs. IV vs. V) was taken as the variable within subjects. Two-way ANOVA with repeated measures (four levels of each variable, blocks II, III, IV, V) did not detect a significant difference between MA and SL groups in the number of words correctly recalled over the entire trial of four blocks (p > 0.05), with a significant effect of block within both groups (both p < 0.001). One-way ANOVA revealed no significant group differences between MA and SL in the scores of T1, T5, delayed recall (T7), or total score (all p > 0.05).
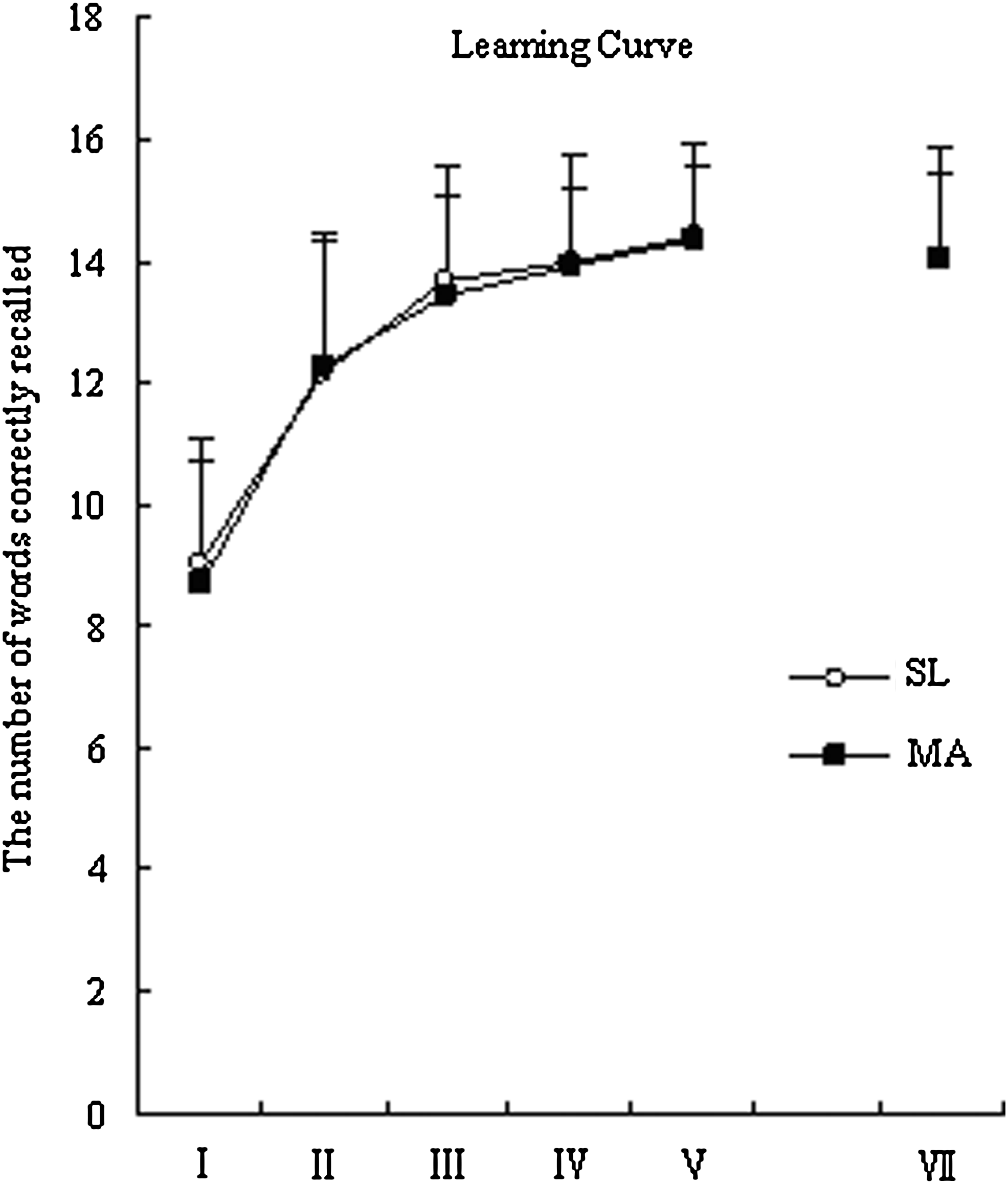
The learning curve and a free recall. The two groups showed similar incremental learning patterns from trial I to trial V. Two-way ANOVA did not reveal a significant difference between MA and SL groups, and T test fails to show any differences in T1, T5, T7, and total score.
Discussion
The present study is the first investigation of the effects of long-term living at MA on human cognition. We tested subjects with a variety of tasks, which included both short- and long-term memory, covering from explicit to implicit and from visual to auditory memory. However, we found that subjects who lived at MA for a long time only performed significantly poorer than the controls at SL in a small number of tests. Moreover, the poor performances were confined to short-term visual construction assessed in the visual reproduction test from the WMS-CR and in the ROCF immediate test.
Previous research mainly focused on the effect of higher altitude exposure on human cognition. In an acute study, Bakharev (1981) reported that performances in a variety of memory tasks were considerably worse during the first day at 3400 to 3600 m. Most observations were conducted based on long-term studies. For example, McFarland (1938) found significant impairments in arithmetic performance, word recognition, attention level, and short-term memory among high altitude expeditioners at altitudes up to 5400 m. Nelson (1982) reported impaired spatial memory in mountaineers at altitudes greater than 5000 m during a 35-day mountaineering expedition to Denali. Hornbein and colleagues (1989) found a decline in both visual and verbal long-term memory in mountaineers from 1 to 30 days following ascent to altitudes between 5488 and 8848 m. Kramer and colleagues (1993) recorded reduced learning and retention in perceptual and short-term memory compared with controls 2 weeks after an ascent to Mount Denali (6194 m). Lieberman (2005) tested Mount Everest climbers soon after they came back to SL and found impaired language comprehension and the ability to change plans. Taken together, these studies suggested short-term memory and visual spatial memory were particularly vulnerable during higher altitude exposure, which was consistent with our present findings at MA. All these studies supported the viewpoint of Wilson and colleagues (2009), who concluded that altitude exposure mainly impaired short-term memory, with long-term memory preserved.
The effect of exposure to MA on cognitive function was mostly based on laboratory acute observation and was a controversial issue. Denison and colleagues (1966) reported an initial transient increase in reaction time on a spatial transformation task in exercising subjects at a simulated altitude of 2438 m. Similarly, Kelman and Crow (1969) found impairment of vigilance performance at a simulated altitude of 2438 m. McCarthy and colleagues (1995) found that significantly slower response times were obtained even at a lower altitude of 2134 m when subjects were required to make speeded judgments concerning the orientation of visual stimuli in a hypobaric chamber. Our present study demonstrated that long-term MA residents had performances similar to those of acute hypoxia-exposed subjects. Our study further confirmed the summary by McFarland (1938) that at altitude lower than 2500 m visual sensitivity and short-term memory had noticeable deficits (Maa, 2010). However, in contrast to the preceding observations, a number of acute studies found that cognitive functions were not impaired by MA up to 3658 m. Figarola and Billings (1966) observed no impairment on practiced tracking and vigilance tasks at 2438 m. Although the study of Fowler and colleagues (1987) showed impairments in visual and auditory reaction-time tasks or in perceptual motor performance occurring after acute exposures to simulated altitude at 3048 m, most studies have failed to test cognitive function changes around this altitude level. For example, Harrison and colleagues (2008) found that two-12 h sessions of exposure to an altitude of 3048 m in a simulated night-operational aircraft environment had no significant negative effects on grammatical reasoning, mathematical processing, simple reaction time, spatial processing ability, and short-term memory. Tripathi and colleagues (2005) did not find working memory and vigilance tasks changed after 36 h or 6 days of exposure to an altitude of 3200 m. Paul and Fraser (1994) found that the ability to perform spatial orientation, serial choice reaction time, and logical reasoning tasks were not impaired at altitudes of 1524, 2438, and 3048 m, whereas the impaired effect only occurred at an altitude of 3658 m. However, early in 1971, Crow and Kelman (1971) did not find short-term memory deficit after subjects' acute exposure to a simulated altitude of 3658 m. Further, enhanced cognitive function was tested at altitude higher than 3658 m. For example, Pagani and colleagues (1998) observed enhanced spatial working memory during a 15-day mountain trek climbing to 5350 m. Harris and colleagues (2009) found mountaineers performed better in the digital span forwards test after an 18-day ascent from 400 to 5100 m.
Almost all MA subjects complained of poor sleep quality during the first few weeks of their arrival, which conforms with previous data that poor sleep quality usually appeared at altitudes over 2000 m and increased at higher elevations during the initial 2 to 3 weeks of acclimatization (Buguet, 2007). However, sleep disturbances occurred in only the first few weeks of the stay at MA and did not continue throughout the MA stay. The higher incidence of abdominal pain in menstruation in MA females (vs. SL controls) also occurred in only the first few weeks of the stay at MA. Weight loss in MA subjects may be owing to a higher metabolic rate and reduced food intake, and hypobaric hypoxia seems to play a major role (Lippl et al., 2010). Taking physiological data into consideration, we suggest that our MA subjects were tested at the stage of good MA acclimatization. We did not find that physiological data, sleep disturbance, mood problems, or gender contributed to poor cognitive performances.
The cognition changes in acclimatized-altitude subjects may be related to glucose metabolism and neurotransmitter synthesis alteration. The immediate recall of both the ROCF test and WMS visual reproduction subtests requires active reorganization of the memory for successful recall, thus reflecting the comprehensive function of the frontal and parietal lobes (Loring et al., 1988). Previously, in a PET study with an altitude exposure period comparable with the current study, Hochachka and colleagues (1999) demonstrated reduced metabolic rates in three frontal areas of marines after 63 days living at altitudes between 3181 and 6157 m. Finally, animal models demonstrated the turnover of several neurotransmitters, such as acetylcholine synthesis, was altered by moderate hypoxia (Gibson et al., 1981).
The limitations of our present study were that we lacked both a SL baseline assessment for all participants and an additional assessment after brief altitude exposure before acclimatization had occurred to determine whether these subjects had worse performance when they first moved to altitude. Finally, the MA group had somewhat larger SDs on the tests, showing significant group differences and suggesting only a subset of MA subjects demonstrated significant altitude effects.
Conclusions
In summary, our study reveals slight effects of prolonged living at MA on human cognitive performances. Because of weak statistical power, clinically meaningful differences between the MA and the SL groups cannot be concluded. The present study may be valuable for human activities at altitude. Future research is needed to clarify the brain mechanisms involved in the poor visual construction that showed among the MA subjects and whether this low ability would be enhanced after return to SL.
Footnotes
Acknowledgments
Funding was provided by the China Postdoctoral Science Foundation (Project No. 20060390129) and the National Science Foundation of China (Project No. 30670674).
Disclosures
The authors have no conflicts of interest or financial ties to disclose.