
Abstract
Abstract
Counter, S. Allen, Leo H. Buchanan, and Fernando Ortega. Brainstem auditory evoked responses in children living at high altitude in the Andes Mountains. High Alt Med Biol 14:155–161, 2013.—
Introduction
In field studies of children living under natural environmental hypoxic conditions at high altitudes in the Andes Mountains, we have recorded BAER as part of a clinical test battery (Counter et al., 1997; Counter, 2002). These studies have demonstrated the feasibility of recording BAER in the field in pediatric populations residing in high altitude areas. Altitude-induced alterations in latency and amplitude of the BAER of the children living at these elevations were not apparent, but in light of the findings of the previously referenced studies, should be systematically investigated. Further definition of the relationship between altitude and BAER would enhance our knowledge of the influence of elevated altitudes on neural synchrony and conduction in the central auditory brainstem pathways. The aim of the present study was to compare the latency and amplitude parameters of the BAER recorded in children living at high elevations with the BAER patterns obtained in a reference group of children of similar ages tested at sea level in a conventional neurophysiology clinic.
Materials and Methods
Participants and location
This study consisted of 34 children aged 5–15 years (mean age: 9.2 years; SD: 2.4) that comprised two groups, a high-altitude group and a low-altitude (sea-level) group. The high-altitude study group was comprised of 12 children, 6 females and 6 males aged 6.5–15 years (mean age: 10.2 years; SD: 2.9; median: 9) living at high elevations in Andean villages in Ecuador, South America, at altitudes of 2800 to 3000 meters. The children in the high-altitude group were examined at a makeshift testing site in the field, and selected for examination from several classrooms at local schools located in the study area. The sea-level reference group consisted of 22 children, 12 females and 10 males aged 5–12 years (mean age: 8.6 years; SD: 1.9; median: 9) from the U.S. who were tested in a conventional neurophysiology clinic on the same calibrated equipment used in the field. Biological calibrations were performed daily in the field on normal hearing staff to monitor the status of the equipment. Both the high-altitude group and the reference group were given otological examinations and audiologic tests prior to BAER testing. Children in both groups had a negative history of hearing loss or chronic middle ear pathology, and a negative family history of hearing loss. Any participant exhibiting a hearing loss was not included in the BAER testing. It should be noted that this study was conducted in the field in a nonstandard clinical situation, and was thus subject to the variables of field work. Unlike a controlled clinical environment, in field investigations, there are inherent uncontrollable variables and constraints, such as participants' availability, work and school schedules, time limitations, and in some instances, the inability to complete bilateral BAER measurements. For example, for one of the high-altitude participants in the present study, time constraints permitted measurement of the BAER in only one ear.
The children in the sea-level group received BAER testing as part of an audiological clinical test battery, and had no history of lead (Pb) exposure. The children in the high-altitude group reside in an area where Pb glazing of ceramics is a local cottage industry. However, each participant in the high-altitude group was found to have a blood Pb level less than 10 μg/dL (the traditional Pb-poison action line of the Centers for Disease Control and Prevention). Further, our previous BAER investigations in the same study area with children who exhibited plumbism revealed no significant association between Pb exposure and the clinical parameters of BAER (Counter et al., 1997; Counter, 2002). Informed consent was obtained from the parents or guardians of all children prior to testing. This study was approved by the Human Studies Committee (Comité de Bioética) of the Universidad San Francisco de Quito Medical School. The study was conducted under the auspices of the Universidad San Francisco de Quito Medical School in Quito, Ecuador.
Brainstem auditory evoked responses
BAER examinations were conducted using the Medlec/GSI-50 (Grason-Stadler, Inc., USA) eletrophysiologic system. The test environment was quiet and each BAER test was conducted using standard clinical protocols. The BAER procedure was used to examine the functional integrity of the 8th nerve, and ascending auditory brainstem tracts and nuclei, including summated response amplitude, absolute latency, and brainstem transmission times as indicated by interwave latencies. Each participant was tested in the supine position on a small bed or flat matted surface. The typical BAER test time was approximately 1½ hours. The participant's scalp, forehead, and ear lobes were thoroughly cleaned with swabs containing isopropanol, and gold-plated electrodes were placed at Cz (active), A1, A2 (reference), and Fpz (ground) according to the 10–20 International Electrode System. The electrode resistance was maintained at less than 5000 ohms. The electrodes (vertex positive) were connected to a physiologic pre-amplifier that amplified the ongoing electroencephalographic (EEG) activity by 105 times. A one-meter copper pipe that was placed deep (0.5 m) in the ground, and connected by wire to the electrophysiologic equipment, served as a grounding source. The BAER recordings were band-pass filtered between 100 and 3000 Hz, and the EEG activity was averaged at 1024 to 2048 sweeps per trial. Broadband monaural rarefaction click stimuli of 80 dB normal hearing level (nHL) and 100-μsec duration were delivered through TDH 39 earphones at rates of 10 and 50 pulses per second (pps) with contralateral masking. The 10 pps stimulus rate was used to conform with the standard BAER clinical protocol, while the higher stimulus rate of 50 pps, which more readily drives the auditory receptors and neurons to fatigue and adaptation (Don et al., 1977; Paludetti et al., 1983), was used to further test the physiological capacity of the auditory system in a high-altitude, low-oxygen environment. For the 12 participants in the high-altitude group, 23 ears were tested and analyzed, and for the 22 children in the sea-level reference group, 44 ears were tested and analyzed. The absolute latencies in milliseconds (ms) of wave peaks I, II, III, IV, and V were measured, and the interpeak latencies of I–III, III–V, and I–V were analyzed as indices of neural transmission (Counter, 2002). In addition, the amplitudes of waves I and V, and the V/I amplitude ratio were calculated. In the present study, the peak voltage rather than the shoulder of wave V was used consistently to specify the wave V latency for both the 10 pps and the 50 pps stimuli. Clinicians and researchers have used both the peak voltage and the shoulder voltage following the peak of wave V for specification of latency (Hall, 2007). The difference in calculating the wave V latency between the peak and shoulder would typically be a fraction of a millisecond. However, the important factor is that one or the other is used consistently, as in the current study. The amplitudes of waves I and V were measured from peak to succeeding trough.
Statistical analysis
Means, standard deviations, and percentiles were calculated for each BAER wave parameter in both the high-altitude group and the sea-level group. Because several of the BAER variables had skewed distributions, nonparametric statistical tests, as well as parametric tests, were used for data analysis. Both parametric and nonparametric analyses yielded comparable statistical results, but for clarity of data presentation, only the nonparametric statistical data are reported in the results. Differences between means were analyzed by the Mann-Whitney U test and the Wilcoxon Signed Rank tests. For the Mann-Whitney U and the Wilcoxon Signed Rank tests, the reported Z values and p values are tied values. An alpha level of ≤0.05 was accepted as an indication of statistical significance.
Results
For the sea-level altitude group at a click stimulus rate of 10 pps, replicated BAER Waves I, II, III, and V were identified in all ears (100% identification), and wave IV was not apparent in one ear of one participant (97.7% identification). At a click stimulus rate of 50 pps for the sea-level group, all waves were identified (100% identification). For the high-altitude group at a stimulus rate of 10 pps, BAER waves I, II, III, and V were identified in all ears (100% identification), and wave IV was identified in 91% of the ears. For the high-altitude group at a click stimulus rate of 50 pps, BAER waves I, III, and V were identified in all ears (100% identification), wave II in 86% of the ears, and wave IV in 94% of the ears. Means and percentiles of the BAER absolute latencies, using the conventional clinical stimulus rate of 10 pps for waves I, II, III, IV, and V for 23 ears of the 12 high-altitude participants and 44 ears of the 22 sea-level children are shown in Table 1, and illustrated in Figure 1A. Also shown in Figure 1A are sample BAER recordings for participants from the sea-level and high-altitude groups, illustrating the clarity and reliability of the replicated waveforms. Mean differences in the absolute latencies as presented in Table 1 are calculated by subtracting the mean absolute latencies of the sea-level group from those of the high altitude group. Mean differences in the absolute latencies between the high altitude and sea-level groups at a stimulus rate of 10 pps are shown in Table 1. For the 50 pps stimulus rate, mean differences between the high-altitude and sea-level groups were 0.052, 0.129, 0.104, 0.235, and 0.286 ms for BAER waves I, II, III, IV, and V, respectively. Data analysis using the Mann-Whitney U test revealed significantly longer BAER absolute wave latencies at a stimulus rate of 10 pps (Fig. 1A) for the high altitude group for waves I (Z=−3.17, p=0.001), II (Z=−2.36, p=0.018), IV (Z=−2.06, p=0.039), and V (Z=−2.71, p=0.006). The latency difference between the two groups for wave III was not statistically significant. Mean BAER absolute latencies at a stimulus rate of 50 pps for waves I, II, III, IV, and V for the high-altitude and sea-level groups are shown in Figure 1B. Sample 50 pps BAER recordings for individuals from the sea-level and high-altitude groups are shown in Figure 1B to illustrate the clarity and reliability of the replicated waveforms. Data analysis for the 50 pps stimulus rate using the Mann-Whitney U test revealed significantly longer BAER absolute wave latencies for the high altitude group for waves IV (Z=−2.34, p=0.019), and V (Z=−2.86, p=0.004). For BAER waves I, II, and III at the 50 pps stimulus rate, there were no significant differences in latencies between groups.
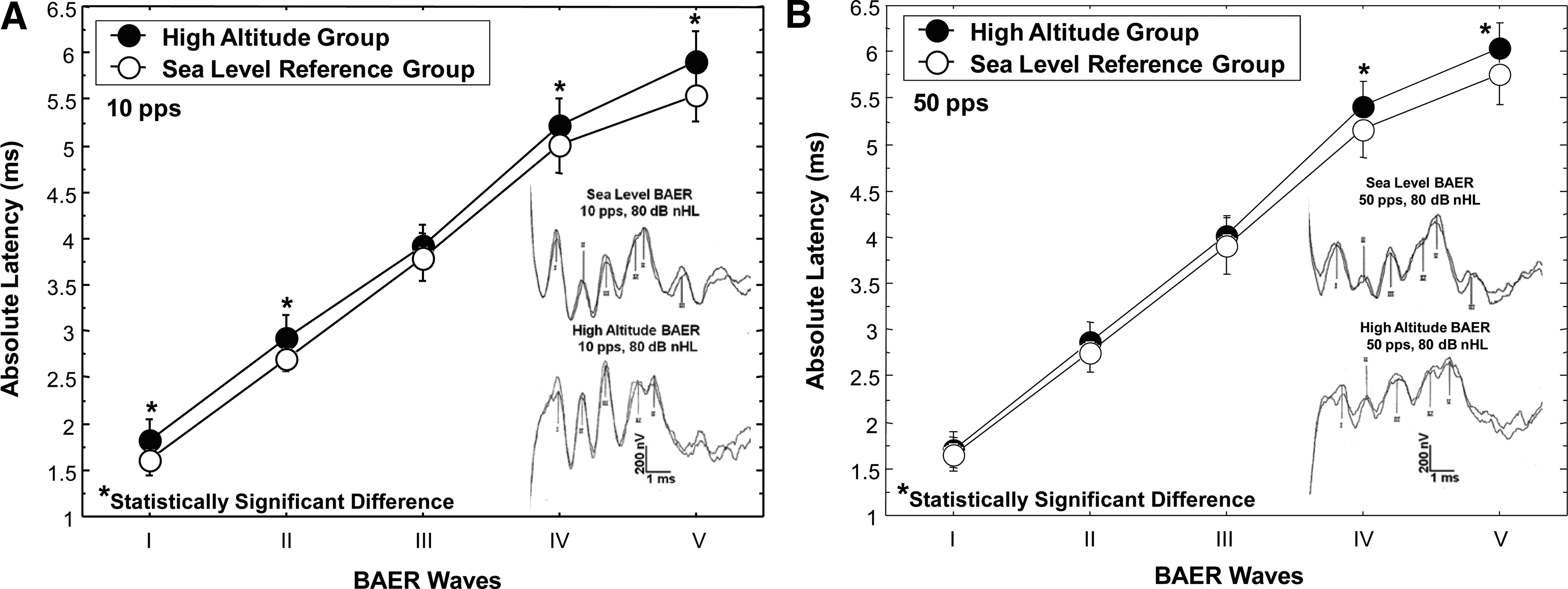
Mean absolute latency (ms) comparison of BAER waves I, II, III, IV, and V for the high-altitude group and a sea-level reference group.
Standard deviation values are shown in parentheses.
Figure 2A illustrates the mean BAER absolute latencies for waves I through V as a function of stimulus rate (10 and 50 pps) for the sea-level group. Figure 2B shows the mean BAER absolute latencies for waves I through V as a function of stimulus rate (10 and 50 pps) for the high-altitude group. As shown in Figure 2A, the sea-level group had significantly longer absolute latencies (Wilcoxon Signed Rank test) at the 50 pps stimulus rate compared to 10 pps for waves I (Z=−3.74, p=0.0002), II (Z=−2.49, p=0.012), III (Z=−4.72, p=<0.0001), IV (Z=−5.30, p=<0.0001), and V (Z=−5.36, p=<0.0001). Similarly, for the high-altitude group (Fig. 2B), BAER absolute latencies were significantly longer at the 50 pps stimulus rate than for 10 pps for waves II (Z=−2.94, p=0.003), III (Z=−3.08, p=0.002), IV (Z=−2.80, p=0.005), and V (Z=−3.30, p=0.001). There was no significant difference in latency between the 10 and 50 pps stimulus rates for wave I for the high-altitude group.

Mean absolute latency (ms) comparison of BAER waves I, II, III, IV, and V for click stimulus rates of 10 and 50 pulses per second (pps) for the sea-level reference group
The mean interpeak latencies and interpeak latency percentiles for waves I–III, III–V, and I–V at the conventional clinical stimulus rate of 10 pps are presented in Table 2. Mean differences in the interpeak latencies, as shown in Table 2, are calculated by subtracting the mean interpeak latencies of the sea-level group from those of the high-altitude group. For the 50 pps stimulus rate, mean differences in interpeak latencies between the high-altitude and the sea-level groups were 0.051, 0.195, and 0.233 ms for BAER interpeaks I–III, III—V, and I–V, respectively. The box plots of Figure 3 show the BAER medians and distributions for the interpeak latencies for 10 pps (Fig. 3A) and 50 pps (Fig. 3B) for the high-altitude and sea-level groups. Statistical analysis using the Mann-Whitney U test revealed significantly longer interpeak latencies at 10 pps (Fig. 3A) for the high-altitude group compared to the sea-level reference group for the interpeak wave latency of III–V (Z=−2.78, p=0.005), but not for the interpeak latencies of I–III and I–V. For the 50 pps stimulus rate (Fig. 3B), the high-altitude group had significantly longer interpeak latencies for interwaves III–V (Z=−3.24, p=0.001) and I–V (Z=−2.46, p=0.013), but not for the interpeak latency I–III.
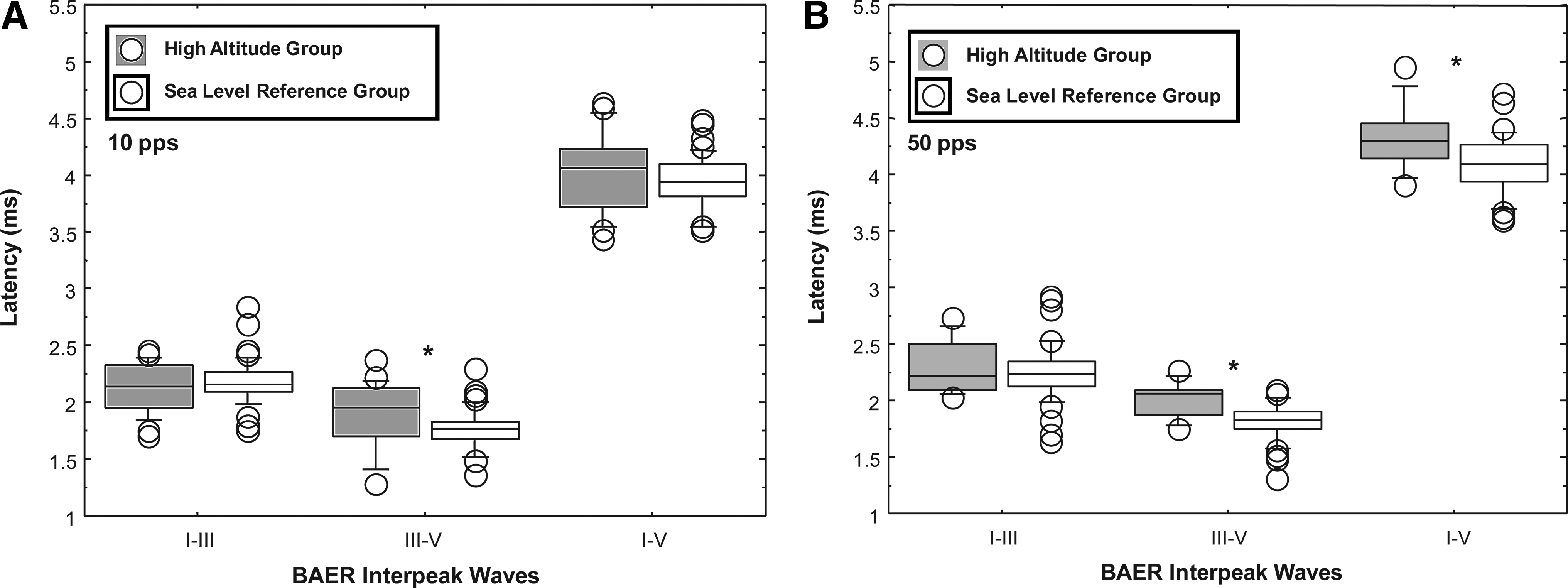
Box plots comparing the interpeak latencies (ms) of BAER waves I–III, III—V, and I–V for the high-altitude group (gray boxes) and a sea-level reference group (white boxes) at click stimulus rates of 10 pulses per second (pps)
Standard deviation values are shown in parentheses.
The box plots of Figure 4A and B illustrate the comparative peak-to-peak absolute amplitudes of waves I (Fig. 4A) and V (Fig. 4B) for the high-altitude group and the sea-level reference group at stimulus rates of 10 and 50 pps. As shown in Figure 4A, the peak-to-peak absolute amplitude for wave I tended to be lower for the high-altitude group for both the 10 pps and the 50 pps stimulus rates, but reached statistical significance only for the 10 pps stimulus (Z=−2.65, p=0.007). The peak-to-peak absolute amplitude for wave V (Fig. 4B) was significantly lower for the high-altitude group than for the reference group at both the 10 pps (Z=−2.71, p=0.006) and 50 pps (Z=−2.97, p=0.003) stimulus rates. The amplitude ratio of wave V to wave I (V/I ratio) was not significantly different for the two groups at the stimulus rates of 10 pps or 50 pps.
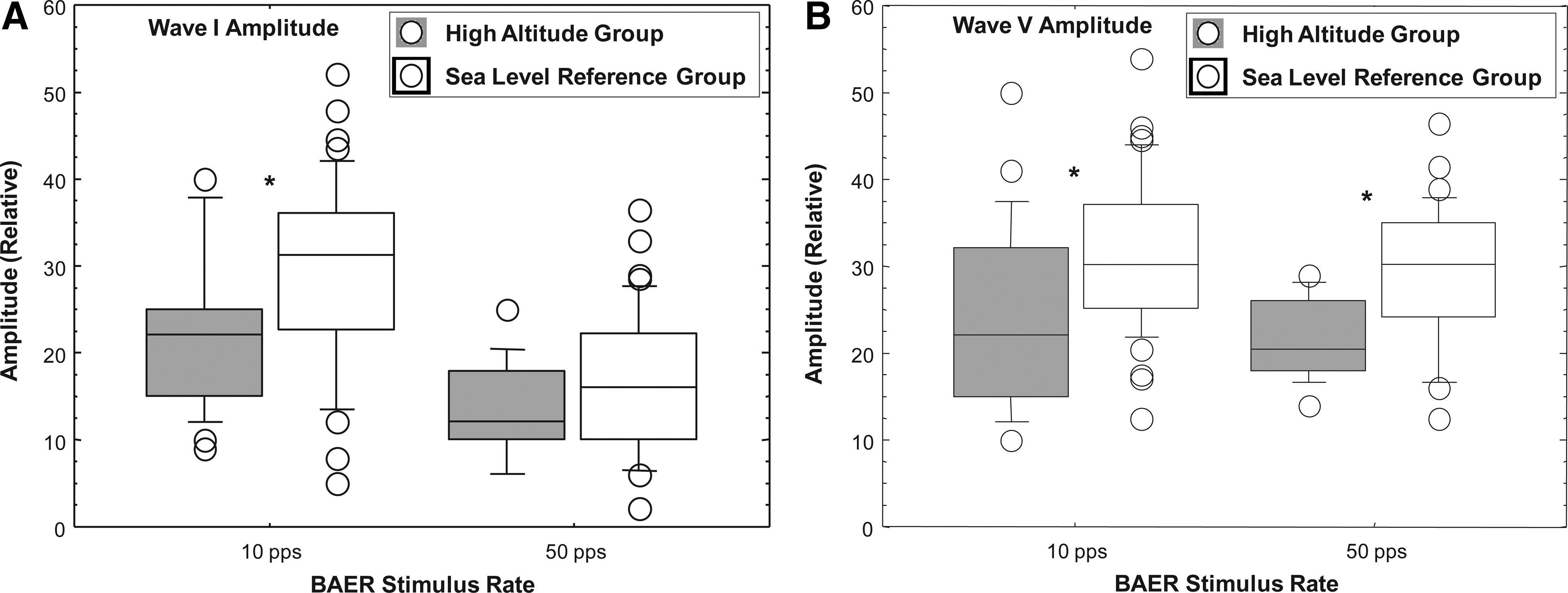
Box plots comparing the amplitude of BAER waves I
Discussion
A number of neurophysiological studies of acute natural hypoxia or simulated hypoxia have reported observable aberrations in human BAER related to high altitudes at around 3000 meters and above. These alterations in BAER amplitude and latency have been attributed to blood oxygen saturation, with hypoxia-induced BAER deficits being associated with blood oxygen saturation levels of between 75 to 85% (Carlile, 1992). Several neurophysiological mechanisms underlying hypoxia have been advanced, including changes in synaptic transmission in the neuronal network modulating the hypoxic respiratory response, and suppression of the membrane excitability of respiratory neurons through modulating ion channels (Hayashi and Fukuda, 2000). Further, some studies have suggested that the auditory brainstem is hypersensitive to small changes in blood oxygen saturation. However, the elevations relative to sea level necessary for adverse effects on the BAER have not been clearly elucidated. The present study investigated standard clinical parameters of the BAER recorded in children living in the Andes Mountains at an elevation close to 3000 meters, and compared the findings with BAER recordings from a typical pediatric reference group tested at sea level in a conventional neurophysiology clinic.
The results of the present study showed that several clinical parameters of the BAER in the high-altitude group (children living under natural environmental hypoxic conditions in the Andes Mountains) were significantly altered when compared with the BAER recorded from a sea-level group. For physiological comparison, the children in the high-altitude group and the sea-level reference group were tested at the standard BAER clinical stimulus rate of 10 pps, and at a higher stimulus rate of 50 pps. It has been shown that increased BAER latencies and reduced amplitudes occur with increasing stimulus repetition rates from 10 to 50 pps in normal subjects at sea level (Don et al., 1977; Paludetti et al., 1983). The higher stimulus rate of 50 pps may be expected to drive the neurons of the 8th nerve and central auditory neural pathways at a more physiologically taxing rate, thereby inducing greater neural desynchronization, fatigue and adaptation than the 10 pps rate, and thus may serve as a more sensitive indicant of the effects of low O2 saturation levels at high altitudes. Not unexpectedly, both the sea-level group and the high-altitude group showed increases in latency and reductions in amplitude of the BAER for the stimulus rate of 50 pps. One exception to this, however, was that the high-altitude group showed no significant difference in wave I latency for the two stimulus rate conditions. This was unexpected, and may be a chance finding that needs further investigation. It should be noted that the high-altitude group exhibited greater increases in BAER latencies and greater reduction in peak amplitudes for the 50 pps stimulus rate than the sea-level group. The finding of a comparatively greater reduction in the amplitudes of waves I and V in the high-altitude group for both 10 and 50 pps may suggest that at higher elevations, synaptic efficiency is reduced and fewer neurons are active in the summated BAER. Both the latency and amplitude results of this study at the 50 pps stimulus rate suggest that higher stimulus rates may provide greater information about the physiological effects of altitude on the auditory sensory-neural system.
The BAER absolute latencies of waves I, II, IV, and V, and the III–V interpeak latency were significantly prolonged in the high-altitude group at both stimulus rates. Since the I–V interval represents the long-range conduction from the 8th nerve to the area of the ipsilateral inferior colliculus, a number of neurobiological factors may affect the latency of neural transmission. However, in some neurologic and neurodevelopmental disorders, the neural transmission deficit may appear only in the I–III interval of the BAER (e.g., Maziade et al., 2000; Sostarich et al., 1993), which represents 8th nerve to superior olive transmission, or in the III–V interval as seen in the present study (e.g., Rosenhall et al., 2003; Selwa et al., 1993), which represents the area from the superior olivary complex via the lateral lemniscus to the inferior colliculus. As observed in some neurodegenerative diseases, it is quite possible to have segmental impairment along the tracts and nuclei conducting the auditory neural signal. Further, it has been shown that the firing rate of neurons that contribute to the BAER may differ in the lower brainstem area (ie, the cochlear nucleus and the superior olivary complex), which contribute to waves I through III, and the neurons of the more central nuclei, including the inferior colliculus, which is the putative generator of waves IV and V (Ballachanda et al., 1992). These differences in neuronal firing rates between the lower brainstem nuclei, and the higher brainstem nuclei may be a possible explanation for the differences observed in this study in the latencies of the later waves between the sea-level and high altitude groups. Another possible explanation for the differential effects of altitude on interwave intervals I–III and III–V that is worth noting has to do with maturation of the BAER, which matures from a caudal to rostral direction. Jiang et al. (2009) have shown differential maturational effects for I–III compared to III–V BAER interwave intervals in pre-term infants. Developmental aspects of the human BAER, however, are unlikely to explain the findings in the present study because, from previous studies (see Hall, 2007 for a review), the latency and amplitude of the human BAER, including the more central components as represented by the III–V response, is essentially mature by 18 month to 2 years of age (Hall, 2007). The children in the present study, who were 5–15 years of age, are well-above the maturational age for the BAER. The authors, however, are not aware of any studies that have investigated the developmental course of the BAER components as a function of altitude. This is probably worthy of study.
Tables 1 and 2 further illustrate numerically that the mean latency differences between the high-altitude and sea-level groups were within one standard deviation, and thus do not rise to the standard of clinical significance, but may have physiological import. The findings of this investigation suggest that the blood O2 saturation at an altitude of 2800 to 3000 meters alters subclinically auditory brainstem responses at the medulo-pontine/midbrain areas, as reflected in the observed alterations in the BAER amplitude and latency in the high-altitude group.
The observed effects on the BAER of children in the high-altitude group, although small, were unexpected, because the children in this group have lived since birth at high altitudes in the Andes Mountains, and are presumed to have adapted physiologically to the low oxygen and barometric conditions of their environment. It has been reported that persons living in high-altitude environments may possess adaptive phenotypes, including physiological traits such as high oxygen saturation of hemoglobin that naturally offset hypoxia (Scheinfeldt and Tishkoff, 2010). Other auditory physiological effects of high elevations have been reported, including increased intracochlear pressure and hemodynamic alterations (Cingi et al., 2010). Also, hydrostatic pressure matching between the middle ear cavity and the ambient pressure may be affected at high altitudes, resulting in decreased auditory sensitivity (Olzowy et al., 2008). These biophysical changes may influence several parameters of the BAER at high altitudes. However, there is no indication that these variables had any effect on BAER parameters in the present study. Nevertheless, further investigation of middle ear and cochlear functioning in populations living at high altitudes in the Andes may provide greater understanding of altitude effects on the auditory sensory-neural system.
In summary, the altitude-related subtle irregularities observed in this field study in the BAER recordings from children living at high elevations may have implications for auditory brainstem clinical assessment. Thus, small physiological effects of altitude on BAER, especially at elevations near 3000 meters and higher, may need to be taken into consideration when examining auditory brainstem integrity. Further study involving a larger number of children at different elevations is prudent before firm conclusions can be drawn regarding the extent of altitude effects on BAER in high altitude inhabitants.
Footnotes
Acknowledgments
The authors thank the administration of Universidad San Francisco de Quito, Fundacíon Capacitar of Ecuador, and the Ecuadorian health authorities for continued support of this project. We are grateful to the Harvard Biological Laboratories, the Harvard David Rockefeller Center for Latin American Studies, the Shriver Center/University of Massachusetts Medical School, the Harvard University Health Services and Dr. Jeremy Bloxham of Harvard University for support. We thank Anthony B. Jacobs for excellent technical assistance.
Disclosure Statement
The authors declare no financial conflict of interest.