
Abstract
Abstract
Xiangyang, Li, Xuejun Wang, Yongping Li, Junbo Zhu, Xiaodong Su, Xingchen Yao, Xueru Fan, and Yabin Duan. The activity, protein, and mRNA expression of CYP2E1 and CYP3A1 in rats after exposure to acute and chronic high altitude hypoxia. High Alt Med Biol 15:491–496, 2014.—The effects of exposure to acute and chronic high altitude hypoxia on the activity and expression of CYP2E1 and CYP3A1 were examined in rats. Rats were divided into low altitude (LA, 400 m), acute moderate altitude hypoxia (AMH, 2800 m), chronic moderate altitude hypoxia (CMH, 2800 m), acute high altitude hypoxia (AHH, 4300 m), and chronic high altitude hypoxia groups (CHH, 4300 m). Probe drugs were administrated orally to all five groups. Then the serum concentration of probe drug and its metabolite was determined by RP-HPLC. The activity of CYP2E1 and CYP3A1 was evaluated using the ratio of the metabolite to chlorzoxazone and testosterone, respectively. ELISA and real-time PCR were used to analyze the protein and mRNA expression of CYP2E1 and CYP3A1 in liver microsomes, respectively. Chronic high altitude hypoxia caused significant decreases in the activity and protein and mRNA expression of rat CYP2E1 and CYP3A1 in vivo. Acute high altitude hypoxia was not found to change the activity, protein or mRNA expression of rat CYP2E1 or CYP3A1. This study showed significant changes in the activity and protein and mRNA expression of CYP2E1 or CYP3A1 in rats after exposure to chronic high altitude hypoxia.
Introduction
T
Hypoxia is a frequent serious complication of a number of common diseases, and changes in the activity of drug-metabolizing enzymes related to this condition may be of clinical importance. Early in vivo and in vitro studies supported the conclusion that acute hypoxia reduces the activity of several biotransformation pathways (Jones, 1981). Modulation of cytochrome P450 by hypoxia was confirmed in vivo in animals with acute moderate hypoxia in which the clearance of theophylline was reduced (Letarte et al., 1984). Subchronic hypoxia was also found to reduce the activity and expression of enzymes involved in the biotransformation of drugs (Shan et al., 1992). However, animal studies measuring the effect of hypoxia on the activity or levels of P450 enzymes in the liver have yielded conflicting results. Some studies have shown that hypoxia does not impair metabolic processes mediated by cytochrome P450 (Streit et al., 2005; Saunier et al.,1987).
The environmental characteristics at high altitudes include, but are not limited to, decreased ambient oxygen tension, increased solar radiation, extreme diurnal ranges in temperature, arid climate, poor soil quality, and hypoxia that can affect the human life and activity considerably (Zhang et al., 1984). Living and exercising at high altitudes can lead to altitude sickness, which can range from the mild symptoms of acute and chronic mountain sickness to the potentially fatal high altitude pulmonary edema and cerebral edema (Duplain et al., 2007). Immediately upon exposure to high altitudes, hypoxia stimulates ventilation and causes respiratory alkalosis, which is subsequently counteracted by increased bicarbonate excretion in the kidneys (Mairbaurl et al., 1994). The increase in plasma pH depends on altitude and duration of stay (Sutton et al., 1988). Circulatory adjustments to hypoxia include the dilatation of coronary and cerebral vessels and the vasoconstriction in skeletal muscle, which induce the redistribution of blood flow to the most metabolically active systems (Heistad et al., 1980; Rowell et al., 1984; Wolff et al., 2000). However, these adjustments cannot prevent a decrease in arterial oxygen (O2). For this reason, O2 diffusion from blood into tissues is decreased.
The prevention and treatment of high altitude illness (Gallagher et al., 2004; Luks et al., 2008) and pharmacokinetic differences in the processing of drugs such as sulfamethoxazole, lithium, acetazolamide, and meperidine at high altitudes have been studied (Arancibia et al., 2003; Li et al., 2009; 2012). These studies found significant changes in the disposition of drugs in healthy subjects after either acute or chronic exposure to high altitude. The few previous studies on rabbits and rats under hypoxia conditions showed a decrease in the expression of CYP1A,1A2, 2B4, 2C5, and 2C16 and increased expression of CYP3A6 after acute exposure to hypoxia induced by a chamber with a Fi
However, there have been few reports dealing with changes in the activity and expression of cytochrome P450 related to the environmental characteristics at high altitudes so far. The effects of exposure to acute and chronic high altitude hypoxia on the activity and expression of CYP1A2, CYP2D6, CYP2C9, CYP2C19, and NAT2 in rats were investigated in a recent study by the present team (Li et al., 2014). However, there have been no reports dealing with changes in the activity and expression of CYP2E1 and CYP3A1 attributable to the environmental characteristics of high altitude areas so far. The present study was designed to investigate the regulation of CYP2E1 and CYP3A1 by acute and chronic hypoxia in rats in high altitude environments. To this purpose, the effects of acute and chronic hypoxia on the activity of CYP2E1 and CYP3A1 and the protein and mRNA expression of two drug metabolizing enzymes were assessed at altitudes of 2800 and 4300 m
Methods
Materials and reagents
The following chemicals were purchased from Sigma-Aldrich Chemical Company (St. Louis, MO, U.S.): 6-hydroxy chlorzoxazone (HCZX), testosterone (TS), bovine serum albumin (BSA), glycerol, glycine, HEPES, β-NADPH, phenol, TRIS, PMSF, potassium dihydrogen phosphate, dipotassium hydrogen phosphate, sodium dodecyl sulfate (SDS), and sodium potassium tartrate. 6β-Hydroxy testosterone (HTS) was purchased from Toronto Research Chemicals. Inc. (Toronto, Canada). Chlorzoxazone (CZX) was purchased from Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China). Rat CYP2E1 and CYP3A1 ELISA Kits were purchased from R&D Systems (Minneapolis, MN, U.S.). HPLC-grade methanol and acetic acid were both purchased from Shandong Yuwang Company, Inc. (Jinan, China). All other chemicals and solvents were obtained from commercial sources at the highest grade of purity available. RNAiso Plus and PrimeScript™ RT Reagent were obtained from Takara (Kyoto, Japan). The primers used in real-time PCR were synthesized by Takara.
Animals and treatment
Sprague Dawley SPF rats (180–220 g, certificate No.: 2007-001) of both sexes were obtained from the laboratory animal center of Xi'an Jiaotong University Medical College (Xi'an, China). All experimental procedures were in strict accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Fifty clean SD rats were randomly divided into five groups by weight, and every group had five male and five female rats. The low altitude group (LA) included healthy rats living at an altitude of ≈400 m in the city of Xi'an in northwest China's Shanxi Province. The acute moderate altitude hypoxia group (AMH) and the chronic moderate altitude hypoxia group (CMH) comprised healthy rats living at low altitudes, but they received a 24-h acute exposure and a 30-day chronic exposure to moderate altitudes prior to drug administration. The rats in these two groups were transported by bus to Gonghe County, which is in northwestern China's Qinghai Province, to an altitude of 2800 m. They arrived at 9:00
The examination of the low altitude group was performed at the Xi'an Traffic University Medical College, and the examination of the moderate and high altitude hypoxia groups in Gonghe County Hospital and Maduo County Hospital, respectively.
To confirm the significance of exposure to high altitudes, capillary oxygen saturation (Sc
ELISA analysis of EPO, HIF-1α, IL-β, and IFN-γ
EPO, HIF-1α, IL-β, and IFN-γ in the serum were assayed with rat ELISA kits (all from R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions.
Measurement of rat CYP2E1 and CYP3A1 activity
CYP2E1 and CYP3A1 activities in rat liver microsomes were determined using the previously reported “cocktail” method (He et al., 2007). Chlorzoxazone and testosterone served as probe substrates of CYP2E1 and CYP3A1, respectively, and two drugs were selected to evaluate the activity of CYP2E1 and CYP3A1. After an overnight fast of ≥12 h (with water allowed ad libitum), rats from LA, AMH, CMH, AHH, and CHH were given orally probe drugs. To evaluate the activity of CYP2E1, rats were given CZS orally at a dose of 15 mg/kg. The blood samples were collected 4 h after the administration of study drugs, and the serum concentration of CZX and metabolite HCZX was determined using RP-HPLC. The activity of CYP2E1 was evaluated by the ratio of HCZX to CZX. To evaluate the activity of CYP3A1, rats were given TS orally at a dose of 25 mg/kg. The blood samples were collected 6 h after administration of the study drug, and the serum concentration of TS and metabolite HTS was determined using RP-HPLC. The activity of CYP3A1 was evaluated using the ratio of TS to HTS.
Preparation of rat hepatic microsomes
The liver microsomes were prepared using differential centrifugation (Yasui et al., 2002). Liver samples were thawed and weighed, and two volumes of ice-cold homogenization medium (50 mmol/L Tris–HCl buffer at pH 7.4 containing 0.25 M sucrose) were then added. The tissue was chopped using scissors and homogenized with an automatic homogenizer at 500 rpm (IKA T8 Labortechnik, Germany). The resultant homogenates were transferred to clean centrifuge tubes and centrifuged at 10,000 g for 30 min at 4°C using a TGL-16B centrifuge (Anting Scientific Instrument Factory, Shanghai, China). The supernatant was collected and centrifuged at 100,000 g for 80 min at 4°C using an Optima MAX-XP ultracentrifuge (Beckman Coulter Inc., U.S.). The microsomal pellet was resuspended with homogenization medium. Hepatic microsomal suspensions (0.5 mL) were aliquoted into Eppendorf tubes and stored at −80°C until use.
ELISA analysis of CYP2E1 and CYP3A1 protein
To establish a bland well without any solution, 50 μL of standard and 40 μL of sample diluted solution were added to standard and sample well with a biotin-conjugated antibody specific to cytochrome P450, respectively. Then 10 μL of sample was added to the sample well. Avidin conjugated to horseradish peroxidase (HRP) was then added to each microplate well and incubated for 30 min at 37°C, after which liquid was removed completely, to fill each well with 200 μL of wash buffer, and stay for 10 sec and spinning. The process was repeated for a total of five washes, and any remaining wash buffer was removed by aspirating of decanting after the last wash. Then 50 μL of substrate A and 50 μL of substrate B were added to each well, mixed well, and incubated for 30 min at 37°C. The enzyme–substrate reaction was terminated by the addition of sulfuric acid solution and the color change measured spectrophotometrically at a wavelength of 450 nm. The concentration of CYP2E1 and CYP3A1 in the samples was then determined by comparing the O.D. of the samples to the standard curve.
RNA isolation and real-time PCR analysis of CYP2E1 and CYP3A1 mRNA
Rats from five groups were killed and their livers were immediately excised. A small portion of liver from the left lobe was snap-frozen in liquid nitrogen and stored at −80°C before extraction of total RNA. Approximately, 100–200 mg of liver tissue was homogenized and total RNA was isolated with the Trizol reagent. The quality of RNA solutions was determined using an ultraviolet light spectrophotometer. Primers for rat CYP2E1 and CYP3A1 and the reaction system were designed by Major Bio Bio-pharm Technology Co., Ltd (Shanghai, China). All PCR reactions were carried out using a SYBR® Premix Ex TaqTM Kit (Takara) in accordance with the manufacturer's instructions. Amplification was performed in PCR-capillaries on a Light Cycler 2.0 Real Time Detection System (Roche, U.S.). Amplification of predenatured products was conducted at 94°C for 60 sec, followed by 45 cycles at 95°C for 30 sec, 58°C for 30 sec and 72°C for 30 sec, 95°C for 10 sec, 65°C for 45 sec, and 40°C for 60 sec. Fold induction values were calculated using to the equation 2-ΔΔCt, where Δ Ct represents the differences in cycle threshold numbers between the target gene and the control gene β-actin, and ΔΔ Ct represents the relative change in the differences between control and treatment groups.
CYP2E1 primer sequence:
5′- GAACTGAGACCACCAGCACAA -3′ (forward primer)
5'- GATCTCATGCACCACAGCATC -3′ (reverse primer)
5' - CTTGGCCCAATAACCCTGTCAAT -3′ (probe)
CYP3A1 primer sequence:
5′- TTCTGTGCAGAAGCATCGAGT -3′ (forward primer)
5′- GACAAAGGAAAGTGTGCTGCT -3′ (reverse primer)
5′- ATGGAGATCACAGCCCAGTCAATC -3′ (probe)
β-actin primer sequence:
5′- CAGGTCATCACTATCGGCAAT -3′ (forward primer)
5′- TGGCATAGAGGTCTTTACGGA -3′ (reverse primer)
5′- CAGCCTTCCTTCCTGGGTATGG -3′ (probe)
Statistical analysis
Data were expressed as the mean±SEM from these independent experiments. Statistical analysis was performed using the Statistical Package for Social Sciences (SPSS, version 13.0) for Windows. Multiple comparisons between more than two groups were performed using one-way analysis of variance (ANOVA). P values lower than 0.05 were considered significant.
Results
High altitude hypoxia and the expression of EPO, HIF-1α, IL-β, and IFN-γ
The concentrations of EPO, HIF-1α, IL-β, and IFN-γ were significantly higher in rats exposed to chronic high altitude hypoxia. They were 231.6, 44.1, 123.9, and 83.8% higher in the CHH group than in the LA group, respectively (Fig. 1). The concentrations in other groups were not significant different from the LA group.
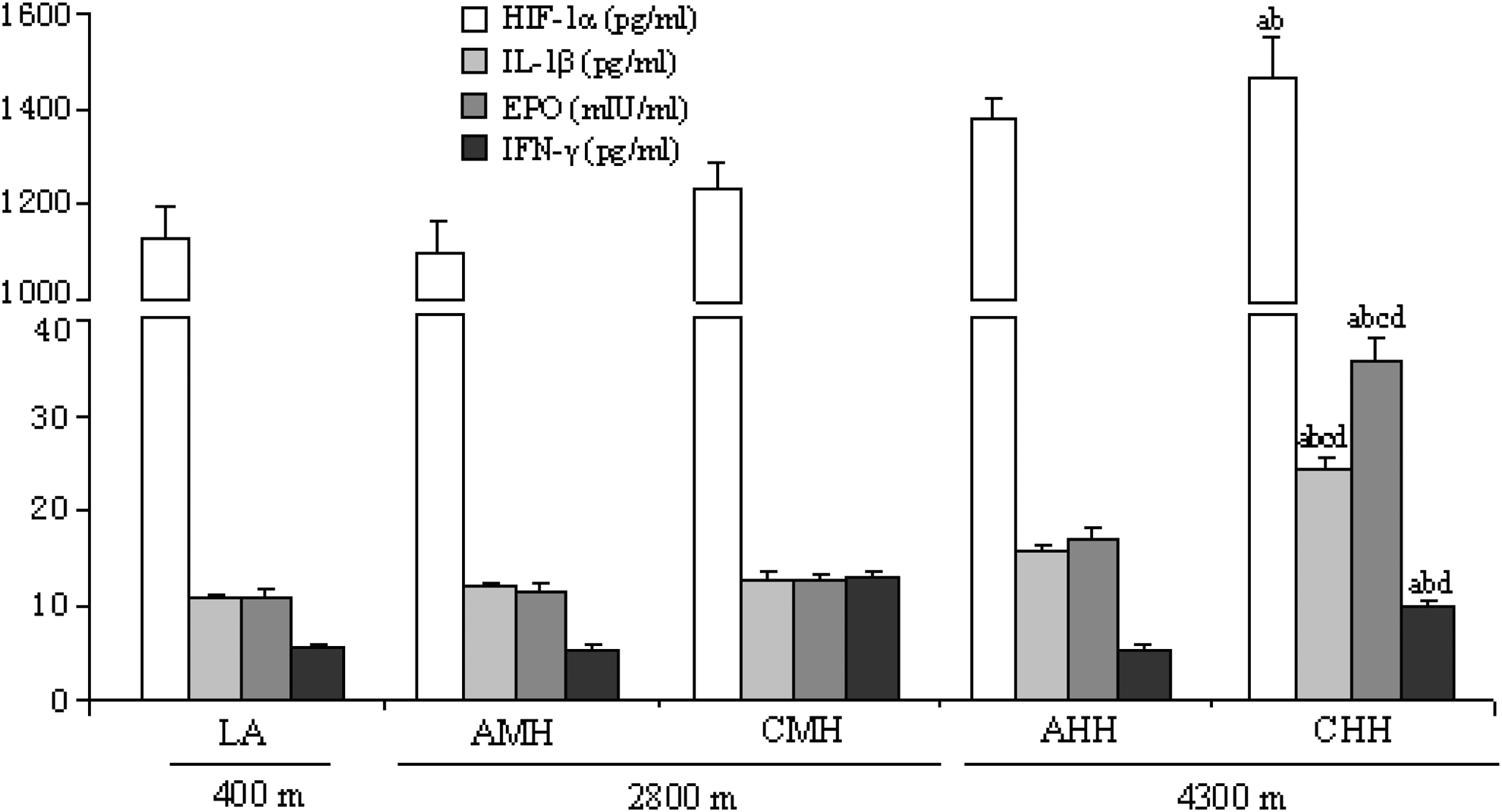
Concentrations of HIF-1α, IL-β, EPO, and IFN-γ in rats in the LA, AMH, CMH, AHH, and CHH groups. AHH: acute high altitude hypoxia; AMH: acute moderate altitude hypoxia; CHH: chronic high altitude hypoxia; CMH: chronic moderate altitude hypoxia; LA: low altitude. n=10, mean±SEM. ap<0.05 vs. LA; bp<0.05 vs. AMH; cp<0.05 vs. CMH; dp<0.05 vs. AHH.
High altitude hypoxia and CYP2E1 and CYP3A1 activity
There was significantly less rat CYP2E1 activity in rats exposed to chronic high altitude hypoxia. It was 60.2% and 44.2% lower in CMH and CHH, respectively, than in LA, (Fig. 2: CYP2E1). The activity of CYP2E1 in AMH and AHH was not significantly different from LA (Fig. 2: CYP2E1). There was significantly less CYP3A1 activity by 43.2% and 50.1% in CMH and CHH, respectively, than in the LA group. Activity levels in the AMH and AHH groups were not significant different from those of the LA group (Fig. 2: CYP3A1).
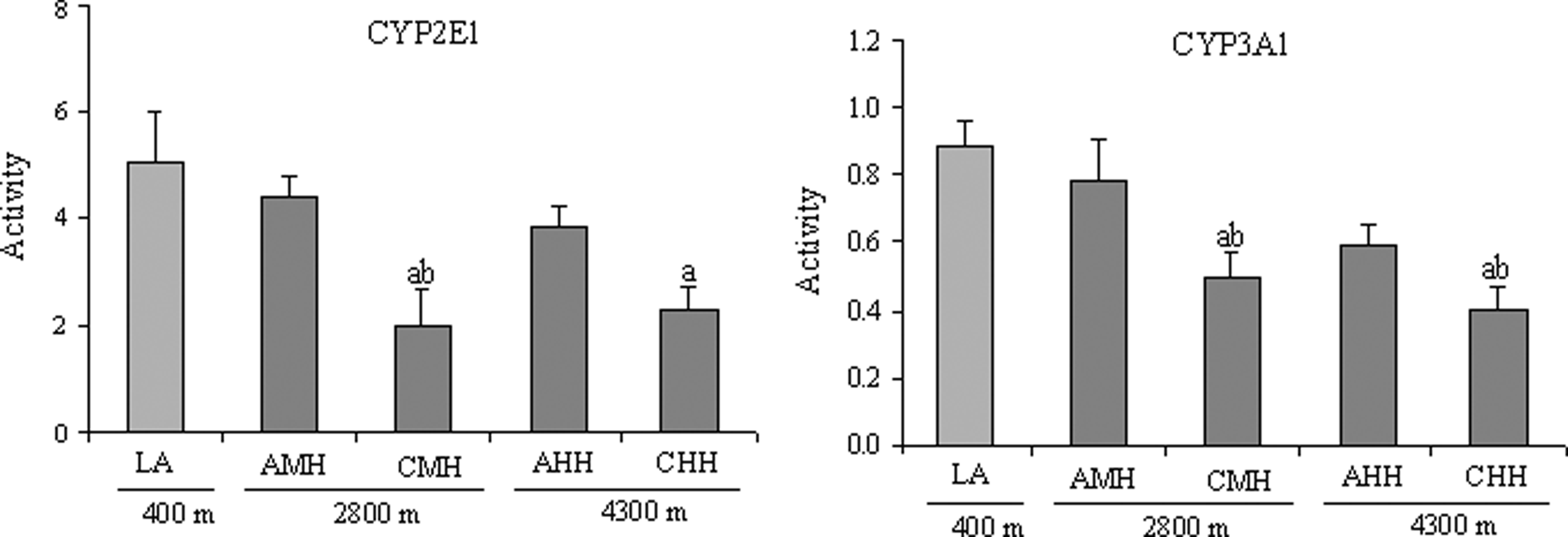
Activity of CYP2E1 and CYP3A1 in rats in the LA, AMH, CMH, AHH, and CHH groups. AHH: acute high altitude hypoxia; AMH: acute moderate altitude hypoxia; CHH: chronic high altitude hypoxia; CMH: chronic moderate altitude hypoxia; LA: low altitude. n=10, mean±SEM. ap<0.05 vs. LA; bp<0.05 vs. AMH.
High altitude hypoxia and the expression of CYP2E1 and CYP3A1 proteins
There was significantly less CYP2E1 and CYP3A1 protein expression in rats exposed to chronic high altitude hypoxia, and exposure to acute high altitude hypoxia showed no significant effect on the protein expression of two drug metabolizing enzymes. The protein expression of CYP2E1 was significantly lower in CHH decreased by 33.9% and 35.5%, than in with LA and AHH, respectively (Fig. 3). Other groups showed no significant differences from the LA group. The protein expression of CYP3A1 was 39.7% and 27.5% lower in CMH and CHH, respectively, than in the LA group. AMH and AHH showed no significant differences from the LA group (Fig. 3).

Protein expression of CYP2E1 and CYP3A1 in rats in the LA, AMH, CMH, AHH, and CHH groups. AHH: acute high altitude hypoxia; AMH: acute moderate altitude hypoxia; CHH: chronic high altitude hypoxia; CMH: chronic moderate altitude hypoxia; LA: low altitude. n=10, mean±SEM. ap<0.05 vs. LA; bp<0.05 vs. AMH; cp<0.05 vs. CMH; dp<0.05 vs. AHH
High altitude hypoxia and the expression of CYP2E1 and CYP3A1 mRNA
There was significantly less expression of CYP2E1 and CYP3A1 mRNA in rats exposed to chronic high altitude hypoxia, and exposure to acute high altitude hypoxia was found to have no significant effect on the mRNA expression of two drug metabolizing enzymes. The mRNA expression of CYP2E1 was significantly lower in CMH and CHH by 56.0% and 52.0%, respectively, than in the LA group. The mRNA expression of CYP2E1 was 50.9% lower in CMH than in AMH, and 54.5% lower in CHH than in AHH (Fig. 4). The mRNA expression of CYP3A1 was significantly lower by 31.1% and 33.7% in CMH and CHH, respectively, than in LA. The AMH and AHH groups were not significantly different from the LA group (Fig. 4).
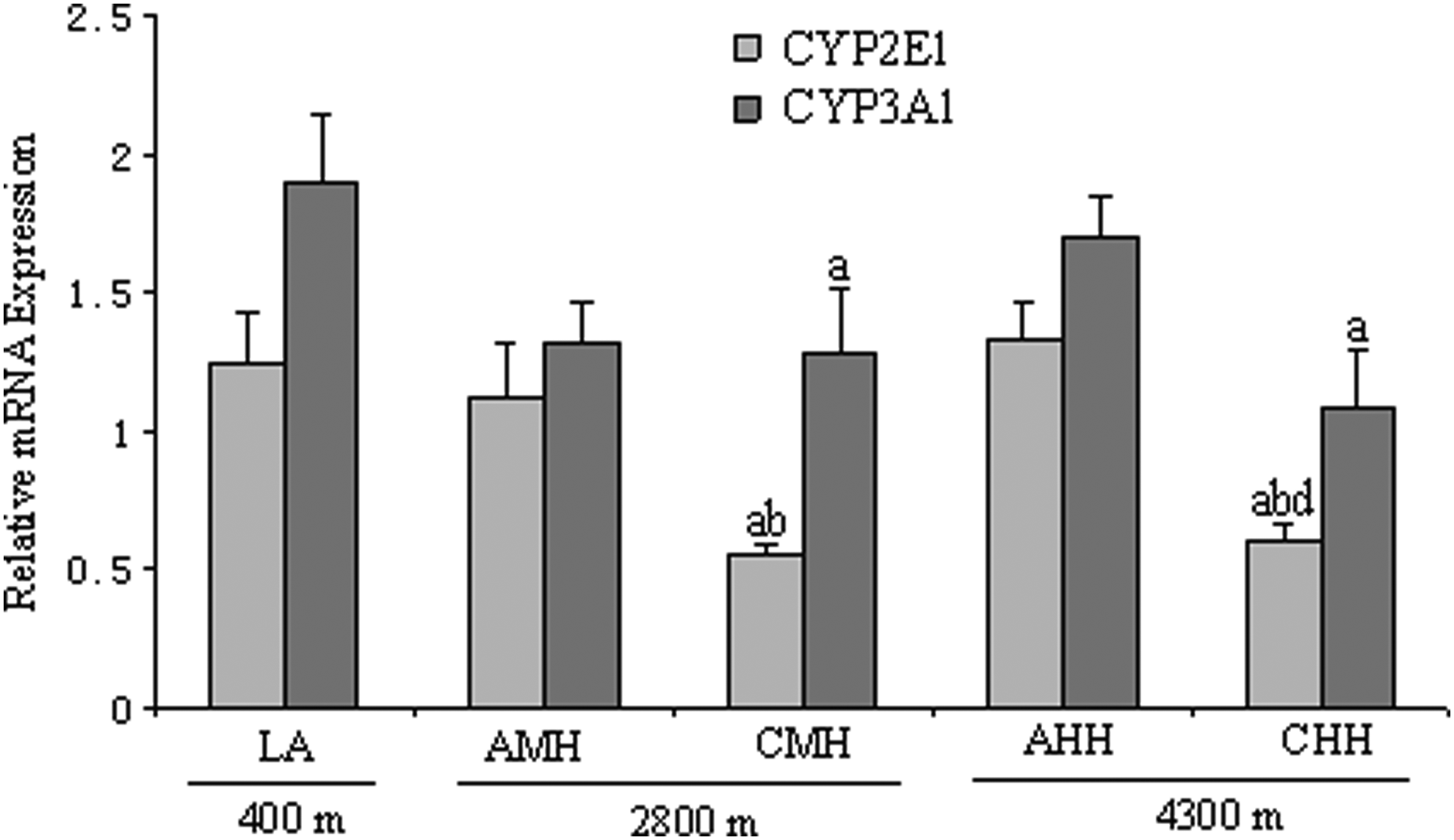
mRNA expression of CYP2E1 and CYP3A1 in rats in the LA, AMH, CMH, AHH, and CHH groups. AHH: acute high altitude hypoxia; AMH: acute moderate altitude hypoxia; CHH: chronic high altitude hypoxia; CMH: chronic moderate altitude hypoxia; LA: low altitude. n=10, mean±SEM. ap<0.05 vs. LA; bp<0.05 vs. AMH; dp<0.05 vs. AHH.
Discussion
The present study demonstrated that the activity, protein, and mRNA expression of CYP2E1 and CYP3A1 were significantly lower in rats exposed to chronic high altitude hypoxia, but acute high altitude hypoxia did not affect the activity, protein, and mRNA expression of two drug-metabolizing enzymes.
A previous study focused on acute moderate hypoxia and the effect of hypoxia on the activity and expression of cytochrome P450. Fradette et al. (2002) demonstrated that the expression of CYP1A and 1A2 was significantly lower in rabbits subjected to a Fi
There have been no reports addressing changes in the activity and expression of CYP2E1 and CYP3A1 in response to the environmental characteristics of high altitude environments. To further study the effects of hypoxia on the activity and expression of cytochrome P450, changes in the activity and expression of CYP2E1 and CYP3A1 was examined in rats exposed to acute and chronic high altitude hypoxia. Data suggested that chronic high altitude hypoxia could cause significant decreases in the activity, protein, and mRNA expression of rat CYP2EA and CYP3A1 in vivo, and acute high altitude hypoxia did not change the activity, protein, and mRNA expression of two drug-metabolizing enzymes.
Several mechanisms may have contributed to downregulation of CYP1A1 and 1A2 under hypoxic conditions. The effects of hypoxia on P450 resemble those elicited by an acute local inflammatory reaction (Morgan et al., 1997). Bleau et al. (2000) found IL-6 to be the serum mediator responsible for the decrease in activity of P450 isoforms in rabbits with an inflammatory reaction and found IL-1β, tumor necrosis factor-α, and IFN-γ to have minor roles. Hypoxia prompts the release of numerous cytokines, such as IL, TNF-α, IFN-γ, EPO, and HIF-1α (Naldini et al., 1997; Lacombe et al., 1999; Semenza et al., 1999). Fradette et al. (2002; 2007) identified the serum mediators implicated in the decrease in activity and expression of CYP1A1, 1A2, and 3A6 as IFN-γ, IL-1β,IL-2, and EPO. HIF-1α was found to be partially responsible for the increase in CYP3A6 expression (Fradette et al., 2003). Though these mediators can induce changes in activity and expression of CYP1A1, 1A2, and 3A6, it is still unclear whether EPO, HIF-1α, IL-β, and IFN-γ can mediate the changes in the activity and expression of CYP2E1 and CYP3A1. The current study showed there to be significantly more expression of EPO, HIF-1α, IL-β, and IFN-γ in rats exposed to chronic high altitude hypoxia, by 231.6%, 44.1%, 123.9%, and 83.8%, respectively, relative to control rats. However, further study is needed to determine whether these mediators are responsible for the decrease in the activity and expression of CYP2E1 and CYP3A1 in high altitude hypoxia environments.
In conclusion, the present study demonstrated that chronic high altitude hypoxia decreased the activity, protein expression, and mRNA expression of CYP2E1 and CYP3A1. The result of these changes on drug kinetics of drugs will depend upon the isoforms involved in their biotransformation. A previous study demonstrated that high altitude hypoxia decreases the activity and expression of N-acetyltransferase 2 (NAT2) (Yang et al., 2013). It also showed that area under the concentration-time curve (AUC) of sulfamethoxazole as metabolized mainly by NAT2 was 17.8% higher and clearance was 17.8% lower in healthy Chinese male volunteers after 16 h of exposure to high altitudes (3780 m) than in control volunteers at low altitudes (400 m) (Li et al., 2009). The AUC of substrates of CYP2E1 and CYP3A1 is here predicted to be bigger because of increased absorption and decreased first pass and renal clearance under high altitude hypoxic environmental conditions. The results of this study may have important clinical implications. It is here suggested that patients living at or traveling to high altitude should be closely monitored, and the dosages of some drugs metabolized by CYP2E1 or CYP3A1 should be reduced.
Conclusion
Acute high altitude hypoxia did not change the activity, protein expression, or mRNA expression of rat CYP2E1 or CYP3A1, but chronic high altitude hypoxia did. This study showed significant changes in the activity, protein expression, and mRNA expression of CYP2E1 and CYP3A1 in rats exposed to chronic high altitude hypoxia.
Footnotes
Acknowledgments
Author Disclosure Statement
No competing financial interests exist. This work was financially supported by the National Natural Science Foundation of China (N