
Abstract
Abstract
Avnstorp, Magnus B., Peter Rasmussen, Patrice Brassard, Thomas Seifert, Morten Overgaard, Peter Krustrup, Niels H. Secher, and Nikolai B. Nordsborg. Cerebral water and ion balance remains stable when humans are exposed to acute hypoxic exercise. High Alt Med Biol 16:18–25, 2015.—
Methods:
On two separate days, in random order, 30 min cycling exercise was performed in either hypoxia (10% O2) or normoxia at an intensity that was exhaustive in the hypoxic trial (∼120 W; n=9).
Results:
Exercise in hypoxia affected muscle metabolism, as evidenced by higher (p<0.05) leg lactate release at 7.5 min and a continuous decline in arterial pH (p<0.001) that was not observed in normoxia. Middle cerebral artery flow velocity increased (p<0.01) with exercise under both circumstances. No cerebral net exchange of Na+ or K+ was evident. Likewise, no significant net-exchange of water over the brain was demonstrated and the arterial and jugular venous hemoglobin concentrations were similar.
Conclusion:
Challenging exercise in hypoxia for 30 min affected muscle metabolism and increased an index of cerebral blood flow, but cerebral net water and ion homeostasis remained stable. Thus, although AMS develops within hours and may be related to exercise-induced disturbance of cerebral ion and water balance, such changes are not detectable when subjects are exposed to acute 30 min maximal exercise in hypoxia.
Introduction
A
Water and ion homeostasis are closely linked. Hypoxic exposure may affect brain water balance by altering cerebral ion homeostasis, which depends on systemic ion homeostasis, because K+ crosses the blood brain barrier (Keep et al., 1995) and brain tissue can take up K+ from the cerebrospinal fluid (Bradbury et al., 1972). Furthermore, plasma Na+ affects brain water content (Overgaard-Steensen et al., 2010). Expression of Na+/H+ exchangers (NHE1) and Na+, K+, Cl- co-transporters (NKCC1) in the cerebral microvascular endothelial cells (Pedersen et al., 2006; Kahle et al., 2009) allows for exchange of H+ and Cl- in addition to Na+ and K+ ions. Also the mono-carboxylate transporters MCT1, MCT 2 and MCT4 (Halestrap and Price, 1999; Rafiki et al., 2003; Chiry et al., 2006) allow for exchange of metabolites and ions between blood and brain by gradient dependent co-transport. The H+/La- transport system is important since lactate oxidation may cover up to 25% of the brain's carbohydrate oxidation during exercise (van Hall et al., 2009). Additionally, NKCC1 and MCT1 are co-transporters of ions and water (Hamann et al., 2003, 2010). Thus, exercise and/or hypoxia-induced disturbance of systemic ion homeostasis could affect cerebral water balance and be of importance for development of AMS.
Plasma K+ and Na+ are elevated at high exercise intensities (Lindinger et al., 1992; Hallen et al., 1994) and even more so when graded exercise is performed in hypoxia (Lundby and van Hall, 2002; Nordsborg et al., 2010; Rasmussen et al., 2010). The exercise-induced increase in plasma ion concentrations results mainly from muscular release (Sejersted and Sjogaard, 2000) but the higher ion concentrations with hypoxic exposure is related to pronounced exercise-induced reduction of plasma volume during graded exercise (Nordsborg et al., 2010). However, most recreational activities at high altitude are not graded but rather performed at a constant load. However, there has been no evaluation of muscular ion homeostasis during exhaustive constant-load exercise in hypoxia.
In addition to challenged ion homeostasis, hypoxia affects cerebral blood flow, which increases by ∼15% during exercise in normoxia (Ahlborg et al., 1986; Secher et al., 2008) and by ∼25% in hypoxia (Imray et al., 2005; Overgaard et al., 2012). Increased cerebral blood flow could exaggerate filtration by enhanced perfusion pressure or by increased total ion-influxes. However, cerebral water and ion balance with exercise and hypoxic exposure has not been determined.
The aim of this study was to investigate the hypotheses that 1) 30 min of exercise at an exhaustive intensity in hypoxia (Fi
Methods
Five men and four women (age: 26±6 yrs., body weight: 68±8 kg, height: 167±11 cm; mean±SD), who were habitually active with no medical issues participated in the study. The subjects provided oral and written informed consent to the study as approved by the Ethical Committee of Copenhagen and Frederiksberg (KF-01287471). Results from the present study, including cerebral lactate fluxes, have been published previously (Overgaard et al,. 2012).
Experimental design
Four to seven days prior to the study, subjects were familiarized to the protocol by a 30-min cycling time-trial in normobaric hypoxia (Fi
The two experimental days were separated by 14 days using a randomized and counter-balanced design. On the day before the experimental days, subjects were asked to refrain from exhaustive exercise and from intake of caffeine (tea/coffee) and to fast 1 h prior to reporting to the laboratory at 8
After instrumentation, subjects rested supine for 30 min after which they exercised at ∼120 W for 30 min in normoxia (Fi
Blood was sampled in pre-heparinized syringes, purged of any atmospheric content and kept anaerobic on ice water and analyzed within 30 min for K+, Na+, Ca2+ lactate, pH, O2 saturation, pO2, and hemoglobin (Hb) content (ABL 725, Radiometer, Copenhagen, Denmark). Femoral venous blood flow was measured by thermodilution (Andersen and Saltin, 1985). Middle cerebral artery flow velocity (MCA Vmean) was evaluated by transcranial ultrasound Doppler (TCD, Transcan, EME, Überlingen, Germany) insonated through the trans-temporal bone window. Depending on the position with the best signal-noise ratio, the proximal part of MCA was insonated at a depth of 40–60 mm and the probe was secured with a headband. The position of the probe was noted to ensure identical placement for the two trials. The MCA was chosen because it supplies the dominant part of the cortex including the motor area. Also changes in flow velocity in the MCA agree well with flow in the internal carotid artery during exercise (Sato et al., 2012).
Calculations
To calculate net-exchange of ions, it was assumed that the amount of Hb entering and leaving the brain was identical. Thus, the net exchange across the brain or leg is the difference between venous plasma ion outflow and arterial ion inflow:
with Q being the blood flow in either MCA or the femoral vein for calculations of head and leg balance, respectively. For this calculation it is assumed that no net exchange of ions takes place between plasma and red blood cells (Juel et al., 1999).
Brain fluid balance was venous minus arterial plasma flow. Cerebral arterial and venous plasma flow were calculated from the arterial blood flow, also assuming that an equal amount of Hb enters and leaves the brain. It was further assumed that a change in hematocrit represents a net loss or uptake of plasma from the bloodstream and not a fluid exchange between plasma and red blood cells. Thus, brain water balance=MCA Vmean×([Hb]a/[Hb]v)×(1-Hctv. jugularis) – MCA Vmean×(1-Hcta).
Statistics
Two-way analysis of variance for repeated measures was performed. Fixed factors were trial (Normoxia; Hypoxia) and time. We evaluated the net release or uptake of ions across the brain between 3–30 min of exercise by calculating a weighted average of all obtained measurements. It was evaluated whether this net release or uptake was different from zero by a one-sample t-test. No corrections were made for these t-tests as the risk of failure to reject a false null-hypothesis (Type II error, i.e. u=0), appeared more evident than a type I error, due to the low number of included subjects that is related to the invasive nature of the study. The level of significance was set at p<0.05 and results are reported as mean and 95% confidence intervals.
Results
Blood pH
At rest, arterial pH was higher (p<0.001) in hypoxia than in normoxia (Table 1) and at 3 min of exercise (7.43 [7.41; 7.45] vs. 7.38 [7.37; 7.39]). Arterial pH, however, declined (p<0.01) during exercise in hypoxia but remained stable in normoxia. Thus, after 30 min of exercise, arterial pH was lower (p<0.001) in hypoxia than in normoxia (7.32 [7.25; 7.39] vs. 7.40 [7.38; 7.41]).
Hb, hemoglobin concentration (mM); Hct, hematocrit; pO2, partial oxygen pressure (mmHg); pCO2, partial carbon dioxide pressure (mmHg); sO2, oxygen saturation (%). n = 7, except for Hb where n = 6. Significant differences are indicated by one, two, or three symbols for p < 0.05; p < 0.01, and p < 0.001, respectively. #Different from the resting sample in the same situation; *Difference between the a. and v. sample; $Different from the corresponding sample in normoxia;
Leg O2 uptake and blood flow
During exercise leg blood flow was higher (P<0.05) in hypoxia than normoxia (5.6 [5.2; 5.9] vs. 3.9 [3.6; 4.1] L×min−1) and yet leg O2 uptake was similar (550 [313; 787] vs. 558 [400; 755] mL×min−1). Both leg blood flow and leg O2 uptake remained stable from 3 to 30 min in the two trials.
Leg lactate exchange
After 3 min of exercise, femoral venous [La-] was higher (p<0.01) in hypoxia than normoxia (6.0 [5.0; 6.9] vs. 2.0 [1.5; 2.5] mM) and after 30 min [La-] increased (p<0.01) to 12.3 [9.9; 14.6] mM in hypoxia but was only 1.3 [1.0; 1.7] mM in normoxia (Fig. 1). An interaction between trial and sample existed (p<0.05) for leg lactate exchange, which was higher (p<0.05) at 7.5 min in hypoxia than in normoxia (2.2 [−2.2; 6.9] vs. 0.1 [−0.4; 0.6] mmol×min−1) but after 20 min, the hypoxic value became negative and thereby lower (p<0.05) than in normoxia (−1.1 [−3.2; 1.1] vs. 0.0 [−0.3; 0.3] mmol×min−1). The net lactate exchange from 3–30 min of exercise was similar in hypoxia and normoxia (0.7 [−0.1; 1.6] vs. 0.1 [0.0; 0.1] mmol×min−1).
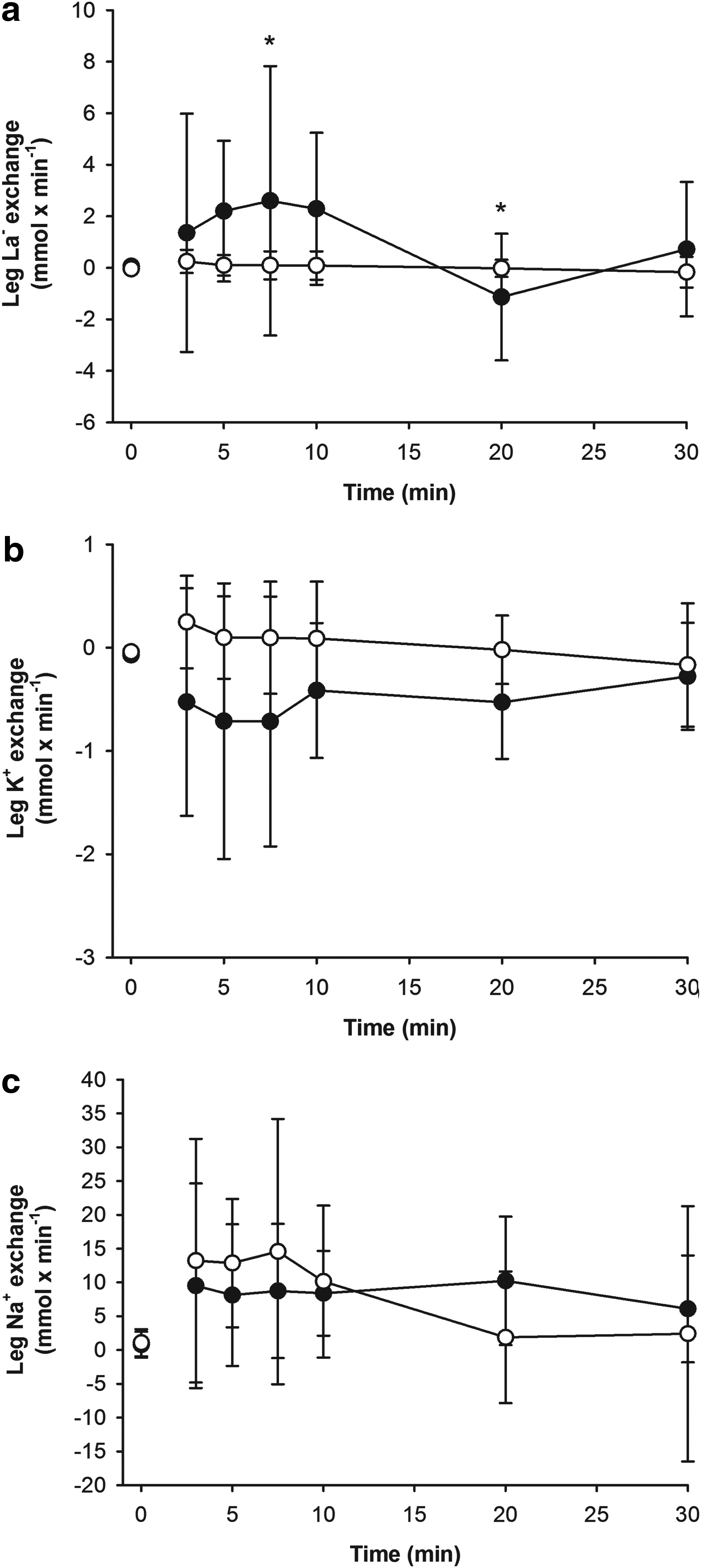
Exchange of lactate
Leg K+ exchange
At 3 min of exercise, femoral venous [K+] was similar in hypoxia and normoxia (4.1 [4.0; 4.3] vs. 4.3 [4.3; 4.4] mM) and did not change further (Fig. 1). Leg K+ exchange differed between conditions (p<0.01) but no time specific difference was detected by the post hoc test. Furthermore, on average there was no leg K+ exchange during exercise in either hypoxia or normoxia (−0.5 [−1.0; 0.0] vs. 0.1 [0.0; 0.1] mmol×min−1) but a tendency for a difference was detected (p=0.08) between conditions.
Leg Na+ exchange
Femoral venous [Na+] was higher (p<0.05) at 3 min in hypoxia than in normoxia (144 [143; 144] vs. 142 [141; 142] mM) and this difference persisted until the 20th min (Fig. 1). No change in femoral venous [Na+] was observed during exercise under the two conditions. Muscular Na+ exchange was similar in both conditions and the average net Na+ loss from 3 to 30 min was similar in hypoxia and normoxia (7 [2; 12] vs. 6 [4; 8] mmol×min−1).
Leg Ca2+ exchange
From the third minute of exercise, femoral venous [Ca2+] was similar in hypoxia and normoxia (1.18 [1.16; 1.21] vs. 1.22 [1.21; 1.22] mM). An average leg Ca2+ release (p<0.001) was observed in both hypoxia and normoxia (0.10 [0.07; 0.13] vs. 0.07 [0.05; 0.09] mmol×min−1) but with no difference between conditions.
Cerebral variables
Hemoglobin
At rest, arterial [Hb] was similar in hypoxia and normoxia (Table 1) and did not change during exercise (30 min: 8.4 [7.7; 9.2] vs. 8.7 [8.0; 9.3] mM). Likewise, at rest, jugular venous [Hb] was similar in hypoxia and normoxia and was not significantly altered by exercise (30 min: 8.5 [7.8; 9.2] vs. 8.7 [8.0; 9.4] mM). The arterial to jugular venous difference was affected by hypoxia (p<0.05) but no time specific differences were identified. At rest, mean values in hypoxia were slightly smaller than in normoxia (−0.1 [−0.2; 0.1] vs. 0.0 [−0.1; 0.1] mM) and that was also the case at 30 min (−0.1 [−0.3; 0.0] vs. 0.1 [−0.2; 0.4] mM) but no significant difference could be detected.
Cerebral water balance
Cerebral net water balance did not change from rest to exercise and it was not affected by hypoxia (Fig. 2). Accordingly, when quantifying cerebral water balance by assuming a resting cerebral blood flow of 46 ml×100 g−1×min−1 (Madsen et al., 1993) and a cerebral volume of 1300 mL (Leonard et al., 2008), no net uptake or release of water could be detected (−5 [−8; −3] ml×min−1 in hypoxia; −2 [−5; 0] mL×min−1 in normoxia).
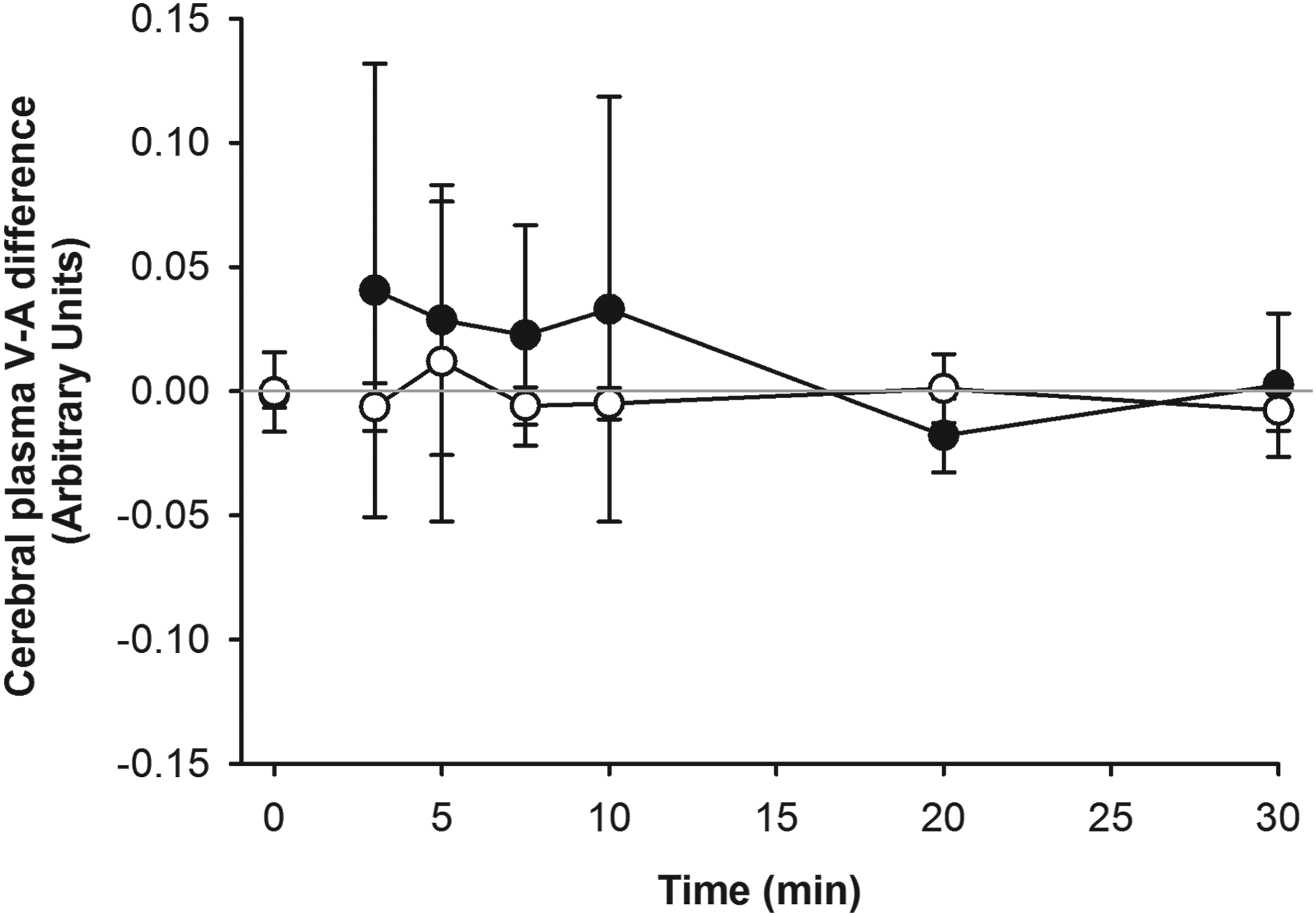
Difference between blood plasma inflow and outflow determined across the brain by sampling from a. brachialis and v. jugularis combined with measures of hematocrit, hemoglobin, and blood flow velocity in the middle cerebral artery. Exercise was performed for 30 min in normoxia (open circles) and severe hypoxia (Fi
Cerebral K+ exchange
Arterial [K+] was higher (p<0.01) during exercise compared to rest in both hypoxia and normoxia (at rest: 3.6 [3.5; 3.7] vs. 3.8 [3.7; 3.9] mM; at 30 min: 4.2 [4.0; 4.4] vs. 4.2 [4.0; 4.3] mM) but no difference between conditions was detected. Average cerebral K+ delivery during exercise in hypoxia was 26 [7; 45]% higher (p<0.05) than in normoxia because of increased (p<0.05) blood flow in hypoxia. No cerebral net K+ exchange was detected from 3 to 30 min of exercise and no difference was detected between conditions. In support, from 3 to 30 min there was no cerebral plasma K+ v-a difference in either hypoxia or in normoxia (0.0 [−0.2; 0.1] vs. 0.0 [0.0; 0.1] mM).
Cerebral Na+ exchange
Arterial [Na+] did not change significantly during exercise but was higher (p<0.01) in hypoxia than in normoxia although the post hoc test did not reveal any difference at a given time point. Specifically, at rest arterial [Na+] was similar in hypoxia and normoxia (139 [138; 140] vs. 139 [137; 140] mM) and that was also the case after 3 min (142 [141; 143] vs. 140 [139; 141] mM) and 30 min (142 [141; 143] vs. 140 [139; 142] mM). Brain Na+ influx in hypoxia was 30 [5; 55]% higher (p<0.01) than in normoxia. There was a different cerebral plasma Na+ v-a in both hypoxia or normoxia (0.4 [0.3; 0.6] mM, p<0.01 vs. 0.8 [0.5; 1.0] mM, p<0.001) but no net brain Na+ exchange could be detected.
Cerebral Ca2+ homeostasis
Arterial [Ca2+] did not change during exercise but was lower (p<0.01) in hypoxia than in normoxia at rest (1.16 [1.14; 1.18] vs. 1.19 [1.17; 1.21] mM; p<0.05) and at 3 min of exercise (1.17 [1.15; 1.19] vs. 1.20 [1.20; 1.21] mM; p<0.05). However, the brain Ca2+ influx in hypoxia was 21 [5; 36]% higher (p<0.001) than in normoxia. No cerebral Ca2+ exchange was detectable during exercise in hypoxia or normoxia.
Discussion
The major findings of this study were that exhaustive constant load exercise in severe hypoxia affects muscular metabolism as evidenced by a marked elevation of plasma [La-] and increased muscular La- release in the initial phase of exercise. On the other hand, muscular ion homeostasis was largely unaffected. Exercise in hypoxia, as compared to normoxia, caused increased brain influx of K+, Na+, and Ca2+, primarily due to increased cerebral blood flow. Additionally, La- influx was elevated due to an increased plasma concentration. Despite the exercise and hypoxia-induced challenges to cerebral ion homeostasis and water balance, no cerebral uptake or release of water was detected. This negative finding is supported by the similar [Hb] in the arterial and jugular venous blood both during exercise in hypoxia and normoxia. However, a trend towards a Hb arterial to venous negative difference was detected, indicating a small movements of water. Thus, the results demonstrate that the etiology of AMS should be sought in other mechanisms than muscle-induced disturbances of systemic ion homeostasis that acutely affects cerebral ion homeostasis with potential effect on cerebral water balance.
Cerebral water balance
We hypothesized that hypoxia-induced increase in brain blood flow, in combination with an exercise-induced elevation of arterial plasma K+, Na+, and La- concentrations would disturb cerebral water balance. However, the changes in brain blood flow may be underestimated as MCA diameter is likely to increase in hypoxia (Wilson et al., 2011; Ogoh et al., 2013) and further that MCA blood flow changes may not be representative for vessels such as the internal carotid artery and vertebral artery under these circumstances (Sato et al., 2011; Willie et al., 2012). However, neither exercise in normoxia nor in severe hypoxia affected the calculated cerebral water balance. In agreement with these observations, no difference was observed between brachial arterial and jugular venous [Hb]. Thus, acute alterations in systemic ion balance when healthy persons rest for 30 min and thereafter exercise for 30 min in 10% O2 appear not to induce development of cerebral edema. Obviously, the present results do not exclude that harmful cerebral water-shifts may occur with hypoxic exposure for more than 30 min of rest and 30 min of exercise. The tendency for a negative arterial to jugular venous Hb difference may indicate that quantitatively small movements of water manifested and could be of importance if the exposure time was prolonged.
Cerebral ion balance
In normoxia, exercise increased arterial plasma [K+] by approximately 15% relative to rest. Yet, despite increased cerebral K+ delivery, cerebral K+ balance was unaffected by exercise in normoxia. When comparing exercise in normoxia and hypoxia, arterial plasma [K+] was similar, but MCA Vmean was ∼25% higher in hypoxia. However, the hypoxia induced reduction of paCO2 may have caused MCA vasonstriction (Wilson et al., 2011; Ogoh et al., 2013; Coverdale et al., 2014) and thus reducing the estimated increase of cerebral blood flow in hypoxia. Because no difference in cerebral K+ balance was detected between exercise in normoxia and hypoxia, it appears that a flow-mediated increase in cerebral K+ delivery does not result in cerebral K+ uptake in humans during 30 min of exercise in acute hypoxia. Even if the hypoxia-induced increase of cerebral blood flow measurements were overestimated, it would not impact the estimation of possible differences in K+ flux in hypoxia vs. normoxia because the cerebral K+ a.-v. difference was not significantly different from 0.
Only modest changes in plasma [Na+] were observed and no effect on cerebral Na+ uptake or release could be detected. Thus, neither a moderate arterial [Na+] concentration change nor increased cerebral blood flow appear to alter cerebral Na+ balance. However, a small undetected cerebral release of Na+ may exist as indicated by the positive v.- a. difference in the first 10 min of normoxic exercise. The expression of Na+/H+ exchangers in the cerebral endothelial cells (Pedersen et al., 2006) could suggest that the increased jugular venous plasma H+ concentration from 3–7.5 min of exercise increased Na+ efflux from the brain. The present findings are in agreement with the increase in venous plasma Na+ during exercise are modest, reaching 2%–8% after intense exercise lasting from 30 sec to 60 min (Kowalchuk et al., 1988; Lindinger et al., 1992; Sostaric et al., 2006; Hew-Butler et al., 2008).
Arterial [Ca2+] was ∼2.5% reduced with hypoxic exposure and this difference persisted during the first 5 min of exercise. Despite the modest reduction in arterial [Ca2+], the ∼25% higher blood flow in hypoxia caused increased Ca2+ influx to the brain. However, no detectable change in net brain Ca2+ flux as a result of hypoxic exposure and exercise existed.
Leg ion balance
The increase in arterial and venous [K+] was established at the onset of exercise and maintained during exercise. The observation of similar leg K+ release during exercise in hypoxia and normoxia is in agreement with observations during incremental exercise (Nordsborg et al., 2010). A continuous leg Na+ release of ∼10 mmol×min−1 was evident from the exercising muscles. However, that did not result in a continuous plasma [Na+] increase or brain uptake of Na+. Thus, Na+ lost from the contracting leg muscles must be distributed to other tissues than the brain.
Limitations
Because no uptake or release of cerebral water, Ca2+, K+, or Na+ ions were detectable, it is of importance to consider potential methodological limitations. In the transition from rest to exercise, a power calculation estimates that the detectable difference in cerebral plasma balance is 21 ml×min−1 (∼8% of the resting level) and the detectable difference in jugular venous [Hb] is 0.11 mM (<2%). In these approximations, a resting cerebral blood flow of 46 ml×100 g−1×min−1 (Madsen et al., 1993) and a cerebral mass of 1300 g (Leonard et al., 2008) results in a cerebral blood flow at rest of 550 mL min−1.
As water shift from blood-plasma to the brain is expected to cause hemoconcentration from the arterial to jugular venous blood, the detection limit for Hb a.-v. difference was calculated and found to be 0.2 mM. This corresponds to the loss of 14 mL of water from one liter of blood passing the brain. When comparing exercise in normoxia and hypoxia, the detection limit for changes of cerebral water balance was ∼7% and no difference larger than 24 ml×min−1 is likely to have taken place. Likewise, a change in jugular venous [Hb] of ∼1.1 mM (∼11% of the resting level) would have been detected between conditions. However, these estimates also demonstrate that small movements of fluid may have occurred and could be of importance if exercise and hypoxic exposure is prolonged. The previous report of increase cerebral blood volume by 2 mL and brain parenchyma volume by 8 mL in a resting person after less than 1 h of hypoxic exposure (Dubowitz et al., 2009) may be related to a higher sensitivity of MRI at rest.
For cerebral K+ balance, a possible undetected difference from rest to exercise is 0.08 mmol×min−1 at 120 W in normoxia where arterial cerebral K+ delivery was 2.2±0.3 mmol×min−1 (∼4% of cerebral K+ influx). Cerebral K+ balance did not change more than 3% to 11% between conditions, corresponding to between 0.04 and 0.16 mmol×min−1, with arterial [K+] being 4.3 mM. Moreover, the detectable difference in K+ v-a difference between conditions was ∼5%. Thus, even small differences in either total plasma or K+ balance would have been detected.
Based on these considerations, the current study demonstrates that no substantial net-uptake or release of ions or water from the plasma compartment occurs during the brain passage under the investigated conditions. However, small undetected changes may have occurred with potential importance for AMS development over time. Also, it should be considered that the study hypothesis was based on observation of AMS symptoms developed when hypoxic exposure is combined with moderate (Roach et al., 2000) and intense (Wu et al., 2012) exercise at altitude. However, mild to moderate intensity exercise apparently does not worsen AMS (Schommer et al., 2012; Rupp et al., 2013). In the present study, a 30 min high-intensity exercise protocol was applied. Although 30 min is too short for AMS to develop, it appears that the combination of intense exercise and severe hypoxic exposure is a protocol expected to cause substantial changes of brain blood flow as well as plasma metabolite concentration. Longer duration exercise, as most often performed in real-life, would require reduced exercise intensity and thus likely reduce the net-water fluxes and ion-disturbances.
Conclusion
Muscle metabolism was affected by hypoxic exposure as evidenced by altered La- release. In contrast, muscular ion homeostasis was only slightly affected. Cerebral arterial Ca2+, Na+, and K+ delivery was elevated during exercise in acute severe hypoxia (Fi
Footnotes
Acknowledgments
Patrice Brassard was the recipient of a postdoctoral fellowship from the Fonds de recherche du Québec—Santé (FRQS) at the time of the study and is now a Junior 1 Research Scholar of the FRQS. Peter Rasmussen was supported by a grant from the Danish National Research Council (10-094110).
Author Disclosure Statement
No competing financial interests exist.