
Abstract
The human spleen can contract and transiently boost the blood with stored erythrocytes. We measured spleen volume and contraction during apneas in two groups, each containing 12 Caucasian participants (each 3 women): one group planning to summit Mt. Everest (8848 m; “Climbers”) and another trekking to Everest Base Camp (5300 m; “Trekkers”). Tests were done in Kathmandu (1370 m) 1–3 days after arrival, before the Climb/Trek. Age, height, weight, vital capacity, resting heart rate, and arterial oxygen saturation were similar between groups (not significant). After 15 minutes of sitting rest, all participants performed a 1-minute apnea and, after 2 minutes of rest, 1 maximal duration apnea was performed. Six of the climbers did a third apnea and hemoglobin concentration (Hb) was measured. Three axial spleen diameters were measured by ultrasonic imaging before and after the apneas for spleen volume calculation. Mean (standard deviation) baseline spleen volume was larger in Climbers [367 (181) mL] than in Trekkers [228 (70) mL; p = 0.022]. Spleen contraction occurred during apneas in both groups, with about twice the magnitude in Climbers. Three apneas in six of the Climbers resulted in a spleen volume reduction from 348 (145) to 202 (91) mL (p = 0.005) and an Hb elevation from 147.9 (13.1) to 153.3 (11.3) g/L (p = 0.024). Maximal apneic duration was longer in Climbers [88 (23) seconds vs. 67 (18) seconds in Trekkers; p = 0.023]. We concluded that a large spleen characterizes Climbers, suggesting that spleen function may be important for high-altitude climbing performance.
Introduction
The spleen is an organ that used to be associated mainly with immunological function in humans (Kruetzmann et al., 2003; Tarantino et al., 2011). While it is considered important in some mammals as a dynamic blood reservoir (Guntheroth and Mullins, 1963), it was until recently considered nonessential in humans, as it could be removed without obvious consequences (Lucas, 1991; Carroll et al., 1992). It is still stated in some medical textbooks that the human spleen is noncontractile (Pappas et al., 2013), but several studies have shown that it has an ability to contract and transiently boost the blood with stored erythrocytes in various situations involving hypoxia (Hurford et al., 1990; Schagatay et al., 2001; Stewart and McKenzie, 2002; Bakovic et al., 2003). Also, exercise induces the response (Laub et al., 1993).
This “blood boosting” ability has been found to be associated with an increased apneic duration when the spleen blood was recruited, leading to an enhanced ability to store and transport oxygen as well as carbon dioxide (Schagatay et al., 2001; Bakovic et al., 2003). While it was found that participants with intact spleens increased their apneic durations across a series of apneas, splenectomized participants did not show any increased durations (Schagatay et al., 2001). Human spleen contraction was first observed after a diving shift in Japanese Ama divers by Hurford et al. (1990), but the Ama divers were not found to have larger spleens than nondivers. Competitive breath-hold divers were later reported to have bigger spleens than nondivers, and among competition divers in a world championship, it was found that the most successful divers had the largest spleens (Schagatay et al., 2012). It was previously found that chronic obstructive pulmonary disease (COPD) patients with severe hypoxia (resting values <90% SpO2) had larger spleens than less hypoxic patients (Schagatay et al., 2015), and it was recently reported that the Bajau, an ethnic group of apnea divers in Indonesia, had larger spleens than a land-based population (Ilardo et al., 2018). Thus individual differences in spleen size could potentially affect the tolerance to hypoxic situations.
Both hypoxia and hypercapnia contribute to initiating spleen contraction (Lodin-Sundström and Schagatay, 2010), with hypoxia being the most important stimulus (Richardson et al., 2012). It is an active contractile response (Bakovic et al., 2003) and it is not to be considered a part of the cardiovascular diving response, as it is not initiated by the same stimuli (Schagatay et al., 2007).
The factors determining individual tolerance to altitude are quite unknown. The spleen could be involved as it stores a reserve of red blood cells ejected into circulation during hypoxia. Spleen volume was previously found to correlate with apnea diving ability, and it may also characterize good climbers. It was previously found that the spleen contracted more during apnea in climbers after they had summited Mt. Everest, compared to before the climb (Engan et al., 2014). In another study of trekkers to less extreme altitude, it was found that spleen volume as well as its contractile ability had increased (Rodriguez-Zamora et al., 2015). In this study, we aimed to investigate whether spleen size differs between experienced high-altitude climbers going to the summit of Mt. Everest and recreational trekkers going to Mt. Everest Base Camp (EBC). We also wanted to study the magnitude of their spleen contractile response during a short bout of hypoxia achieved by breath-holding, and its effect on hemoglobin concentration (Hb).
Methods
Participants
A total of 24 participants were recruited through personal communication by travel/mountaineering agencies from two existing categories of Mountaineers depending on what they already signed up for: 12 “Climbers” (3 women), that is, experienced high-altitude athletes with the ambition and preparation for summiting the world's highest mountain, and 12 “Trekkers” (3 women), that is, recreational mountaineers aiming to trek up to EBC at 5300 m. Among Climbers, all had been over 6000 m before, with a mean (standard deviation [SD]) maximal climbing elevation of 6390 (2360) m. Two had been on over 3000 m and one had slept in a hypoxic high-altitude tent during the last 2 months. Among Trekkers, none had ever been over 6000 m, but nine had been on 3000 m or higher previously, with a mean (SD) maximal peak elevation of 3041 (2023) m. None of the Trekkers had been over 3000 m during the last 2 months. The experience in breath-hold diving was the same among groups, with most participants being completely inexperienced, or had tried breath-hold diving long ago, and no one was a free diver in training. All participants reached the intended altitude after the tests, except one Climber who turned down at 7000 m and one Trekker who was evacuated by helicopter from 4400 m due to acute mountain sickness. All but four Climbers summited using supplementary O2 over 8000 m. The Climbers expedition lasted 7 weeks, of which 35 days were spent above EBC altitude (5300 m). The Trek to EBC took 14 days in total, 9 to reach EBC and an additional 5 days to return to Kathmandu. Information regarding climbing and trekking success was obtained through individual correspondence after the expedition.
After receiving detailed written and oral information about the procedures, participants signed an informed consent form. The test protocol had been approved by regional Human Ethics boards in Sweden and Nepal, and applied with the Helsinki declaration of 2004.
Procedures
All Climbers and Trekkers did an apnea test with spleen volume assessment in Kathmandu (1370 m) 1–3 days after arrival and just before the departure to start the climb/trek. Testing was done in a field laboratory in Kathmandu as this is where many Mt. Everest Climbers and Trekkers from different parts of the world gather before starting the trek. The participants arrived at the laboratory after at least 1 hour without eating, drinking, or performing physical exercise. After collection of anthropometrics (Table 1), they sat down and remained seated during the whole experiment. Information about their prior high-altitude experience was collected, after which vital capacity (VC) was measured in duplicate (Compact II; Vitalograph, Buckingham, England) in the sitting position and the largest value used. Baseline resting arterial oxygen saturation (SpO2) and heart rate (HR) were collected using pulse oximetry (Lifesense LS1-9R; Medair AB, Delsbo, Sweden). The apnea test consisted of two apneas done in the sitting position, the first apnea (A1) with a duration limited to 60 seconds (A1) and, after 2 minutes recovery, another one of maximal voluntary duration (A2; Fig. 1). In six of the Climbers (one woman), the two apneas were followed by a third, maximal apnea (A3) to trigger a maximal spleen contraction (Bakovic et al., 2003; Schagatay et al., 2005) and to detect its effects on Hb. The spleen volume during recovery after apneas was studied for 10 minutes in this group. The tight schedule for the testing of most of the participants did not allow inclusion of a third apnea and only allowed a 5-minute recovery period in the protocol, to keep the total time in the laboratory within 30 minutes.
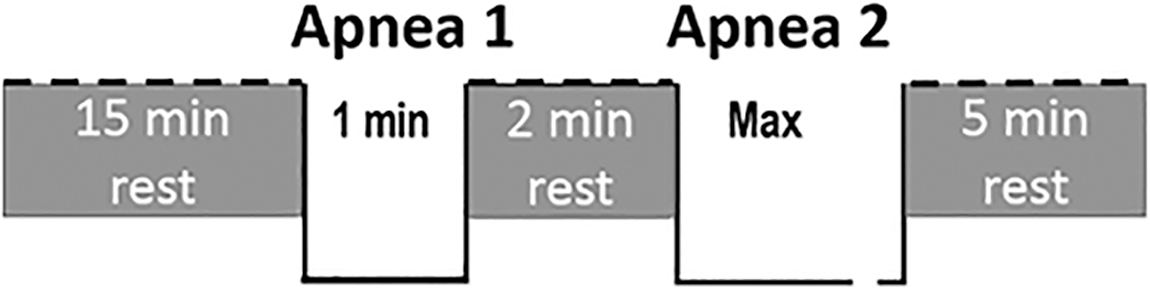
Apnea testing protocol used to induce spleen contraction in 18 subjects. In six climbers, there was a third apnea for maximal duration added after 2 minutes of rest, followed by 10 minutes of rest.
Mean (Standard Deviation) Age, Height, Weight, Vital Capacity, Resting HR, and SpO2 of Participant Groups
HR, heart rate; SpO2, arterial oxygen saturation.
The participants were instructed to avoid hyperventilation before apneas and to perform apneas after one maximal exhalation followed by a deep, but not maximal inspiration, and hold the breath for the intended time without exhaling. These instructions have been shown to lead to lung volumes of ∼85% of VC (Andersson and Schagatay, 1997). After an average of 13 (1) minutes of sitting rest, 2-minute countdown started, at 30 seconds before the start of apnea, a nose-clip was placed, and after a 10-second second-by-second countdown, participants performed a 1-minute apnea, notified of the end by a 5-second countdown. After a 2-minute rest, including 1-minute countdown, the maximal duration apnea was performed without time cues (Fig. 1).
Every minute before and after the apneas, three axial maximal spleen diameters were measured by ultrasonic imaging (M-Turbo Ultrasound system; FUJIFILM SonoSite Inc, Bothell, Washington) from the back in the sitting subject for volume calculation (Fig. 2). In the six Climbers performing three apneas Hb was assessed in duplicate finger capillary blood samples collected in microtainer cuvettes and analyzed in a portable hemoglobin analyzer (Hemocue AB, Ängelholm, Sweden) before and after the three apneas, and after 10 minutes of recovery.
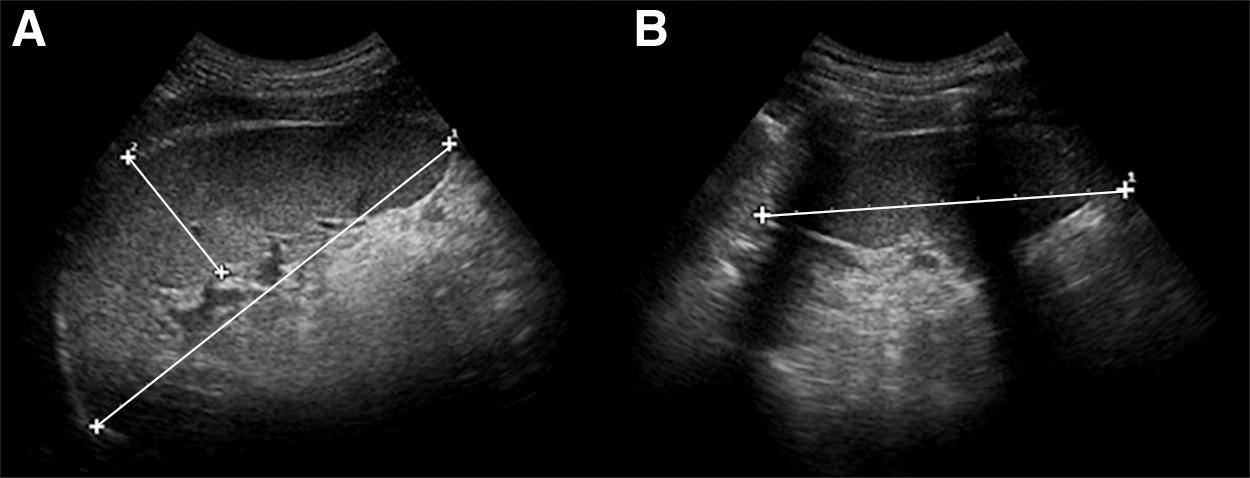
Ultrasonic image of the spleen, with measurements of maximal length and thickness
Analysis
The three axial spleen diameters were used to calculate spleen volume by the Pilström equation L (WT − T2)/3, where spleen length (L), thickness (T), and width (W) were obtained from the ultrasound images (Schagatay et al., 2005). For all the 24 participants, nadir spleen volumes after A1 and A2, and the value after 5 minutes of recovery were compared to the baseline volume before apneas. For the six Climbers performing three apneas, values after each apnea and after 10 minutes of rest were compared with the spleen volume at 2 minutes before A1, and the Hb values collected before and after the three apneas at the same time points as spleen values were compared.
Statistical analysis
The within-group differences in spleen volumes and Hb values before and after the apneas were evaluated using a one-way repeated measures analysis of variance, with Bonferroni corrections for repeated measures. To compare the mean difference between Trekkers and Climbers on baseline spleen volume, spleen volume contraction, and postapnea recovery, a mixed model analysis of covariance was conducted, with Bonferroni corrections for repeated measures. Where significant differences were found, further independent samples t-tests were carried out to measure the difference. Cohen's d was used to estimate effect size (ES) and calculated for each result along with 95% confidence intervals (CI). Interpretation of ES was based on three categories: 0.0–0.3 was considered a small effect; 0.4–0.7 a medium effect; and ≥0.8 a large effect (Lee, 2016). Significance was accepted at p < 0.05 and nonsignificant trends were denoted for p < 0.1.
Results
Mean (SD) baseline spleen volume during rest was larger in Climbers, at 367 (182) mL than in Trekkers, 228 (70) mL (p = 0.022, ES = 1.01; 95% CI 0.13–1.82; Fig. 3).
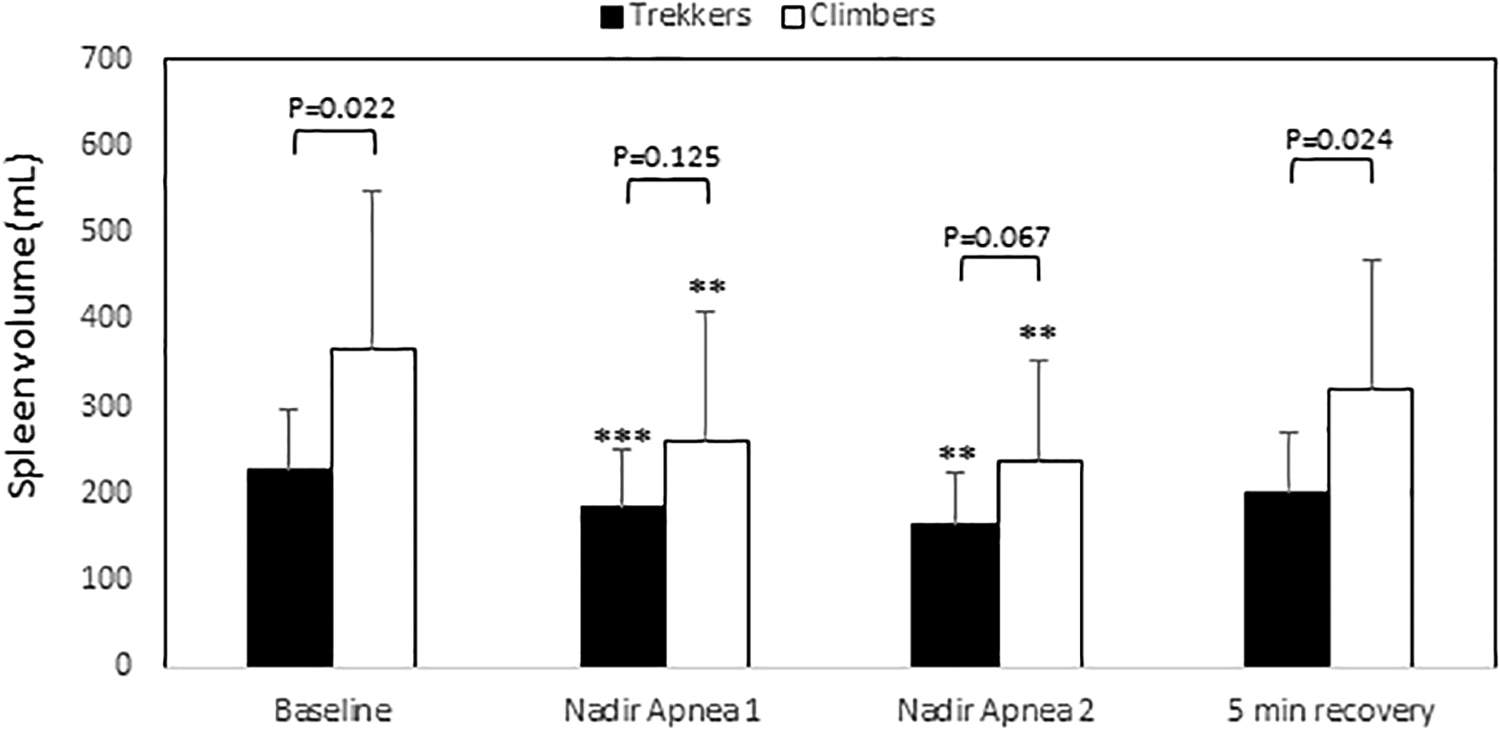
Mean (SD) spleen volumes in Trekkers (n = 12) and Climbers (n = 12). p-Values for differences between groups are shown above columns. ***indicates p < 0.01 from baseline, and **p < 0.05 from baseline. SD, standard deviation.
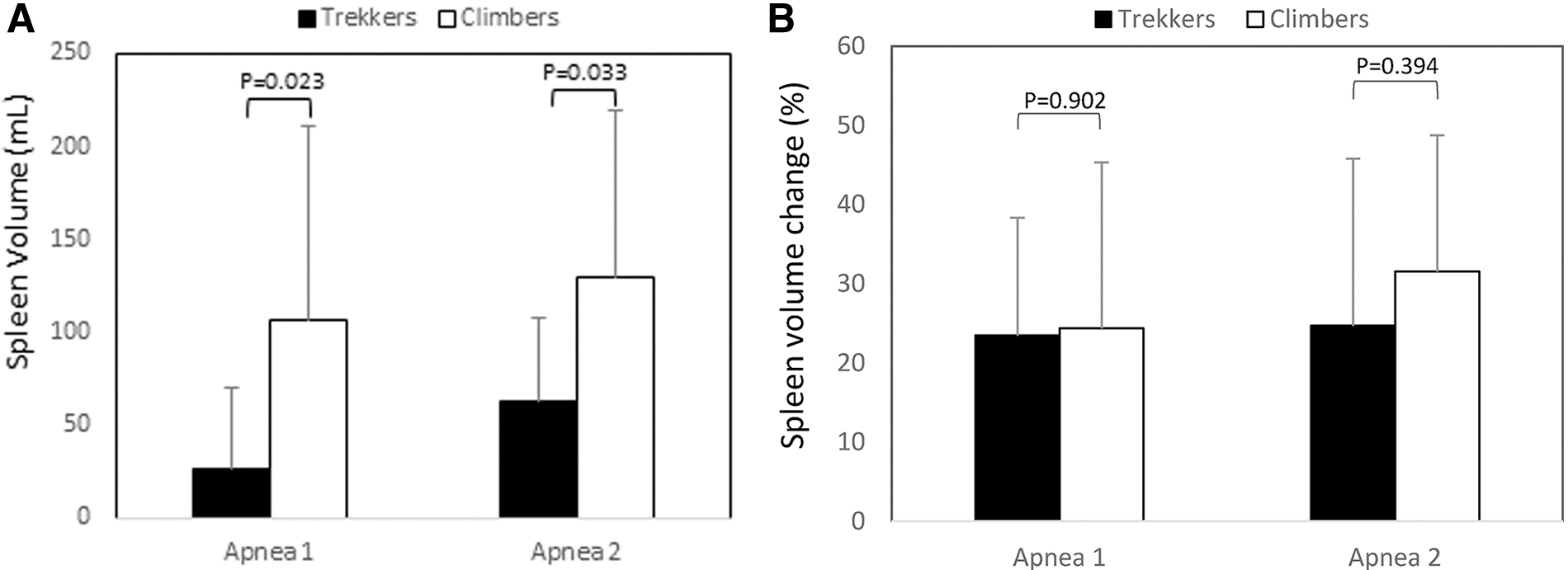
Spleen contraction occurred during apneas in both Climbers and Trekkers after A1 and A2 (all p < 0.01 from baseline; Fig. 3), with about twice the magnitude in Climbers for both A1 (p = 0.023, ES = 1; 95% CI 0.12–1.81) and A2 (p = 0.033, ES = 0.94; 95% CI 0.06–1.74; Fig. 4).
After 5 minutes of recovery, spleen volume had increased to preapnea values in Climbers (p = 0.095 compared to baseline), and in Trekkers, it had increased, but not reached preapnea values (Fig. 3; p = 0.04 from baseline).
Mean (SD) apneic durations were similar in A1 for Trekkers, 54 (8) seconds, and Climbers, 58 (9) seconds (p = 0.350), with five participants in each group not reaching 60 seconds. The maximal apneic duration in A2 was longer in Climbers at 88 (23) seconds versus 67 (18) seconds in Trekkers (p = 0.023, ES = 1.02; 95% CI 0.13–1.83). Of the trekkers who could not reach 60 seconds in A1, only one increased the apnea duration over 60 seconds in A2. For the Climbers who could not hold their breaths for 60 seconds, all but one was able to increase the duration more than 60 seconds for A2.
Six of the Climbers did three apneas with a mean (SD) duration of 58 (13), 83 (21), and 92 (23) seconds, respectively, which induced a spleen response similar to that of the other Climbers. Their mean (SD) spleen volume was reduced from 348 (145) mL at 2 minutes before apneas to 270 (124) mL directly after A1 (p = 0.016; ES = 0.6; 95% CI −0.6 to 1.7), 251 (101) mL after A2 (p = 0.036; ES = 0.8; 95% CI −0.5 to 1.9), and 219 (103) mL after A3 (p = 0.011; ES = 1.0; 95% CI −0.2 to 2.1; Fig. 5). The lowest volume was observed 1 minute after A3 at 202 (91) mL (p = 0.005; ES = 1.2; 95% CI −0.1 to 2.39 compared to preapnea) where the spleen volume was also smaller than after A2 (p = 0.012). After 10 minutes of rest, spleen volume had increased to 309 (160) mL, and approached preapnea volume (not significant). Hb increased across three apneas from a preapnea mean (SD) of 147.9 (13.1) to 153.3 (11.3) g/L (p = 0.024, ES = 0.5; 95% CI −1.6 to 0.7) post apnea 3. After 10 minutes of recovery, it was still elevated at 153.2 (11.1; p = 0.045; ES = 0.5; 95% CI −1.6 to 0.7) compared to preapneas (Fig. 6).

Minute by minute mean (SD) spleen volumes in six climbers across three apneas spaced by 2 minutes of breathing, and after 10 minutes of rest. Arrow indicates blood sampling, and the time points used for statistical analysis between 2 minutes preapnea and first postapnea spleen values. p-Value compared to preapnea is shown above postapnea value. After 10 minutes of rest, spleen volume was similar to the preapnea value (not significant).
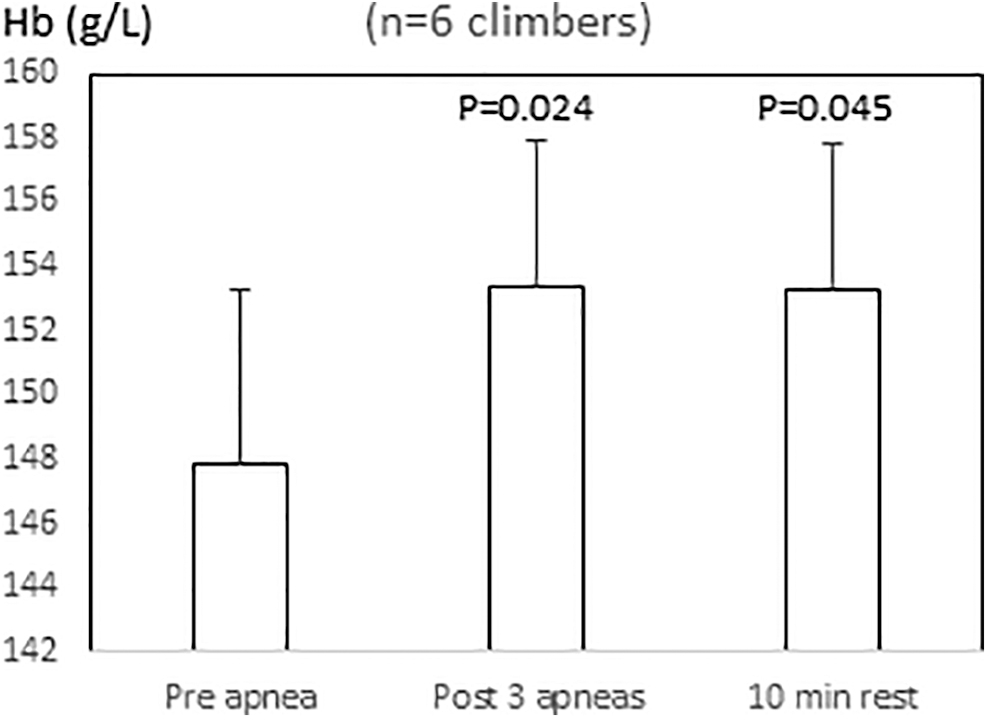
Hemoglobin concentration (Hb) in six climbers during rest at 2 minutes before apneas, directly after three apneas, and after 10 minutes of rest. p-Value compared to preapnea is shown above postapnea values.
Discussion
This study shows that baseline spleen volume during rest is larger in Climbers than in Trekkers, which is a novel finding. The mean spleen volume observed in Climbers, 367 mL, was about 60% larger than in Trekkers, but similar to the average spleen volume of 14 male competitive apnea divers, 336 mL (Schagatay et al., 2012), while the Trekkers' spleen volume corresponded with the 215 mL observed in 140 lowlanders of both genders (Prassopoulos et al., 1997). They did not report any difference between genders or age groups (Prassopoulos et al., 1997). Another group in which large spleens have been reported was 9 patients with COPD, with resting SpO2 below 90% mean spleen volume of 340 mL, versus 203 mL in 15 COPD patients with baseline SpO2 above 90% (Schagatay et al., 2015), suggesting that chronic exposure to hypoxia may increase spleen volume. Such an effect could possibly be involved in enlarging the spleens of elite Climbers. While studies on elite apnea divers (Schagatay et al., 2012) and COPD patients (Schagatay et al., 2015) suggest that large spleens could be the result of frequent hypoxic exposure in these groups, it was recently reported that spleen volume is greater in the Bajau, native apnea divers in Indonesia, than in a land-based ethnic group, which was suggested by the authors to be a result of identified genetic traits (Ilardo et al., 2018).
The effects of the spleen contraction on Hb were evident in the subgroup of Climbers doing three apneas, in accordance with the blood boosting effect reported in several previous studies (Schagatay et al., 2001, 2005; Bakovic et al., 2003). Although the human spleen is a lymphoid organ with known immunological function, we do not think this is likely the functional link to the difference in spleen volume between Climbers and Trekkers, as there is, to our knowledge, no evidence of association between spleen volume increase and infection. It is more likely that the difference between groups in spleen volume represents an increased complementary blood reservoir function in the Climbers.
The larger spleen volume in Climbers than in Trekkers, together with the enhanced spleen contraction found in the Climbers, suggests that spleen function as a dynamic red cell reservoir could be important for climbing to extreme altitude, just as it was previously shown to be related to diving performance in apnea diving competition (Schagatay et al., 2012). It further implies that either a large spleen volume is a prerequisite for climbing high, that is, Climbers are genetically predisposed enabling them to be successful at climbing high, or the spleen volume is in fact increased by repeated climbing to high altitude. Summiting 8000 m peaks requires considerable climbing experience and physiological preparation, including physical training and ascent of other high peaks, which could be responsible for a training effect. Spleen size is highly variable, spanning from 107 to 342 mL in normal healthy individuals (Prassopoulos et al., 1997). Most likely a favorable genetic predisposition is necessary to climb high, and our results suggest that this could include genes promoting spleen growth. We speculate based on previous research that the differences in spleen size and contraction between groups in this study could be due, to some extent, to both genetic predisposition and repeated exposure to hypoxic environments.
There are some earlier studies that suggest that spleen growth could be involved. The mammalian spleen appears to have a high regenerative ability. In baboons, after experimental removal of 2/3 of the spleen, the remaining spleen tissue showed a compensatory growth (Van Rensburg et al., 1991) and in humans, after splenectomy or removal of the main spleen, growth of a small, accessory spleen has been demonstrated (Beahrs and Stephens, 1980). Prevalence of accessory spleens in cetaceans, who much like high-altitude climbers are regularly exposed to hypoxia, is more than 60%, and it increases with the typical diving depth of the species (e Silva et al., 2014). Accessory spleens with contractile abilities have also been observed in breath-hold divers (Richardson et al., 2009). After hepatectomy, the human spleen increased in volume by 35% of its original size within 6 months in parallel with liver regeneration, suggesting that the same growth factors are involved in spleen and liver growth (Ibrahim et al., 2005).
We speculate that spleen growth at altitude could probably be functionally related to the polycythemia caused by the hypoxic exposure, where it would have an obvious function in storing a larger amount of red cells between spells of exercise to reduce blood viscosity. The enhanced spleen contraction during apnea could be a result of frequent contraction occurring with desaturation during the exercise of climbing at high altitude. Hypoxia and exercise are factors that could potentially affect spleen volume in the long term, as both have been shown to contribute to its contraction (Laub et al., 1993; Richardson et al., 2008).
Several studies have demonstrated that spleen function is important for maximal apnea diving performance (Schagatay et al., 2001, 2012; Bakovic et al., 2003), and a recent long-term study involving hypoxic exposure by apnea training showed spleen enlargement after 8 weeks, but no increase in spleen contraction (Bouten et al., 2019). The role of the spleen at high altitude has not yet been fully clarified, but in a longitudinal study of Mt. Everest climbers, it was found that spleen contraction was greater after summiting than before the 6-week climb, while the baseline spleen volume remained the same (Engan et al., 2014). In a study of the same duration involving trekkers to less extreme altitude, it was observed that both spleen volume and contraction had increased after a 6-week exposure (Rodríguez-Zamora et al., 2015). The difference between these two studies could possibly be ascribed to a general catabolism in the Death Zone, not allowing spleen enlargement. Although the Climbers of this study were not fully acclimatized to high altitude at the time for testing before the climb, their pre-expedition preparations, including high-altitude exposure, may have enhanced their spleen volume and function, while Trekkers preparation did not include high altitude.
Our findings of enhanced spleen volume with high-altitude exposure are contradictory to those of Sonmez et al. (2007). They conducted a study on healthy military recruits at different time points after they had started living at high altitude, and concluded that spleen volume gradually decreased with time at altitude (Sonmez et al., 2007). The authors did not rule out that lifestyle changes associated with moving to high altitude could also have affected the results. Their participants performed apnea during baseline measurements and it makes it difficult to interpret their results as they provide no information on the durations of apneas that were performed, as there is clear evidence that apnea induces spleen contraction from previous (Schagatay et al., 2005) and this study. Responses to apnea could change by residing at high altitude. The findings that spleen contraction during apnea was enhanced after 6 weeks at high altitude (Engan et al., 2014; Rodríguez-Zamora et al., 2015) may suggest that such changes were present after residing for an extended period at high altitude in the study of Sonmez et al. (2007).
Previous studies using simulated altitude reported a resting spleen volume reduction of 16% during 20 minutes of normobaric eupnea of 14.2% O2, equivalent to 3000 m (Lodin-Sundström and Schagatay, 2010), and a spleen volume reduction of 18% when participants were exposed to12.8% O2, equivalent to 4000 m, with concomitant increases in Hb (Richardson et al., 2008). These results contrast with those of a recent investigation of spleen volume and reactivity at high altitude, which reported unchanged resting spleen volume with incremental ascent at altitude (Purdy et al., 2018). While injection of phenylephrine hydrochloride elicited spleen contraction at all altitudes investigated (1045, 3440, and 4240 m), hand grip exercise failed to induce spleen contraction at higher altitudes. The authors concluded that the spleen does not contribute to acclimatization to chronic hypoxia (Purdy et al., 2018).
Our data suggest that the enhanced spleen contraction occurring during apnea in Climbers compared to Trekkers is likely, to a large extent, a consequence of their larger spleen volumes, as previously observed in COPD patients where the most hypoxic patients had larger spleens and contraction leading to higher elevation of Hb during exercise (Schagatay et al., 2015). Multiple factors may contribute to spleen contraction, including hypoxia, hypercapnia (Lodin-Sundström and Schagatay, 2010; Richardson et al., 2008, 2012), exercise, and general sympathetic activation (Stewart and McKenzie, 2002), but in the studied situation, the apnea in itself and hypoxia are likely the major stimuli. When apneas are performed at 1370 m altitude, the drop in SpO2 occurs rather fast, and both groups started with the same SpO2. For the 1-minute apnea, the desaturation would be similar for both groups, but with the maximal apneic duration longer in Climbers, it could possibly induce a somewhat stronger spleen contraction. However, a more pronounced spleen contraction in Climbers was seen also in the first apnea, suggesting the capacity to contract at a given stimulus may be greater in Climbers. Of the Climbers who could not hold their breaths for 60 seconds, all but one were able to increase the duration more than 60 seconds for A2, while in Trekkers, one did so. An interpretation could be that the larger spleen contraction in Climbers in A1 induced an Hb elevation that helped them to breath-hold longer in A2.
The responses in six Climbers after three apneas suggest that maximal spleen contraction was probably not induced after two apneas. The magnitude of spleen volume contraction has previously been shown to be correlated to the magnitude of the transient Hb increase after apnea (Richardson et al., 2008). In the subgroup of climbers where Hb was recorded, the spleen volume reduction after three apneas to only 202 mL was associated with an increase in Hb from 147.9 to 153.3 g/L. However, while spleen volume had nearly returned to baseline after 10 minutes, Hb was still elevated after this period. The remaining elevation of Hb could suggest a time delay before all red blood cells have been restored in the spleen after its re-expansion.
Large lung volume has previously been identified in high-altitude populations (Wu and Kayser, 2006), a characteristic that is also typical of apnea divers (Schagatay et al., 2012). In our study, VC was only slightly larger in elite Climbers than in Trekkers, and some other researchers have reported lung volume in climbers to be similar to normal values (Schoene, 1982; Oelz et al., 1986). Bernardi et al. (2006) found the same VC in climbers successfully reaching >8500 m without oxygen compared to those using O2. Aside of large lungs and a pronounced spleen contraction, a powerful diving response has been found to be important for apnea performance (Schagatay and Andersson, 1998). It appears that the same changes could be of general benefit in apnea diving and high-altitude climbing, and it could be interesting to reveal if the same genetic predisposition is favorable in these different natural situations involving hypoxia.
Sensitivity to asphyxia may also differ between Climbers and Trekkers. Acclimatized extreme climbers show lower hypoxic ventilatory response during rest compared to less extreme climbers (Bernardi et al., 2006), and during exercise compared to nonclimbers (Puthon et al., 2016). The hypercapnic ventilatory response during rest was also lower in climbers compared to matched control at similar environmental conditions (Puthon et al., 2016). The more powerful contraction in Climbers in our study is thus not likely a result of stronger chemoreceptor drive.
Other physiological characteristics of the Climbers, with a general benefit in hypoxic situations, could have contributed to a prolongation of the apneas, in line with the idea that the same characteristics are needed for climbing and apnea diving.
Recovery was studied only during 5 minutes in most participants, due to time limitations for testing. This means we could not likely see their full recovery of spleen volume, as this takes 8–9 minutes (Schagatay et al., 2005). This was supported by the spleen volumes studied in six Climbers, which had returned to baseline after 10 minutes.
While our study shows that large spleens characterize good climbers, further studies are needed to reveal the physiological effects and functions at high altitude of their larger spleens and volumes of contraction compared to Trekkers, and whether their causes are mainly due to genetic predisposition or induced by previous exposure or training.
Conclusions
We concluded that spleen volume during rest, as well as the volume of spleen contraction during apnea, are enhanced in Climbers compared to Trekkers. This leads to elevation of Hb across apnea and likely other hypoxic challenges, which suggest a significant role of the spleen as an active red cell reservoir at high altitude. To reveal whether these differences between groups are due to genetic predisposition or previous exposure to hypoxia awaits further study.
Footnotes
Acknowledgments
We thank the participants for their kind contributions, despite their tight schedules. We also thank the many people helping us with various matters during our work in Nepal: Mr. Chhiring Dorje Sherpa and collegues at Rolwaling Excursions, Mr. Ram Sapkota at Mountain Delights Treks and Expeditions, Mr. Nicke Sundström, Pathfinder Travel, Mr. Pasang Karma Sherpa, Ms. Annelie Pompe, Dr. Irina Zelenkova, Dr. Arjun Karki, Patan Academy Hospital, and Mr. Pradeep Pakhrin, Manager at Fuji Hotel where our laboratory was placed.
Authors' Contributions
E.S.: Ideas and development of the research questions, planning and organizing laboratory, data collection, data analysis, article writing, and proof reading. P.H.: data collection, planning and organization of laboratory, data analysis, article writing, and proof reading. E.M.: planning and organization of laboratory, data collection, and proof reading. P.L.: data collection and proof reading. F.S.S.: planning and organizing laboratory, data collection, and proof reading. H.E.: data collection and proof reading. A.L.-S.: planning and organization of laboratory, data collection, data analysis, article writing, and proof reading. All authors have read and approved of this article.
Author Disclosure Statement
No competing financial or other interest exists.
Funding Information
This study was supported by a grant from the Swedish National Centre for Research in Sports (CIF) and Mid Sweden University Research Funding.