Albert, Tyler J., and Erik R. Swenson. Peripheral chemoreceptor responsiveness and hypoxic pulmonary vasoconstriction in humans. High Alt Med Biol. 15:15—20, 2014.—Objective: Studies in animals have shown that interruption of carotid body afferent hypoxic signaling or efferent CNS activity to the lung enhances hypoxic pulmonary vasoconstriction (HPV). Whether a similar influence of the CNS on HPV strength is present in humans has never been studied, owing to the invasive nature of physical neural ablation or nonspecific systemic effects of pharmacological blockade of putative neural pathways. In order to demonstrate a peripheral chemoreceptor-mediated modulation of HPV in man, we hypothesized that individuals with high hypoxic ventilatory responsiveness, indicative of strong peripheral hypoxic chemosensitivity, should have less HPV in response to inspired hypoxia. Methods: In 15 healthy men and women, we measured the normobaric poikilocapnic hypoxic ventilatory response (HVR; L min−1 % SPo2−1) during 15 min of hypoxia (FIo2=0.12). On the following day, we then measured pulmonary artery systolic pressure (PASP) using echosonography while subjects randomly breathed 0.21, 0.18, 0.15, and 0.12 FIo2, each for periods of 15 min. We chose this strategy to obtain an equivalent stimulus for HPV in all subjects, using SPo2 as a surrogate for alveolar Po2. HPV was assessed as PASP at a common interpolated arterial oxygen saturation (SPo2) of 85%. Results: We recorded a sufficient six-fold range of HVR (0.05–0.30, mean 0.13 L min−1 % SPo2−1) similar to previously published data on normobaric, poikilocapnic HVR. HPV at SPo2 of 85% was 28.5 mmHg (range 21.7–41.3). There was a significant inverse relationship between poikilocapnic HVR and HPV (p=0.006, R2=0.38). Discussion: Previous studies of individuals with susceptibility to high altitude pulmonary edema (HAPE) have suggested that both low HVR and high HPV are important risk factors. We show that these two responses are inversely correlated and conclude that a greater magnitude of peripheral chemoreceptor response to hypoxia limits hypoxic pulmonary vasoconstriction in healthy subjects.
Introduction
Hypoxic pulmonary vasoconstriction (HPV) is a vasomotor response to alveolar hypoxia, mediating ventilation/perfusion (VA/Q) matching and optimizing arterial oxygenation (Pao2) by reducing blood flow to low VA/Q and shunt regions. It is intrinsic to pulmonary vascular smooth muscle, but importantly modulated by extrinsic factors including the vascular endothelium, erythrocytes, acid-base status, and neuro-humoral mediation (Swenson, 2013; Sylvester et al., 2012). Initially observed as pulmonary vasoconstriction with asphyxia, a critical landmark study demonstrated it was alveolar hypoxia and hypercapnia that elicited vasoconstriction (von Euler and Liljestrand, 1946). They proposed that HPV, and in a similar fashion hypercapnic pulmonary vasoconstriction, limits perfusion to lung regions with reduced or absent ventilation, occurring focally with conditions such as pneumonia, atelectasis, or edema. Interestingly, this was a concept presaged by JS Haldane a quarter century earlier (Haldane, 1922). A subsequent study showed that alveolar hypoxia in humans also induces pulmonary vasoconstriction (Motley et al., 1947). While HPV is an important mechanism of regional VA/Q matching in addition to concurrent CO2-mediated effects on vascular and airway tone and lung tissue compliance (Emery et al., 2013; Swenson et al., 1988), it may cause detrimental pulmonary hypertension when alveolar hypoxia is global, such as with hypoventilation, diffuse lung disease, or at high altitude.
The hypoxic ventilatory response (HVR), arising from stimulation of the peripheral chemoreceptors by low arterial Po2, increases ventilation to improve arterial oxygenation. While the critical link of carotid and aortic body chemoreceptors with HVR was first noted in the 1930s (Daly and Ungar, 1966; James and Daly Mde, 1969), it was not until the mid-century that an apparent input of the peripheral chemoreceptors to control of the pulmonary circulation was noted (Daly and Daly, 1959). They demonstrated a reduction in normoxic pulmonary artery pressure and vascular resistance when the isolated carotid bodies in dogs were perfused with hypoxic or hypoxic/hypercapnic blood. Subsequent studies have shown that denervation of the carotid bodies, resulting in loss of afferent input from the peripheral chemoreceptors and cerebral sensing of arterial Po2, enhances HPV (Levitzky et al., 1977; Naeije et al., 1989; Wilson and Levitzky, 1989). By contrast, when peripheral chemoreceptor activity is stimulated in animals with intact lung innervation, HPV is diminished (Chapleau et al., 1988; Fitzgerald et al., 1992; Levitzky, 1979). Because vagotomy blunts HPV (Daly and Daly, 1959; Naeije et al., 1989; Wilson and Levitzky, 1989), the efferent limb of peripheral chemoreceptor signaling is conveyed via innervation of the lung.
Although a link between the peripheral chemoreceptors and HPV appears well established in mammals, it has never been studied in humans, in whom such perturbations as carotid body denervation, isolated carotid body perfusion, lung denervation, or unapproved selective carotid body stimulants (such as almitrine) cannot be performed. Indirectly supportive of peripheral chemoreceptor modulation of HPV in humans is the finding that individuals with low HVR (and thus diminished peripheral chemoreceptor sensitivity) have exaggerated HPV and are at greatest risk for high altitude pulmonary edema (HAPE) (Bartsch et al., 2005). Thus, in an attempt to demonstrate a peripheral chemoreceptor-mediated modulation of HPV in humans, we hypothesized that HVR would be inversely correlated with HPV, such that individuals with high HVR (a response largely determined by peripheral chemosensitivity) would have less HPV.
Methods
Subjects
Nineteen healthy, normal weight subjects (15 male, 4 female; age range 26–37 years, mean 29.4) volunteered to take part in this study approved by the institutional review board (IRB) and ethics committee of the Veterans Affairs Puget Sound Medical Center, Seattle, Washington. All subjects provided informed written consent after receiving verbal and written instructions outlining the experimental procedures. Participants were not taking any medications, all were nonsmokers, all had refrained from alcohol or caffeine for at least 12 hours, and none had any history of cardiovascular or respiratory disease. Additionally, no subject had been above an altitude of 2000 m within the past 6 months. Each volunteer was familiarized with the laboratory facilities, especially with breathing through a mouthpiece while wearing a nose clip, and all subjects were required to make a preliminary visit to the laboratory for screening of a measurable tricuspid regurgitation (TR) by transthoracic echosonography (TTE).
Experimental protocol
Measurements were conducted over the course of 2 days at an elevation of 100 m and a barometric pressure of 760±5 mmHg. On the first day we measured normobaric, poikilocapnic HVR during 15 min of breathing 12% oxygen (FIo2=0.12). After undergoing a background medical history and physical examination, including blood pressure monitoring and baseline electrocardiography per IRB safety requirements, subjects were seated comfortably in an upright position and breathed through a mouthpiece connected to a Servo-I ventilator (Maquet, Inc) set in the low resistance continuous positive airway pressure (CPAP) mode without added positive airway pressure. In this fashion, ventilation was continuously measured. After a control period of 15 min breathing ambient air, the fraction of inspired oxygen was lowered from 0.21 to 0.12 with the appropriate addition of 100% nitrogen (N2) to the inspired gas mixture. Over the next 15 min, monitored parameters included SPo2, respiratory rate (RR), tidal volume (Vt), expired ventilation (VE), heart rate (HR), partial pressure of end-tidal CO2 (Petco2), arterial blood pressure by pressure cuff, inspired O2 concentrations (on a breath-by-breath basis using CO2 and O2 sensors), and continuous electrocardiogram. As this was a poikilocapnic protocol, Petco2 varied accordingly to the strength of each subject's hypoxic ventilatory drive. Hypoxic ventilation was averaged over the last 5 min of breathing 12% oxygen. HVR was calculated as change in ventilation from normoxia to hypoxia divided by the change in SPo2 (Δ ventilation/Δ SPo2) and given as L min−1 % SPo2−1.
On the second day, we assessed HPV by using TTE to calculate PASP while the subjects randomly breathed 0.21, 0.18, 0.15, and 0.12 FIO2, each for periods of 15 min while lying in a partial left lateral position. Again, monitored parameters included SPo2, RR, Vt, VE, HR, Petco2, arterial blood pressure by pressure cuff, inspired O2, and continuous electrocardiogram. Echocardiographic measurements were performed with an Acuson Sequoia C-512 cardiac ultrasound machine (Siemens, Inc) with a 4V1c two-dimensional transducer (2–4 MHz). PASP was measured by a blinded (to FIo2) echosonographer, and Doppler echocardiography was used to visualize a detectable regurgitant jet through the tricuspid valve during systole. Bernoulli's equation (ΔP=4V2) was used to calculate the pressure gradient across the valve, to which 5 mmHg was added as an estimate of right atrial pressure to yield PASP since all subjects displayed collapsibility of the inferior vena cava on inspiration.
Data and statistical analysis
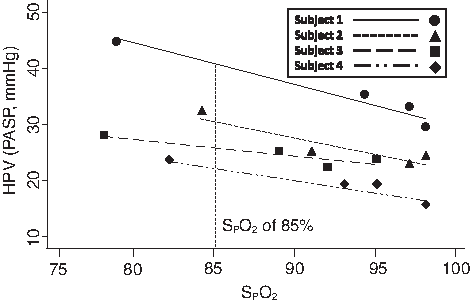
PASP was recorded at each FIo2 to construct a dose response of PASP versus SPo2, so that by interpolation (except in the case of subject # 10, in whom the value was extrapolated) PASP at SPo2 of 85% could be obtained for each subject, thus providing a common equal stimulus for HPV using SPo2 as a surrogate for PAo2 (Fig. 1). Although PAo2 was not measured, the range of small Petco2 changes with hypoxia (1–5 mmHg) resulting from the slight rise in ventilation in each individual would result in less than 1% deviation of Spo2 versus Po2 relationship caused by Bohr effect shifts in the hemoglobin–oxygen dissociation curve (Swenson and Hughes, 1993). Then, to eliminate any counteracting vasodilating effect of resultant hypocapnia, we recalculated that subject's effective isocapnic PASP. To do so, we estimated the separate contribution of hypocapnia to lower PASP in each subject by extrapolating from published data on this effect in humans (Balanos et al., 2003). This was calculated as 0.7 mmHg PASP×Δetco2 from each individual's PASP at 85% SPo2 and was added to their measured PASP. Both actual poikilocapnic HVR and calculated isocapnic PASP were then correlated with HVR. Statistical analysis was conducted with Stata (version 9.2). Linear regression was used to evaluate the relationship between HVR and HPV. A p value of<0.05 was used to determine statistical significance.
HPV assessed as PASP at a common interpolated SPo2 of 85% in four subjects.
Results
Four recruited participants (all male) did not have measurable TR and were excluded from the study. The other 15 subjects all completed the study protocol. Baseline results for these subjects included the following: SPo2 95–98%, mean 97%; Petco2 37–42, mean 39 mmHg; PASP 14–30, mean 23 mmHg. We recorded a broad range of HVR (0.05–0.3, mean 0.134 L min−1 % SPo2−1) equivalent to previously reported data (Steinback and Poulin, 2007). Measured Petco2 decreased with increasing HVR (0–5.0, mean 2.44 mmHg, p<0.035). As mentioned previously, HPV was measured as PASP at a common SPo2 of 85% (21.7–41.3, mean 28.5 mmHg). We found a significant inverse relationship between poikilocapnic HVR and HPV (p=0.006, R2=0.38) as seen in Figure 2. To correct for possible effects of CO2 on HPV, we calculated an isocapnic HPV assuming PASP decreased 0.7 mmHg/mmHg fall in Petco2 (Balanos et al., 2003). This correction factor agrees well with the slope of all of our PASP versus Petco2 data at SPo2=85% (-0.644 mmHg/mmHg Petco2). The calculated isocapnic HPV also showed a significant inverse relationship to the HVR (p=0.028, R2=0.28).
Poikilocapnic HVR is inversely correlated with HPV.
Discussion
We found a significant relationship between poikilocapnic HVR and HPV in healthy human subjects. The broad range of HVRs and changes in end tidal CO2 we recorded match previously published normobaric, poikilocapnic results (Steinback and Poulin, 2007). Peripheral chemoreceptor hypoxic sensitivity, as assessed indirectly by the poikilocapnic HVR, showed a significant inverse correlation with both poikilocapnic and effective isocapnic HPVs. These findings link the strength of the two most important acute responses to hypoxia and high altitude.
Our study is the first to explore directly whether a peripheral chemoreceptor-mediated modulation of HPV exists in humans, as has been clearly established in other mammals. In the only human study relevant to this issue and using a common Spo2 rather than a fixed FIo2 to measure PASP in hypoxia, Nespoulet et al. (2012) found that a lower mean HVR in subjects with acute mountain sickness (AMS) was associated with a higher mean PASP with hypoxia, in contrast to a higher HVR and lower PASP in those without AMS. However, they did not report any individual data, thus it is not possible to determine whether an inverse correlation such as the one we have shown exists. In various animal models, complete interruption of afferent signaling from the carotid bodies by denervation (Levitzky, 1979; Naeije et al., 1989) or elimination of efferent transmission via vagal innervation of the lung (Chapleau et al., 1988; Daly and Daly, 1959; Wilson and Levitzky, 1989) results in greater HPV for the same alveolar Po2. Furthermore, enhancement of peripheral chemoreceptor afferent output by isolated perfusion of the carotid body with hypoxic-hypercapnic blood results in reduction of HPV (Chapleau et al., 1988). Given the inability to perform any of these perturbations in our subjects, we approached the issue indirectly by demonstrating that individuals with higher HVR have lower HPV, suggesting a peripheral chemoreceptor-mediated modulation of pulmonary vascular tone in hypoxia in humans.
The neurobiology of the peripheral chemoreceptor link to HPV is not well understood and in part reflects our limited understanding of the complex nature of lung innervation. The pulmonary vasculature is innervated from the large conduit arteries and veins down to 50 μm vessels in larger species such as humans and canines, but much less so in smaller species (Kummer, 2011). In normoxia, the autonomic nervous system appears to exert an overall vasodilatory influence on the pulmonary circulation, with beta-adrenergic vasodilating activity overriding alpha-1 adrenergic mediated vasoconstriction (Murray et al., 1986). Other opposing vasodilatory neurotransmitters such as vasoactive intestinal peptide may also be released with pulmonary neural efferent signaling (Iwabuchi et al., 1997), along with parasympathetic innervation that is NO dependent (McMahon et al., 1992). Hypoxia, sensed at the peripheral chemoreceptors which project afferents to the medullary cardiovascular and respiratory control areas in the brain stem, activates both parasympathetic and sympathetic outflow to the lung. Studies using atropine and propranolol suggest that vasodilating parasympathetic activity is more dominant than sympathetic activity in HPV inhibition (Levitzky, 1979; Marshall, 1994; Wilson and Levitzky, 1989). However, other data suggest a stronger sympathetic contribution (Brimioulle et al., 1997), and not all studies find evidence for neural modulation of HPV (Lejeune et al., 1989; Liu et al., 2007; Lodato et al., 1988). The reasons for these discrepancies are not clear and may depend on the animal species studied, anesthesia, and differences in baseline pulmonary vascular tone. Most studies that find no effect of the peripheral chemoreceptors on HPV have largely employed adrenergic and cholinergic receptor blocking drugs rather than neural pathway interruption, thus it is entirely possible that peripheral chemoreceptor-mediated modulation of HPV may additionally involve non-adrenergic and non-cholinergic neurotransmitter release via lung innervation.
Studies of individuals who are susceptible to HAPE have suggested both low HVR and high HPV are important contributing risk factors (Bartsch et al., 2005). Our study shows that these two processes in humans are, in fact, inversely correlated, suggesting that peripheral chemoreceptor activity has an influence on the pulmonary circulation in hypoxia, as already shown in animal studies and described above. In this regard, we previously demonstrated that peripheral chemoreceptor sensitivity also alters renal function and urine output with acute hypoxic exposure, and are positively correlated (Swenson et al., 1995). Thus, we can speculate that peripheral chemoreceptor monitoring of arterial oxygen at high altitude enhances acute hypoxia tolerance and lessens the risk of HAPE by increasing ventilation to improve alveolar and arterial oxygenation, moderating pulmonary vasoconstriction to limit excessive pulmonary hypertension, and promoting increased sodium and water excretion to prevent fluid and edema accumulation.
Limitations of the study
We studied young, healthy subjects, possibly limiting extension of our findings to older patients and those with cardiopulmonary disease. Although HVR testing is traditionally done with maintenance of isocapnia in order to study the effect of hypoxia alone on peripheral chemoreceptor activity and organ function, spontaneously breathing animals will always have a variable degree of hypocapnia as a consequence of their increased hypoxic ventilation. As such, we opted to measure poikilocapnic HVR as opposed to isocapnic HVR in order to study the influence of combined hypoxia and hypocapnia on HPV, as these are the in vivo conditions most relevant to pulmonary vascular resistance when subjects breathe inspired gas of lower oxygen content or partial pressure. This decision created an inherent limitation to our results, as hypocapnia has a direct pulmonary vasodilating effect (Balanos et al., 2003), in addition to altering peripheral chemoreceptor activity (Blain et al., 2010). Although the confounding effect was small, we attempted to correct for any hypocapnic depression of HPV by calculating an “effective” isocapnic HPV from data in awake humans (Balanos et al., 2003) and still found an equally strong correlation of HVR and HPV. While this method yields only a rough estimate and certainly has its own limitations, it is perhaps validated by our own data as the co-efficient for end-tidal CO2 in our adjusted model was −0.644, virtually matching the co-efficient of −0.7 that we calculated from Balanos et al. (2003). Dependent on the magnitude of HVR, hypoxia evokes hyperventilation in part by larger tidal volumes, which can cause reflex systemic vasodilation (Marshall, 1994). There is, however, no evidence that increased lung inflation and increased ventilation at rest has any pulmonary vasodilating influence (Croft et al., 2013) via lung afferent signaling (Cassidy et al., 1979). Mixed venous Po2 can inhibit HPV if sufficiently low (Pellett et al., 1997), however the range at which this becomes evident (15–20 mmHg) was unlikely to have occurred at rest in our subjects as arterial oxygen saturations never fell below 70% (Table 1). An additional limitation is that the accuracy of transthoracic echocardiography is less than perfect, and it does not measure pulmonary vascular resistance. However, under resting conditions in healthy humans, PASP changes with hypoxia reflect increased pulmonary vascular resistance with good fidelity (Allemann et al., 2000), as the changes in cardiac output and pulmonary capillary wedge pressure are quite small (Allemann et al., 2000; Reeves et al., 1987). Lastly, HVR and HPV were measured one day apart in each individual. Week to week variability in HVR has been studied in subjects over 4–7 weeks and found to have a coefficient of variation of 15%–25% ( Macnutt et al., 2011; Sahn et al., 1977; Zhang and Robbins, 2000). This is to be contrasted with the very small variability of approximately 3%–4% within a single day (Sahn et al., 1977). To our knowledge, there are no data on next day variability in HVR measurements, but any such variability would be much smaller than weekly variability and would have more likely acted to diminish the correlations rather than create or increase their magnitude.
Individual Subject Data (mean±SD)
FIO2 0.21
0.18
0.15
0.12
HR
RR
VE
EtCO2
SpO2
PASP
HR
RR
VE
EtCO2
SpO2
PASP
HR
RR
VE
EtCO2
SpO2
PASP
HR
RR
VE
EtCO2
SpO2
PASP
HVR
Subject
(per min)
(L/min)
(mmHg)
%
(mmHg)
(per min)
(L/min)
(mmHg)
(%)
(mmHg)
(per min)
(L/min)
(mmHg)
(%)
(mmHg)
(per min)
(L/min)
(mmHg)
(%)
(mmHg)
L/min/%SpO2
1
67 (3)
17 (4)
6.7 (0.7)
43 (1)
98 (1)
29.2
65 (3)
18 (3)
6.7 (0.6)
44 (1)
95 (3)
33.9
70 (4)
13 (5)
7.4 (1.3)
42 (2)
93 (3)
30.5
73 (5)
15 (4)
8 (1.7)
42 (1)
83 (3)
30.7
0.15
2
57 (6)
11 (2)
7.1 (1.1)
43 (1)
95 (2)
29.1
61 (5)
12 (3)
8.3 (2.3)
44 (4)
93 (2)
31.8
61 (1)
10 (2)
7 (0.9)
43 (1)
87 (5)
30.1
65 (3)
11 (2)
6.7 (0.9)
43 (2)
74 (3)
29.7
0.18
3
79 (5)
22 (3)
7 (0.2)
37 (1)
97 (3)
28.8
79 (2)
20 (5)
7.3 (0.9)
36 (2)
98 (1)
32.9
80 (9)
24 (4)
7.3 (0.7)
37 (1)
93 (2)
35
87 (7)
23 (5)
8 (0.2)
36 (0)
80 (2)
45.5
0.07
4
73 (2)
23 (5)
7.0 (1.3)
46 (2)
95 (1)
28.8
75 (5)
19 (5)
7.7 (1.4)
43 (6)
88 (1)
25.7
79 (2)
21 (3)
7.5 (0.6)
45 (1)
89 (3)
27.1
82 (5)
20 (6)
8.8 (1.6)
43 (2)
78 (1)
25.7
0.21
5
56 (7)
11 (2)
4.8 (0.5)
44 (2)
95 (1)
24.2
57 (4)
10 (2)
5.4 (0.3)
46 (2)
93 (1)
21.5
58 (3)
10 (2)
5.9 (0.9)
44 (1)
90 (3)
25.2
68 (8)
10 (1)
6.6 (1)
44 (2)
80 (3)
27.7
0.27
6
66 (1)
14 (4)
6.7 (1)
39 (1)
98 (0)
22.4
66 (2)
11 (2)
5.7 (0.6)
44 (1)
95 (2)
31.7
65 (1)
12 (1)
6.4 (0.4)
40 (2)
93 (3)
30.5
76 (3)
11 (1)
6.3 (0.6)
43 (1)
81 (3)
33.3
0.07
7
63 (5)
16 (2)
6.7 (1.1)
36 (1)
98 (1)
16.5
65 (5)
18 (2)
6.7 (0.7)
34 (2)
97 (1)
19.5
67 (3)
22 (4)
7.2 (0.6)
35 (1)
94 (1)
19.3
72 (2)
23 (4)
8 (0.6)
35 (2)
85 (3)
23
0.3
8
53 (4)
24 (2)
8.3 (1.5)
46 (3)
96 (1)
14
53 (6)
21 (4)
7.9 (0.9)
46 (4)
93 (1)
21.1
52 (3)
21 (2)
8.1 (0.9)
47 (1)
89 (1)
23.4
63 (6)
23 (4)
8.2 (0.7)
46 (2)
78 (2)
29.8
0.16
9
49 (6)
22 (8)
4.7 (0.8)
41 (4)
98 (0)
18
49 (3)
18 (1)
4.3 (0.6)
40 (3)
97 (1)
16.5
52 (4)
16 (4)
3.9 (0.4)
43 (1)
94 (3)
27.9
64 (4)
19 (5)
4.1 (0.4)
43 (2)
78 (2)
30.8
0.05
10
45 (5)
17 (4)
11.8 (3.9)
35 (4)
98 (0)
17.2
42 (3)
17 (2)
13.8 (1.8)
28 (2)
99 (0)
20.5
45 (2)
19 (5)
11.3 (2.5)
26 (1)
98 (1)
22.3
45 (3)
16 (2)
10.4 (1.4)
26 (1)
95 (2)
22.3
0.24
11
70 (2)
11 (3)
5 (0.4)
41 (2)
98 (0)
29.3
69 (3)
14 (4)
5.5 (1.2)
40 (1)
98 (1)
28.5
74 (6)
14 (4)
4.8 (0.5)
41 (1)
94 (2)
28
79 (4)
12 (3)
5.5 (0.2)
39 (1)
82 (1)
36.4
0.04
12
58 (4)
17 (2)
5.5 (0.5)
39 (1)
98 (1)
24.2
63 (1)
12 (3)
5.4 (0.5)
40 (1)
96 (3)
22.6
65 (8)
12 (2)
5.3 (0.7)
38 (2)
91 (2)
25.3
66 (4)
13 (2)
6.5 (.5)
35 (1)
85 (1)
32.1
0.07
13
73 (5)
16 (1)
7.9 (0.7)
39 (1)
97 (1)
15.6
72 (3)
18 (1)
7.6 (0.7)
39 (1)
97 (1)
20.9
77 (4)
15 (1)
8 (0.5)
38 (1)
94 (2)
19.1
80 (5)
17 (1)
8.5 (0.9)
37 (1)
87 (2)
22.1
0.18
14
58 (3)
13 (2)
7.1 (2.3)
49 (2)
96 (0)
29.9
60 (4)
12 (1)
7 (0.6)
46 (2)
93 (1)
28.2
61 (3)
16 (3)
6.5 (0.7)
44 (2)
90 (3)
28.7
62 (4)
13 (1)
7.9 (1.5)
44 (1)
80 (1)
34.8
0.07
15
68 (2)
12 (2)
7.4 (2.6)
39 (2)
98 (1)
25.6
71 (4)
10 (1)
6.7 (1.1)
41 (2)
95 (1)
19.4
73 (4)
11 (2)
6.1 (0.7)
42.5 (2)
88 (1)
23.8
80 (6)
9 (1)
6.7 (1.3)
44 (1)
78 (2)
32.8
0.26
The data represent the average of values taken in the last 5 min of each hypoxic exposure. Of note, HVR was calculated from data obtained on the preceding day when this test was performed (see Methods section).
In conclusion, our results show an inverse correlation between peripheral chemoreceptor-mediated ventilatory response to hypoxia and hypoxic pulmonary vasoconstriction in healthy subjects. While animal studies have shown a direct moderating influence of the peripheral chemoreceptors on HPV magnitude by complete ablation of peripheral chemoreceptor afferent signaling to the CNS or its efferent signaling to the lung, our data are the first in humans providing evidence of the same phenomenon. Greater elucidation of this peripheral chemoreceptor-lung vascular relationship may yield better understanding of pulmonary hypertension and new means of treatment.
Footnotes
Acknowledgments
The authors would like to thank Patricia Sills for her extraordinary teaching, skill, and patience in the acquisition of the echocardiographic estimates of PA pressure, and Whitney Harrington for her outstanding statistical prowess.
Author Disclosure Statement
No competing financial interests exist.
References
1.
AllemannY, SartoriC, LeporiM, et al. (2000). Echocardiographic and invasive measurements of pulmonary artery pressure correlate closely at high altitude. Am J Physiol, 279:H2013–2016.
2.
BalanosGM, TalbotNP, DorringtonKL, and RobbinsPA. (2003). Human pulmonary vascular response to 4 h of hypercapnia and hypocapnia measured using Doppler echocardiography. J Appl Physiol, 94:1543–1551.
3.
BartschP, MairbaurlH, MaggioriniM, and SwensonER. (2005). Physiological aspects of high-altitude pulmonary edema. J Appl Physiol, 98:1101–1110.
4.
BlainGM, SmithCA, HendersonKS, and DempseyJA. (2010). Peripheral chemoreceptors determine the respiratory sensitivity of central chemoreceptors to CO2. J Physiol, 588:2455–2471.
5.
BrimioulleS, VachieryJL, BrichantJF, DelcroixM, LejeuneP, and NaeijeR. (1997). Sympathetic modulation of hypoxic pulmonary vasoconstriction in intact dogs. Cardiovasc Res, 34:384–392.
6.
CassidySS, EschenbacherWL, and JohnsonRL, Jr. (1979). Reflex cardiovascular depression during unilateral lung hyperinflation in the dog. J Clin Invest, 64:620–626.
7.
ChapleauMW, WilsonLB, GregoryTJ, and LevitzkyMG. (1988). Chemoreceptor stimulation interferes with regional hypoxic pulmonary vasoconstriction. Respir Physiol, 71:185–200.
8.
CroftQ, FormentiF, TalbottN, LunnD, RobbinsP, and DorringtonK. (2013). Variations in alveolar partial pressure for carbon dioxide and oxygen have additive not synergistic effects on human pulmonary vasoconstriction. PLoS One, 8:e67886
9.
DalyID, and DalyMD. (1959). The effects of stimulation of the carotid body chemoreceptors on the pulmonary vascular bed in the dog: The 'vasosensory controlled perfused living animal' preparation. J Physiol, 148:201–219.
10.
DalyM, and UngarA. (1966). Comparison of the reflex responses elicited by stimulation of the separately perfused carotid and aortic body chemoreceptors in the dog. J Physiol, 182:379–403.
11.
EmeryMJ, EvelandR, MinJH, HildebrandtJ, SwensonER. (2013) Carbon dioxide relaxation of the rat lung parenchyma. Respir Physiol Neurobiol., 186:33–39.
12.
FitzgeraldRS, DehghaniGA, ShamJS, ShirahataM, and MitznerWA. (1992). Peripheral chemoreceptor modulation of the pulmonary vasculature in the cat. J Appl Physiol, 73:20–29.
13.
HaldaneJS. (1922). Respiration. Yale University Press, New Haven, CT.
14.
IwabuchiS, OnoS, TanitaT, KoikeK, and FujimuraS. (1997). Vasoactive intestinal peptide causes nitric oxide-dependent pulmonary vasodilation in isolated rat lung. Respiration, 64:54–58.
15.
JamesJE, and Daly MdeB. (1969). Cardiovascular responses in apnoeic asphyxia: Role of arterial chemoreceptors and the modification of their effects by a pulmonary vagal inflation reflex. J Physiol, 201:87–104.
16.
KummerW. (2011). Pulmonary vascular innervation and its role in responses to hypoxia: Size matters!. Proc Am Thorac Soc, 8:471–476.
17.
LejeuneP, VachieryJL, LeemanM, et al. (1989). Absence of parasympathetic control of pulmonary vascular pressure-flow plots in hyperoxic and hypoxic dogs. Respir Physiol, 78:123–133.
18.
LevitzkyMG. (1979). Chemoreceptor stimulation and hypoxic pulmonary vasoconstriction in conscious dogs. Respir Physiol, 37:151–160.
19.
LevitzkyMG, NewellJC, KrasneyJA, and DuttonRE. (1977). Chemoreceptor influence on pulmonary blood flow during unilateral hypoxia in dogs. Respir Physiol, 31:345–356.
20.
LiuC, SmithTG, BalanosGM, et al. (2007). Lack of involvement of the autonomic nervous system in early ventilatory and pulmonary vascular acclimatization to hypoxia in humans. J Physiol, 579:215–225.
21.
LodatoRF, MichaelJR, and MurrayPA. (1988). Absence of neural modulation of hypoxic pulmonary vasoconstriction in conscious dogs. J Appl Physiol, 65:1481–1487.
22.
MacnuttMJ, De SouzaMJ, TomczakSE, HomerJL, and SheelAW. (2011). Resting and exercise ventilatory chemosensitivity across the menstrual cycle. J Appl Physiol, 112:737–747.
23.
MarshallJM. (1994). Peripheral chemoreceptors and cardiovascular regulation. Physiol Rev, 74:543–594.
24.
McMahonTJ, HoodJS, and KadowitzPJ. (1992). Pulmonary vasodilator response to vagal stimulation is blocked by N omega-nitro-L-arginine methyl ester in the cat. Circ Res, 70:364–369.
25.
MotleyHL, CournandA, et al. (1947). The influence of short periods of induced acute anoxia upon pulmonary artery pressures in man. Am J Physiol, 150:315–320.
26.
MurrayPA, LodatoRF, and MichaelJR. (1986). Neural antagonists modulate pulmonary vascular pressure-flow plots in conscious dogs. J Appl Physiol, 60:1900–1907.
27.
NaeijeR, LejeuneP, LeemanM, MelotC, and ClossetJ. (1989). Pulmonary vascular responses to surgical chemodenervation and chemical sympathectomy in dogs. J Appl Physiol, 66:42–50.
28.
NespouletH, WuyamB, TamisierR, et al. (2012). Altitude illness is related to low hypoxic chemoresponse and low oxygenation during sleep. Eur Respir J, 40:673–680.
29.
PellettAA, CairoJM, and LevitzkyMG. (1997). Hypoxemia and hypoxic pulmonary vasoconstriction: Autonomic nervous system versus mixed venous PO2. Respir Physiol, 109:249–260.
30.
ReevesJT, GrovesBM, SuttonJR, et al. (1987). Operation Everest II: Preservation of cardiac function at extreme altitude. J Appl Physiol, 63:531–539.
31.
SahnSA, ZwillichCW, DickN, et al. (1977). Variability of ventilatory responses to hypoxia and hypercapnia. J Appl Physiol, 43:1019–1025.
32.
SteinbackCD, and PoulinMJ. (2007). Ventilatory responses to isocapnic and poikilocapnic hypoxia in humans. Respir Physiol Neurobiol, 155:104–113.
33.
SwensonE. (2013). Hypoxic pulmonary vasoconstriction. High Alt Med Biol, 14:101–110.
34.
SwensonER, DominoKB, and HlastalaMP. (1988). Physiological Effects of Oxygen and Carbon Dioxide. Marcel-Dekker Press, New York. pp. 511–547.
35.
SwensonER, DuncanTB, GoldbergSV, RamirezG, AhmadS, and SchoeneRB. (1995). Diuretic effect of acute hypoxia in humans: Relationship to hypoxic ventilatory responsiveness and renal hormones. J Appl Physiol, 78:377–383.
36.
SwensonER, and HughesJM. (1993). Effects of acute and chronic acetazolamide on resting ventilation and ventilatory responses in men. J Appl Physiol, 74:230–237.
von EulerU, and LiljestrandG. (1946). Observations on the pulmonary arterial blood pressure in the cat. Acta Physiol Scand, 12:301–320.
39.
WilsonLB, and LevitzkyMG. (1989). Chemoreflex blunting of hypoxic pulmonary vasoconstriction is vagally mediated. J Appl Physiol, 66:782–791.
40.
ZhangS, and RobbinsPA. (2000). Methodological and physiological variability within the ventilatory response to hypoxia in humans. J Appl Physiol, 88:1924–1932.