
Abstract
Abstract
MacCormick, Ian J.C., John Somner, Daniel S. Morris, Thomas J. MacGillivray, Rupert R.A. Bourne, Suber S. Huang, Alasdair MacCormick, Peter A. Aspinall, J. Kenneth Baillie, A.A. Roger Thompson, and Bal Dhillon. Retinal vessel tortuosity in response to hypobaric hypoxia. High Alt Med Biol 13:263–268, 2012.— Purpose: Retinal vascular tortuosity is associated with retinopathy of differing etiologies, including hypertension, diabetes, and hypoxia. However, detailed understanding of the underlying pathophysiology is lacking. The aim of this study was to map changes in tortuosity associated with hypoxia at high altitude, and to determine the influence of sildenafil and an antioxidant preparation on altitude-induced tortuosity.
Methods:
We measured the tortuosity of retinal vessels using a semi-automated method in 35 young, healthy subjects exposed to hypobaric hypoxia for 7 days at 5200 m, and compared the measurements to those from the same vessels at sea level. These subjects simultaneously took part in a randomized double-blind placebo-controlled trial of sildenafil and antioxidant. Comparison of tortuosity between these subgroups was performed.
Results:
High altitude was associated with the development of retinal tortuosity in individual vessels. A nonsignificant trend suggests this is limited by prophylaxis with sildenafil or antioxidant.
Conclusions:
Retinal vessel tortuosity increases rapidly at high altitude. We suggest that retinal vessel tortuosity at altitude may result from increased sheer stress causing elongation of vessel segments and that this might be limited by agents that act to preserve nitric oxide dependent vasodilation.
Introduction
While the observation of tortuosity often signals disease, precise measurement of these changes could be used to quantify the degree of stress imposed on the retinal microvasculature. In circulations outside of the eye, vessel elongation (leading to tortuosity) is known to be caused by high flow rates (Sho et al., 2004), and much work has been involved in defining the precise biophysical parameters needed to cause changes in vascular structure (Hademenos and Massoud, 1997). In contrast, although the etiology of retinal vascular tortuosity can be roughly divided into broad categories (Sutter and Helbig, 2003), our understanding about the interaction between hemodynamic forces and components of the vessel wall remains incomplete.
Retinal tortuosity in the context of hypoxia is particularly interesting because of the close analogy between retinal and cerebral circulations. Retina and brain have common embryological origins, and both have important anatomical and physiological similarities. For example, both retinal and brain neurovasculatures have blood–retina or blood–brain barriers, and both perfuse tissue with extremely high metabolic rates (Patton et al., 2005; Forrester et al., 2008).
Specific retinal changes are associated with neurovascular conditions such as lacunar stroke (Doubal et al., 2010) and cerebral malaria (Beare et al., 2006).
Increased retinal vascular tortuosity at altitude has been noted previously (Rennie and Morrissey, 1975; Morris et al., 2006). However, it has not been quantified using image analysis of paired vessel segments in serial photographs at altitude and sea level. We aimed to confirm and quantify, using robust and reproducible methodology, the important observation of retinal vessel tortuosity in healthy subjects at high altitude, and to investigate dynamics of retinal vascular changes over time at altitude. Study subjects were also part of a nested drug trial (Baillie et al., 2009; Bates et al., 2011) and this gave the additional opportunity to compare tortuosity between drug groups.
Materials and Methods
Setting and participants
The retinal photographs analyzed in this study were taken during the Apex 2 altitude research expedition in 2003. One hundred and four healthy subjects flew from Edinburgh, United Kingdom (UK) (sea level) to La Paz, Bolivia, where they acclimatized for four days at 3600 m before being driven to the Chacaltaya Laboratory (5200 m). Retinal photographs were taken on days one, three, and seven at 5200 m, and a final post-expedition photograph was taken at sea level one month after subjects returned to the UK. Peripheral oxygen saturation was measured with a pulse oximeter. Acute mountain sickness (AMS) was assessed using the Lake Louise questionnaire. Subjects did not descend below 2000 m while in La Paz, or below 5000 m while at Chacaltaya. Vigorous exercise was discouraged while at altitude, and subjects did not smoke or drink alcohol during the study. All subjects on the Apex 2 expedition took part in multiple experimental protocols, including a randomized double blind placebo-controlled study of sildenafil citrate (Bates et al., 2011, NCT00627965) and an antioxidant mixture (Baillie et al., 2009, NCT00664001). The trial adhered to the tenets of the Declaration of Helsinki and ethical approval was granted by the Lothian Research Ethics Committee. All subjects gave informed consent, and took the drug or placebo for the duration of the time at altitude and follow-up test session.
Retinal photography
Digital retinal photographs were taken in a darkened room using a Nidek NM100 digital camera. Pupils were not dilated and images had a resolution of 144 pixels per inch.
Initial image analysis
The Retinal Diseases Image Analysis Reading Center (REDIARC) of Case Western Reserve University performed initial analysis of the retinal images using a semi-automated method. Two readers, blinded to testing session and drug group, independently assessed the images for quality using the OCULAB Image System (Digital Healthcare, UK). Photo quality was rated good, fair, or poor, depending on visibility of vessel borders.
Subsequent image analysis
Analysis of vessel tortuosity was carried out at the Image Analysis Core laboratory of the Welcome Trust Clinical Research Facility, Western General Hospital, Edinburgh. This was done by one reader who was blind to drug group and testing session of each retinal image. For the purposes of this study only, photographs from the left eye were analyzed. Analysis involved the following steps: color–gray conversion; image enhancement using Contrast Limited Adaptative Histogram Equalization (CLAHE); image filtering; image segmentation with hysteresis thresholding; skeletonization; and measurement of tortuosity after placing boundaries at 1–3×the optic disc diameter (Patton et al., 2005; MacGillivray et al., 2007). A custom vessel-tracking program was used, and tortuosity was measured using an established method as the ratio of curved to direct path length on four vessel segments per photograph (Fig. 1; Heart et al., 1999). This method does not take account of changes in vessel width.

The stages involved in image analysis and measurement of vessel tortuosity.
The same vessel segments were used consistently for all photographs within subjects to give paired data. Wherever possible, the vessel segments included a primary arteriole, a primary venule, a secondary arteriole, and a secondary venule. However, this was not always possible, and measurements from arterioles and venules were analyzed together. Primary vessel segments were measured between the optic disc edge and first bifurcation, or up to three times the optic disc diameter from the disc edge if there was no bifurcation within that distance. Secondary vessel segments were measured between the bifurcation from the parent vessel and three times the optic disc diameter. Tortuosity measurements were repeated three times for each vessel segment and averaged. For each photograph, an average tortuosity value was calculated based on values from four vessel segments. This value averages measurements from both arterioles and venules.
Sample size and drug groups
Sixty-two subjects had photographs taken at all four testing sessions. Images were screened for quality before further analysis, and 48 of 62 subjects had left eye photographs rated good or fair quality at all four testing sessions. A further five subjects were excluded due to missing data, leaving 43 subjects eligible for inclusion.
Custom software was needed to analyze the relatively low-resolution photographs. Unfortunately, this became prematurely unavailable and data collection from the series was halted at this point. Therefore, from the above eligible group of 43, data were collected from the first consecutive 35 subjects (18 females, 17 males, aged 18–29 years, mean age 21).
Of this group of 35, 11 subjects (8 males, 3 females) received 150 mg sildenafil per day in three divided doses, starting five days before the first testing session for the duration of the expedition, and commencing again five days before the post-expedition testing session. Ten subjects (4 males, 6 females) received antioxidant (1 g Vitamin C, 400 IU Vitamin E, and 600 mg α-lipoic acid) per day in four divided doses for the same period, and 14 received only identical placebo tablets (5 males, 9 females). Sildenafil and matched placebo were provided by Pfizer Ltd., UK, antioxidant and matched placebo were provided by Cultech Ltd., UK. Subjects in the sildenafil group also received antioxidant placebo, and vice versa. Subjects were allocated randomly to drug groups, and neither investigators nor subjects knew who was in each drug group until after data analysis from the expedition was complete.
Statistics
Repeated-measures ANOVA was performed to determine if a significant difference in vessel tortuosity existed between altitude and sea level (i.e., hypoxia and normoxia), or between drug groups at any time point. The within subject factor was tortuosity, the between subject factor was drug group. Mauchly's test of sphericity was significant (Chi Square 17.1, df=5, p=0.004) and in order to compensate for this a multivariate output was used.
Tortuosity was compared between subjects with AMS (a score of 3 or more) and no AMS (<3) on days 1, 3, and 7 at 5200 m using a double-sided t-test assuming unequal variances. Systolic blood pressure was also compared between sea level and day 1, day 3, and day 7 at 5200 m using paired t-tests, correcting for multiple comparisons.
Results
Descriptive statistics of each group at sea level are shown in Table 1. One subject in the placebo group did not have these measurements. Oxygen saturation varied between sea level and altitude, but not with time at altitude (Table 2).
There were 11 subjects in the sildenafil group, 10 in the antioxidant group, and 14 in the placebo group. Data are mean and standard deviation.
There were 11 subjects in the sildenafil group, 10 in the antioxidant group, and 14 in the placebo group. Data are mean and standard deviation.
Tortuosity was significantly greater at altitude compared to sea level (Roys root=13.8, df=3, p=0.001). Although there is a trend to greater tortuosity in the placebo group, the difference in tortuosity between drug groups and placebo was not significant (F=0.64, df=2, p=0.53). Tortuosity initially increased during time at altitude before reaching a plateau, increasing between days 1 and 3 at 5200 m (p=0.001) but not between days 3 and 7 (p=0.07) (Fig. 2).
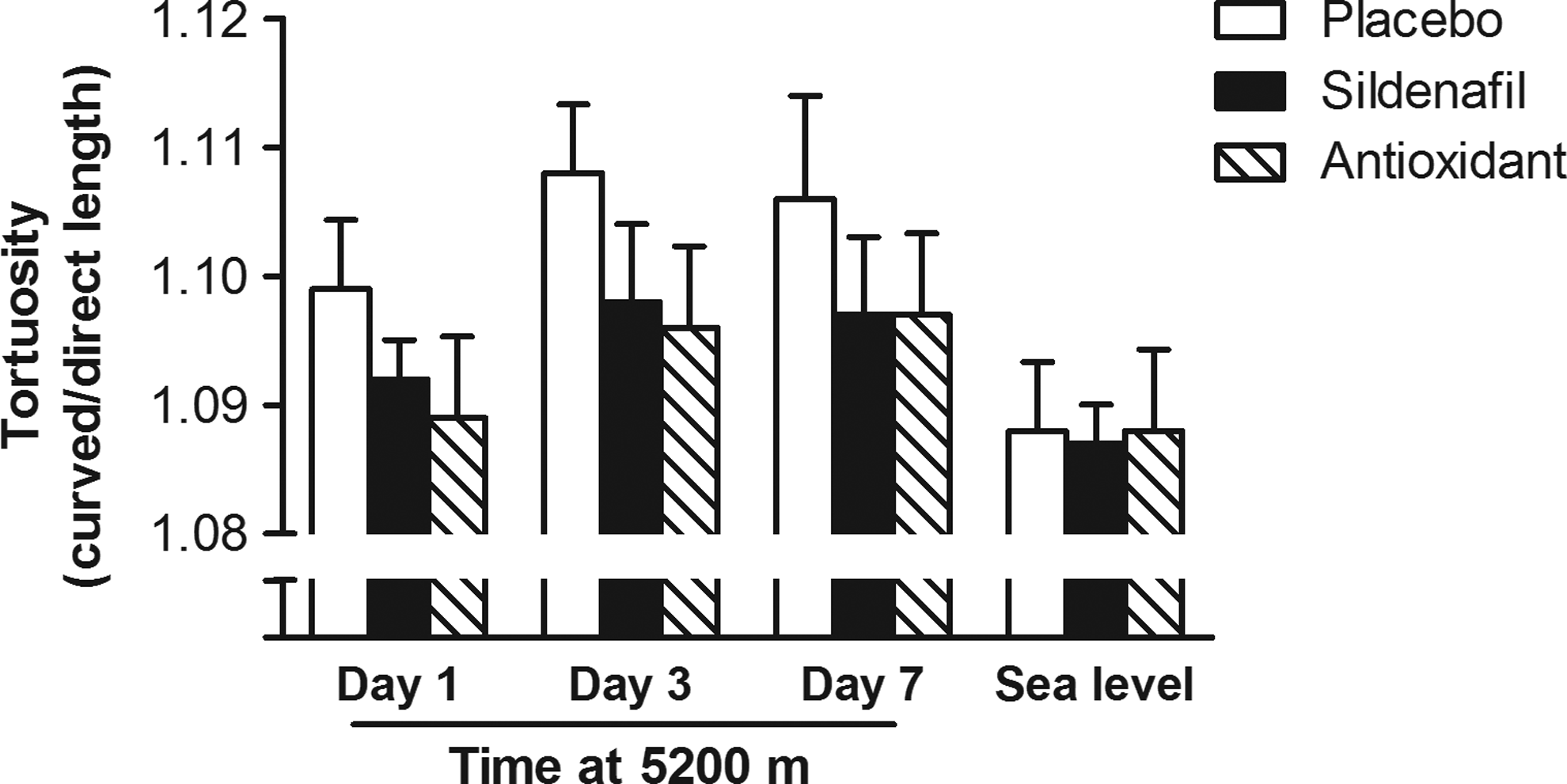
Retinal vessel tortuosity for each drug group (curved/direct length, mean and SEM) at each testing day at 5200 m and at sea level. In this study, tortuosity was an average of measurements from both arterioles and venules, and did not include measurements of vessel width. Tortuosity was significantly greater at altitude compared to sea level, but there was no significant difference between drug groups.
There was no significant difference between tortuosity in those with AMS compared to those with no AMS (day 1, p=0.2; day 3, p=0.2; day 7, p=0.7) (Fig. 3).
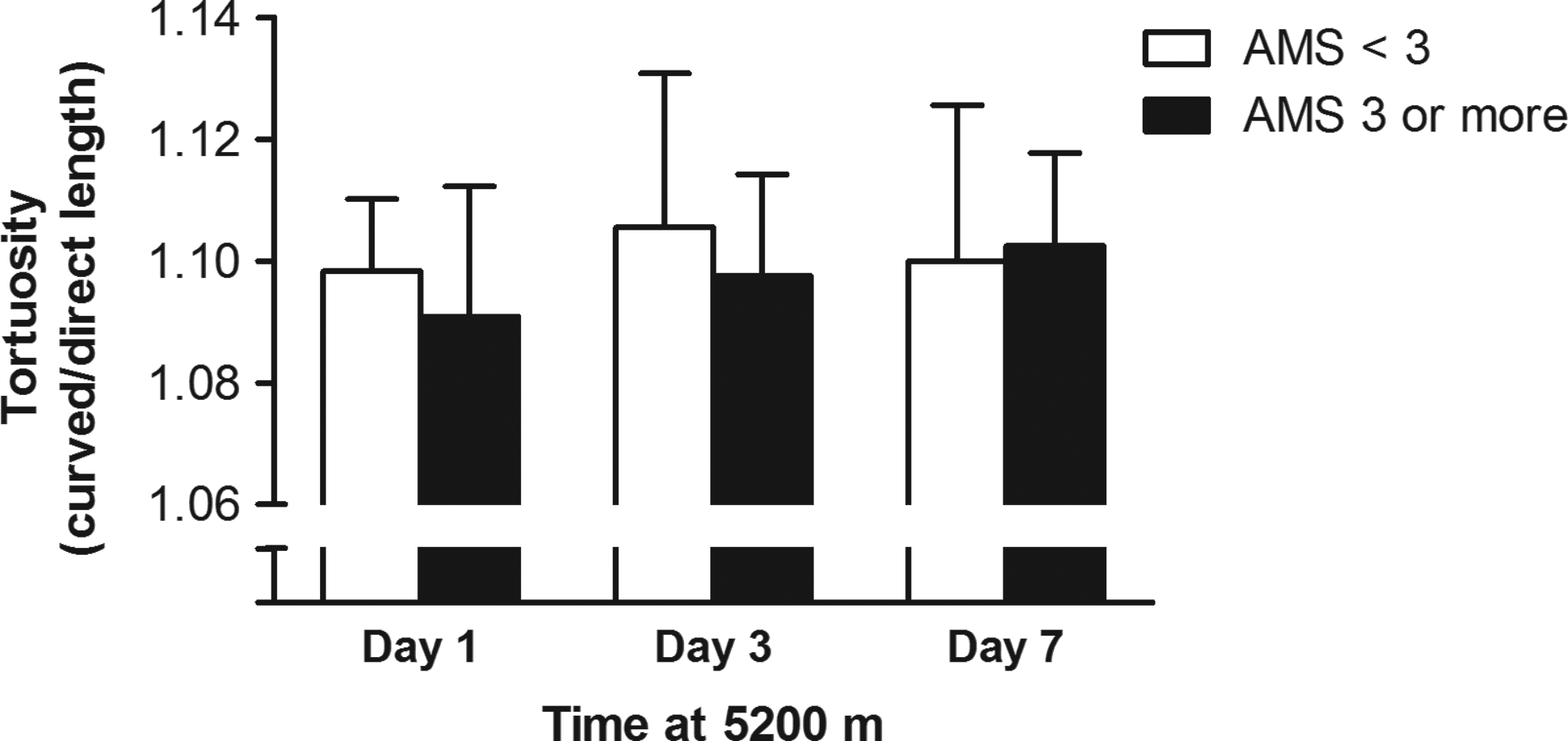
Mean and standard deviation tortuosity for the whole group (n=35), in those with AMS (≥3) or no AMS (<3) on days 1, 3, and 7 at 5200 m. The difference in tortuosity between the two groups was not significant on any day (p≥0.2).
Systemic blood pressure remained within normal values at all testing sessions. There was a difference of 14 mmHg in average systolic blood pressure between sea level and day 1 at 5200 m (p<0.001) (Fig. 4).
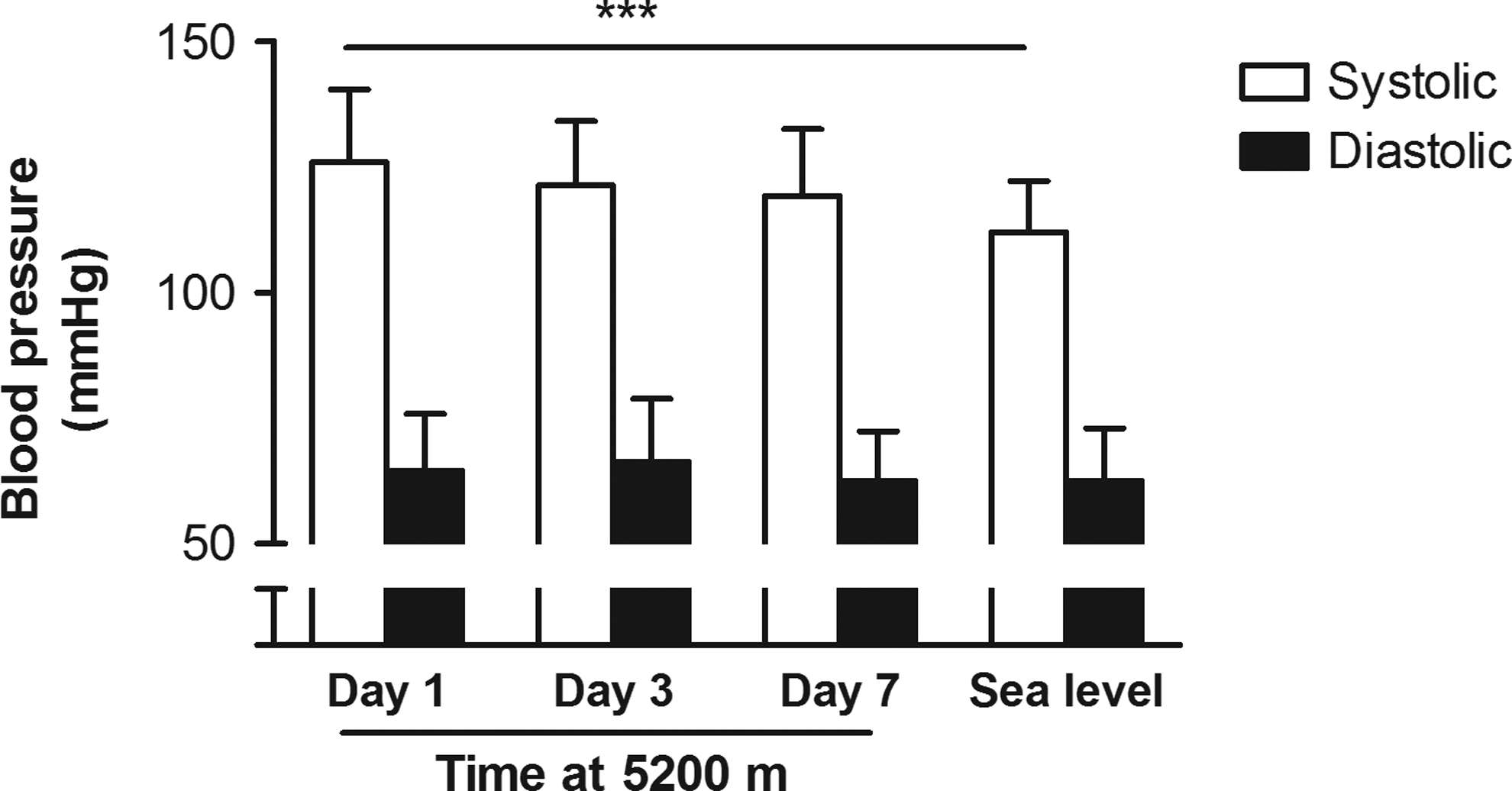
Mean and standard deviation systolic and diastolic blood pressure for the whole group (n=35) on days 1, 3, and 7 at 5200 m, and at sea level. Systolic blood pressure was significantly higher on day 1 at altitude compared to sea level (***p<0.001).
Discussion
This study found that retinal vessel tortuosity increases after acute exposure to hypobaric hypoxia in healthy subjects. There was no association between tortuosity and acute mountain sickness, and it is likely that the tortuosity observed in this particular study results from physiological adaptation to high altitude, rather than indicating retinal pathology.
Whilst retinal tortuosity is known to occur alongside retinal hemorrhages in conditions such as retinal vein occlusion, and retinal hemorrhage is a feature of high-altitude retinopathy (Morris et al., 2006), the microscopic changes we observed were much smaller than the clinically obvious vessel changes that occur in retinal vein occlusion. However, the type of methodology used here to quantify tortuosity will permit future investigation of the association between tortuosity and retinal hemorrhage at altitude.
Systolic blood pressure was greater at altitude than sea level. Ocular perfusion pressure is proportional to brachial artery pressure and intraocular pressure, and intraocular pressure increases slightly at altitude (Somner et al., 2007). The observed changes in systolic blood pressure could therefore influence development of tortuosity by increasing local flow velocity and shear stress through greater ocular perfusion pressure. However, although higher at altitude, systolic pressure remained within the normal range. Therefore, it is likely that local control of vessel tone and blood flow also have an important role in the development of increased tortuosity.
Basal arterial tone in vessels of both inner retina and choroid is regulated by continual release of nitric oxide (NO) from the endothelium (Dorner et al., 2003; reviewed by Schmetterer and Polak, 2001). NO is also produced in response to triggers such as sheer stress, and is necessary to increase retinal blood flow in response to hypoxia in animal models (Nagaoka et al., 2002). Shear stress exists because of friction between blood and the vessel wall, and is related to blood velocity, viscosity, and vessel width. If velocity and viscosity are constant, vessel dilation reduces shear stress. Similarly, if a vessel does not dilate, increasing velocity or viscosity causes greater shear stress (Popel and Johnson, 2005). NO mediates reactive vasodilation in response to shear stress, but hypoxia is associated with the generation of reactive oxygen species (ROS) (Magalhães et al., 2005; Kim et al., 2006), which inactivate NO through conversion to peroxynitrite. Besides this, hypoxia-induced hyperventilation at high altitude causes hypocapnia, which encourages arterial vasoconstriction. The combination of hypocapnic vasoconstriction and dysfunctional vasodilation due to ROS inactivation of NO could potentially lead to increases in retinal vessel sheer stress and subsequent tortuosity, since vessels that fail to dilate in the face of increasing flow velocity could stretch between fixed points. Indeed, this type of strain is known to cause elongation and widening of vessels outside the eye (Sho et al., 2004).
Sildenafil and antioxidants might then be expected to ameliorate vasoconstriction in retinal vessels at high altitude. This is because sildenafil selectively inhibits phosphodiesterase E5 (PDE5) and increases NO-dependent vasodilation by preventing endothelial cGMP breakdown in selective vascular beds, including the choroid (Lue, 2000; Schmetterer & Polak, 2001; Dorner et al., 2003; Bates et al., 2007). Since oxidative stress is known to inactivate NO, antioxidant supplementation could act to preserve NO bioavailability, allowing it to induce normal vasodilation in response to increases in blood velocity or viscosity. The effect of NO on retinal vessel architecture could be mediated directly, or through vascular steal directing a greater proportion of ocular blood flow to the choroid.
Unfortunately this study cannot answer questions about how altitude-associated tortuosity might be related to vessel widening, which is seen with tortuosity in some clinical conditions, such as venous congestion (Sutter and Helbig, 2003). Further research is needed to describe the interplay between neurovascular blood velocity and vessel geometry at altitude, and answer questions about when tortuosity is likely to be physiological and when it may represent a maladaptive response.
Having discussed the possible role of NO in tortuosity at altitude, it should be stressed that although a trend suggested that NO-conserving agents limited altitude-induced retinal tortuosity, this was not significant at our sample size. High altitude exposure, like other hypoxic states, is known to be associated with potentially protective endogenous antioxidant responses, including the release of urate from hypoxic tissue (Baillie et al., 2007), and supplementation with the antioxidant Vitamin E in the context of hypobaric hypoxia is known to reduce markers of oxidative damage (Simon-Schnass and Korniszewski, 1990; Magalhães et al., 2005). However, several trials have found antioxidant supplementation ineffective in reducing hypoxic pulmonary hypertension at altitude, another pathology involving NO-mediated vasodilation (Faoro et al., 2007; Basnyat et al., 2008; Baillie et al., 2009).
Limitations
Due to technical difficulties with image acquisition, a number of subjects were excluded from the study because of low image quality. In addition, data collection was halted prematurely. We have no reason to suspect that the technical factors that limited image quality would have any relationship to the presence or absence of retinal findings. Our cohort of 35 subjects on whom we completed image analysis is the largest such group to date.
Conclusions
This study found that retinal tortuosity increases rapidly following exposure to hypobaric hypoxia. It is possible that altitude-associated retinal tortuosity results from increased retinal blood flow velocity or viscosity under conditions that inhibit vasodilation at altitude. In this context, tortuosity could indicate longitudinal stretching of vessels that fail to dilate in the face of increasing shear stress.
The degree of change in tortuosity measured here is not detectable by standard clinical examination. This image analysis technique allows subtle changes in vessel shape to be quantified and potentially used as markers of vascular stress. Measurement of retinal vessel tortuosity is a cheap, noninvasive and responsive tool for assessing the microvascular impact of systemic hypoxia.
Footnotes
Acknowledgments
This research was supported by the registered Scottish charity, Apex (Altitude Physiology Expeditions SC030345; ![]() ). Pfizer UK supplied the sildenafil citrate and Cultech Ltd supplied the antioxidant cocktail. Image analysis software was developed with support from the Wellcome Trust Clinical Research Facility in Edinburgh. Sponsors had no role in study design; in collection, analysis, and interpretation of data; or in preparation of this manuscript.
). Pfizer UK supplied the sildenafil citrate and Cultech Ltd supplied the antioxidant cocktail. Image analysis software was developed with support from the Wellcome Trust Clinical Research Facility in Edinburgh. Sponsors had no role in study design; in collection, analysis, and interpretation of data; or in preparation of this manuscript.
We thank the Wellcome Trust Clinical Research Facility, Edinburgh; the Instituto de Investigaciones Fisicas, Universidad Mayor de San Andres; and the Instituto Boliviano de Biología de Altura, La Paz, Bolivia. Special thanks go to Kirsten Scott and Anna Thompson for helping with data collection on the Apex 2 expedition and to the team at REDIARC for the initial imaging analysis.
Disclosure Statement
No competing financial interests exist.