
Abstract
High altitudes are physiologically challenging: the hypobaric hypoxia, cold, and increased ultraviolet radiation mean humans ascending to high altitude faster than they acclimatize risk life-threatening illnesses. Despite such challenges, birds can thrive at high altitudes and some even complete metabolically costly migrations across the world's highest mountain ranges. We outline the aspects of avian anatomy and physiology that confer advantages at each level of the oxygen transport cascade and compare them with those of human and nonhuman mammals. We also discuss additional adaptations that have been described for high-altitude specialist species of birds and how these are mirrored in high-altitude adapted mammals.
Introduction
High-altitude environments present a range of challenges to life, including cold, dehydration, increased exposure to solar radiation, malnutrition, and hypobaric hypoxia (Bouverot, 1985). Above 2000 m altitude, humans may begin to suffer from high-altitude illnesses (reviewed in Bärtsch and Swenson, 2013). Despite this, elite human mountaineers may spend years (and tens of thousands of dollars) preparing for ascents of the world's highest peaks (Fig. 1).

Schematic diagram showing records of humans and birds found at high altitude in relationship to commercial aircraft cruising altitude and some of the world's highest mountains. High-altitude birds are, for the majority of the time, found at much lower altitudes, but make rare flights above 5500 m altitude. Sources: Noel (1927), Laybourne (1974), West (2002), Mukherjee et al. (2008), McCracken et al. (2009a), Borras et al. (2010), Hawkes et al. (2011), Beall (2014), Dawson et al. (2016), Parr et al. (2017), and Everest by Numbers (2018).
To survive in high-altitude environments, animals require a suite of physiological adjustments along the oxygen transport cascade to maintain an adequate supply of oxygen to respiring organs and tissues (Fig. 2). These adjustments may take any, or all, of three forms. First, those that are fixed as the result of generations of adaptation (the process of selection pressures increasing the expression of genes that alter physical or behavioral features at a species or population level) (Willmer et al., 2005). For example, some human populations have adapted to life at high altitude, and these adaptations vary with respect to geographical origin (e.g., Tibetan vs. Andean people, see reviews elsewhere; Beall, 2007; Azad et al., 2017). Second, short-term phenotypically plastic responses (referred to as “acute hypoxia responses” from now on), which are almost instantaneous responses to hypoxia exposure, but are usually temporary and reversible (Piersma and van Gils, 2011). Third, longer-term plastic responses following continued exposure, also known as “acclimatization,” which can last for days to years. Acclimatization enables humans native to low altitudes to transiently visit extremely high altitudes, for example, to climb the world's 14 mountains over 8000 m without the use of supplementary oxygen (Wolfel et al., 1991).
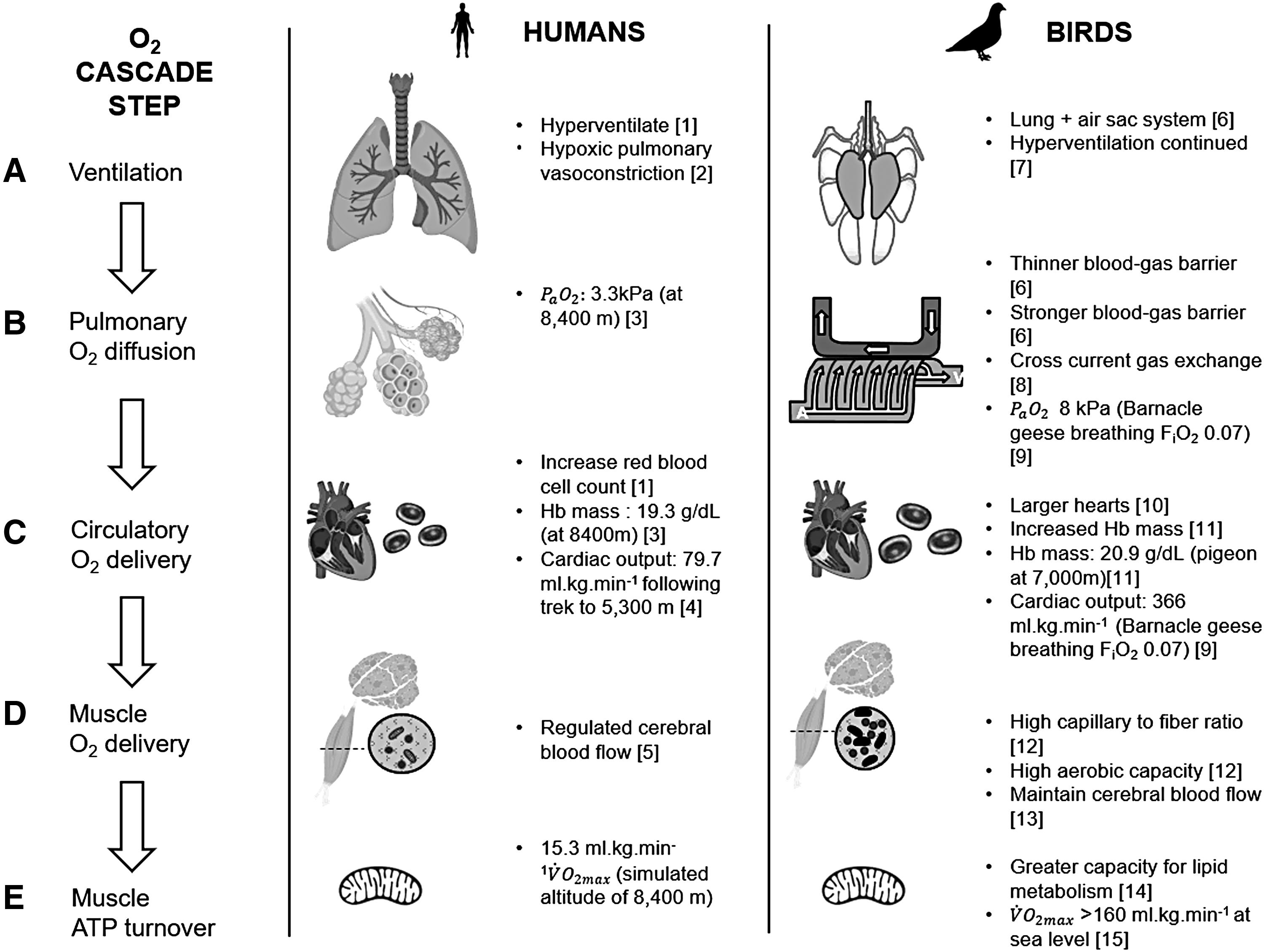
Diagram of each stage
Birds, regardless of the altitude at which they live, display essential features of their anatomy and physiology that put them at an advantage compared to mammals in hypoxic environments at high altitude. This was demonstrated by Tucker (1968) using a very simple hypoxic chamber experiment. After rapid ascent to a simulated altitude of 6100 m, a mouse was rendered comatose, while a house sparrow was able to maintain flight in a wind tunnel. The features of avian physiology that enabled this high tolerance for hypoxia are not necessarily the direct result of evolutionary adaptations following chronic hypoxic selection pressures, rather they likely evolved due to the high oxygen demands required to sustain flight. Flight is a very costly form of locomotion in terms of oxygen demand, requiring some 10–12 times resting metabolic rate (Bishop et al., 2002). Powered flight also evolved in bats, which (like birds) are also capable of high rates of oxygen consumption ( ) (reviewed in Maina, 2000). High may therefore serve as a preadaptation for tolerating hypoxia in birds, and to some degree bats (Thomas et al., 1995), which weather radars have estimated to transiently visit altitudes up to 3000 m (Williams et al., 1973; Peurach, 2001).
) (reviewed in Maina, 2000). High may therefore serve as a preadaptation for tolerating hypoxia in birds, and to some degree bats (Thomas et al., 1995), which weather radars have estimated to transiently visit altitudes up to 3000 m (Williams et al., 1973; Peurach, 2001).
In contrast to the majority of avian species, some species of birds spend a portion of their lives at high altitude (either migrating or residing there, Fig. 1) and can thus tolerate hypoxia better, due to additional adaptations or greater ability to make further temporary physiological adjustments (such species will be referred to as “high-altitude adapted” birds from now on). The best studied high-altitude adapted avian species is the bar-headed goose (Anser indicus), which completes a trans-Himalayan migration, including a climb of 4000 to 5000 m from sea level in just 7–8 hours between wintering grounds in south Asia to breeding grounds in central China and Mongolia (Hawkes et al., 2011, 2012). On exposure to progressive hypoxia in laboratory studies (so removing the other stressors present at high altitude and the added costs of flight), bar-headed geese were able to remain standing and alert at a simulated altitude of 12,190 m, whereas lowland Canada geese were unable to stand at altitudes higher than 10,668 m (Black and Tenney, 1980a), although both altitudes were considerably higher than low-altitude mammals could tolerate (Tucker, 1968).
In addition to bar-headed geese, other avian migrants fly at very high altitudes (>5000 m), potentially in search of favorable conditions (Senner et al., 2018). However, to date, the highest altitudes directly recorded using tracking tags are 7290 m for a bar-headed goose, and 6800 m for a ruddy shelduck, although both species remain below 5500 m for the overwhelming majority (95%) of their migration (Hawkes et al., 2011; Parr et al., 2017). In addition to visitors to high altitude, several avian species that are resident at high altitude have become the subject of recent studies of high-altitude physiology and comparative transcriptomics. These species include passerines (perching birds) of the Himalayas (Grewal, 2017; Hao et al., 2019) and multiple species of waterfowl in the Andes, such as torrent duck (Merganetta armata, found up to 4500 m in riverain habitats of the Andes) (Dawson et al., 2016) and Andean geese (Neochen melanoptera, resident at 6000 m) (Fig. 1) (Maina et al., 2017).
Several mammalian species also reside at high altitude, including yak (Bos grunniens), llama (Lama glama), and vicuna (Vicugna vicugna). The high-altitude physiology of deer mice (Peromyscus maniculatus) has also been increasingly studied, given the large altitudinal ranges and possibility for translocation experiments between low- and high-altitude populations (reviewed in Storz et al., 2019). This review contrasts the mammalian and, in particular, human responses to high-altitude hypoxia with those of birds, at each level of the oxygen transport cascade, highlighting where the high-altitude environment has resulted in similar and divergent phenotypic adaptations.
Ventilation
In humans, the partial pressure of arterial oxygen (Pa
Birds too display an HVR, increasing breathing rates as Pa
In high-altitude adapted avian species, the HVR is more powerful than in lowland birds (Scott and Milsom, 2007). Bar-headed geese increase total ventilation on exposure to hypoxia by increasing tidal volume, in addition to respiratory frequency, a strategy that differs from low-altitude waterfowl that increase only ventilation rate (Scott and Milsom, 2007). Increasing tidal volume reduces the proportional contribution of anatomical dead space ventilation, and as a result, bar-headed geese sustain greater total ventilation for a given fraction of inspired oxygen than low-altitude geese, suggesting they have evolved a greater capacity for acute responses to hypoxia (Lague et al., 2016).
In addition, exposure to hypoxia during development appears to modify this response further: bar-headed geese raised at high altitude (3200 m) can sustain a greater HVR than bar-headed geese raised at sea level (Lague et al., 2016). Both groups of bar-headed geese in this case may respond to declines in Pa
Pulmonary Oxygen Diffusion
At the pulmonary interface, oxygen must diffuse into the blood at a rate determined by the pulmonary oxygen diffusion capacity. This is increased in Tibetan and Andean human populations through a larger pulmonary volume relative to lowlanders (Brutsaert, 2007). Larger lungs have also been found in mice (Phyllotis darwini) living at high altitudes (4660 m) relative to sea level populations (Pearson and Pearson, 1976). High-altitude adapted birds also have larger lungs relative to low-altitude species (Scott et al., 2011), but (regardless of altitude exposure) birds have on average 15% larger lung surface areas than mammals of equivalent body mass (reviewed in West, 2009). The blood-gas barrier is also considerably thinner in birds than mammals (Fig. 2B). Nonvolant mammalian blood-gas barriers can range from 0.33 μm (Etruscan shrew, Suncus etruscus) (Gehr et al., 1980) to 0.72 μm (domestic pig, Sus scrofa domesticus) (Meban, 1980), although heightened oxygen demand in bats has selected for a thinner blood-gas barrier (range 0.20 μm in the hairless bat, Cheiromeles torquatus, to 0.28 μm in the lesser short-nosed fruit bat, Cynopterus brachyotis) relative to nonvolant mammals (Maina and King, 1984). In contrast, blood gas barriers of 0.1 μm are not unusual in birds (such as waterfowl, sea birds, and passerines, but thickness can range from <0.1 μm (hummingbirds) to 0.56 μm in nonvolant birds such as ostriches) (Dubach, 1981; Maina and West, 2005; reviewed in West et al., 2007). Perhaps, surprisingly, given the thinness of the avian blood-gas barrier, the avian lung surface is much stronger compared with mammals. It is less susceptible to stress failures and more resistant to pressure than in the mammalian lung (Watson et al., 2007, 2008).
In the avian lung, gas exchange is also far more effective than in mammals, not only as the result of unidirectional air flow but also because of a cross-current gas exchange system (Fig. 2B) that theoretically allows PaO2 to exceed the partial pressure of oxygen (PO2) of expired air (Scheid and Piiper, 1972). Air capillaries are intertwined with blood capillaries that deliver oxygenated blood to blood vessels running perpendicular to rigid parabronchioles, and therefore, the diffusion difference of oxygen is not diminished across the length of the avian parabronchioles (reviewed in Scott, 2011). Indeed, the cross-current arrangement of the air and blood capillaries in birds may be so significant that Powell and Scheid (1989) estimated that a human climber on Mount Everest could climb 780 m higher if they had an avian parabronchial lung arrangement (reviewed in Maina, 2015). Although it has been little studied in wild flying birds (given the technical challenges), the avian lung is likely overall a far more effective gas exchange organ than the mammalian lung (but see Thomas et al., 1984).
In addition to the general avian advantage over mammals, high-altitude adapted birds may be able to increase pulmonary oxygen extraction over that of low-altitude species (McCracken et al., 2009a; Natarajan et al., 2015; York et al., 2017). For example, work in waterfowl (Andean geese and crested ducks) has suggested that extraction efficiency can increase with altitude up to 90% (recorded at a simulated altitude of 11,000 m in Andean geese), but is ∼45% at 12.2 kPa partial pressures of inspired oxygen (∼4500 m). In contrast, bar-headed geese maintain a consistent extraction efficiency of ∼40% across all altitudes (Laguë et al., 2017). The increased capillarity of the blood-gas interface, and a proportionally larger lung surface area of Andean geese relative to bar-headed geese (despite the relatively larger lung of bar-headed geese in comparison to lowland waterfowl) (Scott et al., 2011), has been suggested as a possible mechanism for this difference in extraction capabilities (Laguë et al., 2017; Maina et al., 2017; York et al., 2017). Differences in Andean geese and bar-headed geese lung morphology and extraction efficiency in acute hypoxia highlight the variation in responses to high-altitude selection pressures present in waterfowl. This is potentially due to the differing nature of chronic or transient altitude exposure, or geographic origins (for review see Laguë, 2017), as when the same selection pressures are present across species in the same geographic area, a recent transcriptomic study found shifts in gene expression were similar across three high-altitude species relative to closely related low-altitude species (Hao et al., 2019).
High-altitude pulmonary edema
In humans at low altitudes, when regions of the lung become hypoxic (due to uneven or poor ventilation), pulmonary vascular resistance increases through hypoxic pulmonary vasoconstriction, an adaptive vasomotor response that redistributes blood to better-ventilated lung segments (reviewed in Moudgil et al., 2005). However, this response can be maladaptive at altitudes where hypoxia is ubiquitous. As a result of increased or uneven blood flow to localized regions of the alveolar capillary bed (Maggiorini et al., 2001), the gas exchange barrier can become damaged, allowing proteins and erythrocytes to leak into the alveoli, restricting the potential for gas exchange, thus amplifying the vasoconstriction and stimulating an inflammatory response (reviewed in Swenson et al., 2002). This has been well described in human mountaineers (reviewed in Basnyat and Murdoch, 2003) as a cause of noncardiogenic pulmonary edema, known as high-altitude pulmonary edema (HAPE) (Bartsch et al., 2005, reviewed in Dehnert et al., 2007). In prolonged exposure, pulmonary arteries may thicken in response to chronic vasoconstriction and the resulting increase in pressure may cause right ventricular hypertrophy and failure (Tucker et al., 1975). Similarly in cattle at high altitude (above 2100 m), hypoxic pulmonary hypertension has been linked to right heart failure (brisket disease) and is a major cause of livestock death in high altitudes (Rhodes, 2005). In contrast, yaks (a closely related bovine) appear to be well adapted to the high-altitude environments of Tibet (found up to 4500 m) (Dolt et al., 2007; Qiu et al., 2012). Generations of evolution appear to have eliminated hypoxic vasoconstriction in this species (Anand et al., 1986) and yaks have thin-walled pulmonary vessels, despite chronic altitude exposure (Durmowicz, 1993). In addition, to the authors' knowledge, large herbivores such as llamas, which have been domesticated in high altitudes of South America 6000–7000 years ago, do not develop right ventricular hypertrophy or pulmonary hypertension (Heath et al., 1974; Riek et al., 2019).
In contrast to the majority of mammals, birds are not known to suffer from HAPE, because, similar to the high-altitude adapted yak, avian pulmonary vessels do not vasoconstrict in response to hypoxia, regardless of previous exposure of high altitudes (reviewed in West et al., 2007). Instead, pulmonary pressure only increases in proportion to cardiac output (Burton et al., 1968; Black and Tenney, 1980b; Faraci et al., 1984a), and thus, pulmonary edema does not appear to occur in birds when exposed to high altitude. Even if it did, the unidirectional ventilation and a structural honeycomb network of support struts of avian airways would ensure they could not become closed or blocked (Watson et al., 2007). However, in commercial poultry production in high-altitude environments, maladaptation from selective breeding of fast-growing chicks may have led to pulmonary arterial hypertension or “ascites syndrome” and cardiac failure (Yersin et al., 1992; Rostami et al., 2016). This is characterized by excessive accumulation of fluid in the abdominal cavity due to increased demands for cardiac output in hypoxic conditions, which is exacerbated by breeding programs for increased body size that is disproportional to heart and lung mass (Khajali and Wideman, 2016).
Circulatory Oxygen Delivery
Cardiac output
In addition to changes in ventilation, humans also respond to hypoxia by increasing cardiac output (Fig. 2C). Within 40 hours of exposure to a simulated altitude of 4600 m, heart rate increased by 34% in a study of resting lowlanders (Vogel and Harris, 1967). By comparison, high-altitude adapted humans from the Tibetan Plateau have a greater ability to increase heart rate, achieving higher maximal heart rates during exercise (for review, see Gilbert-Kawai et al., 2014). Similarly, high-altitude deer mice were found to sustain higher heart rates than low-altitude populations, both in high- and low-altitude conditions (reviewed in Ivy and Scott, 2017). Birds, in general, can sustain greater cardiac outputs; a 1 kg bird at rest has a cardiac output of ∼290 mL/min compared to 166 mL/min for a mammal of the same mass; in addition, birds have proportionally larger hearts than mammals (Calder, 1968; Grubb, 1983) and can more than double their cardiac output in hypoxia through a combination of increased heart rate and stroke volume (Black and Tenney, 1980a; Lague et al., 2016).
On acclimatization to hypoxia, much like humans, birds can also increase cardiac output, even in species already adapted to high altitude; bar-headed geese that had undergone a 4-week period at a simulated altitude of 5450 m had a greater cardiac output than unacclimatized bar-headed geese (by ∼200 mL/min) (Black and Tenney, 1980a). Similarly, bar-headed geese raised at high altitude increased cardiac output in response to diminished oxygen sooner than bar-headed geese raised at sea level (Lague et al., 2016), further demonstrating the importance of developmental phenotypic plastic responses, in addition to acclimatization for increasing the capacity to deliver oxygen through both increased ventilation and cardiac output (Monge and León-Velarde, 1991; Storz et al., 2010).
Blood properties
In addition to acute hypoxia responses, humans also make hematological adjustments after days to weeks at high altitude to sustain oxygen delivery to systemic tissue, typically by increasing hematocrit and total body hemoglobin mass (Fig. 2C), and therefore increasing arterial oxygen content (reviewed in Monge and León-Velarde, 1991). This also appears to occur as a fixed, evolved response to high-altitude hypoxia in humans native to the high-altitude Andes (Beall et al., 1998). The cost benefit of this is uncertain; despite the potential increase in arterial oxygen content, the resulting increase in blood viscosity creates a greater cardiac load, which may impair microcirculation, thus ultimately decreasing oxygen delivery to peripheral tissues (for reviews see Monge and León-Velarde, 1991; Storz et al., 2010; Barve et al., 2016). At its extreme, prolonged elevation of hematocrit (secondary to central hypoventilation or increased bone marrow erythropoietin production) can be associated with chronic mountain sickness in humans (reviewed in Villafuerte and Corante, 2016) and may perhaps explain why this adaptation is not seen in humans native to Tibet (Beall et al., 1998) or other high-altitude adapted mammals such as Andean camelids (reviewed in Monge and León-Velarde, 1991).
Low-altitude species of birds also increase hematocrit when experimentally exposed to simulated high altitude for prolonged periods (e.g., 3 weeks or more) (Mcgrath, 1971; Weatheres and Snyder, 1974; Weinstein et al., 1985). A strong erythropoietic response was found in Pekin ducks that underwent a 4-week acclimatization to a simulated hypobaric hypoxia of 5450 m (Black and Tenney, 1980b), and hematocrit levels of wild citril finches (Serinus citrinella) increased by 6% after ascending to 2000 m (Borras et al., 2010). In addition to hypoxia, high-altitude environments impose multiple challenges on vertebrates that may moderate blood properties (Morton, 1976). For example, the increased oxygen demand for thermogenesis in the colder environment at high altitude may also play some role in inducing an erythropoietic response; indeed lower temperatures experienced by wild citril finches following ascents to 2000 m were suggested to contribute to the increase in hematocrit levels (Borras et al., 2010). Thus, hypoxia alone may not be the sole driver for hematological changes.
In contrast to the Pekin duck erythropoietic response, increases in hematocrit level were not seen in bar-headed geese when exposed to a simulated altitude of 5450 m (Black and Tenney, 1980b). This may be because other adaptations in bar-headed geese, as well as some other bird species, sufficed to maintain oxygen delivery (reviewed in Butler, 2010; Scott et al., 2015b; Monge and León-Velarde, 1991), thus avoiding the increased cardiac load that results from increasing hematocrit (Black and Tenney, 1980b). This also appears to be the case in Tibetan natives and other high-altitude nonhuman mammals that have a blunted erythropoietic response to acute hypoxia, such as deer mice (Lui et al., 2015).
A range of species living at high altitude have hemoglobin with an increased affinity for oxygen (McCracken et al., 2009b, 2010; Natarajan et al., 2015). Both bar-headed geese and Andean waterfowl have a lower P50 (the PO2 at which hemoglobin is 50% saturated) relative to other avian species (Black and Tenney, 1980b; Liang et al., 2001; Meir and Milsom, 2013), although the mutation that causes this higher affinity in bar-headed geese differs from that in Andean waterfowl (Leu-β55 to Ser-β55 in Andean geese compared with Pro-α119 to Ala-α 119 in bar-headed geese) (Weber et al., 1985). Camelids and high-altitude populations of deer mice have also been found to have hemoglobin with a higher affinity for oxygen than their low-altitude counterparts (Storz, 2007). However, the overall increase in oxygen consumption that can be achieved through a high oxygen affinity is the result of a trade-off between oxygen loading and unloading, sensitive to the altitude experienced (Storz, 2007). A recent comparative analysis demonstrated that there was ambiguous evidence for increased hemoglobin oxygen affinity in high-altitude mammals, although across avian species, there was a strong positive relationship between bird species' native elevation (taken as the mid-point of each species' elevation range) (Projecto-Garcia et al., 2013) and hemoglobin oxygen affinity (Storz, 2016). This suggests that high-altitude selection pressures on birds act to increase hemoglobin oxygen affinity and this contributes to high-altitude birds' ability to sustain adequate oxygen supply without incurring the costs of elevated hematocrit levels that lowland birds and mammals experience.
Cerebral Oxygen Delivery and High-Altitude Cerebral Edema
Human lowlanders respond to hypoxia-induced decreased PaO2 with temporarily increased cerebral blood flow. Upon initial exposure to high altitude, cerebral vasculature dilates and for a short period, blood flow and oxygen delivery to the brain increase to compensate for reduced PaO2 (reviewed in Sanborn et al., 2015). However, this increase is subsequently diminished as a result of other acute hypoxia responses. First, increased ventilation leads to hypocapnia, which causes vasoconstriction of cerebral blood vessels (Fig. 2D). Second, other acclimatory changes, such as increased hematocrit, result in greater arterial oxygen content, so that cerebral blood flow can decrease without diminishing total cerebral oxygen delivery (for review see Hoiland et al., 2018). Birds' response to high-altitude hypoxia also begins with increased ventilation and a dilation of the brain vasculature so that cerebral blood flow is increased. However, an insensitivity of avian brain tissue to hypocapnia means that avian cerebral vasculature does not appear to vasoconstrict in response to the ensuing hypocapnia caused by increased ventilation, so cerebral blood flow is not diminished following prolonged hypoxic exposure (Faraci et al., 1985; Faraci, 1991) (Fig. 2D).
In humans, alterations to cerebral blood flow in response to hypoxia have been implicated in the onset of high-altitude cerebral edema (HACE) in climbers. HACE is thought to be a progression of acute mountain sickness that is marked by ataxia, headache, diminished conscious level, and ultimately, death (reviewed in Luks et al., 2017). It has been suggested that HACE may result from localized differences in cerebral blood flow, or where vasodilation and increased inflow are not matched to venous outflow (Wilson and Imray, 2016), leading to microhemorrhages and a diverse range of symptoms (Sagoo et al., 2017). In birds, the lack of vasoconstriction in response to hypocapnia means that cerebral blood pressure should not increase. despite the increase in cerebral blood flow (Grubb et al., 1977, 1978). Furthermore, avian neurons are less sensitive to hypoxia than mammalian neurons (Ludvigsen and Folkow, 2009); consequently, HACE is not known to occur in birds (Scott, 2011).
Skeletal Muscle Oxygen Delivery and Use
At the tissue and cellular level, further adaptations and acute hypoxia responses improve oxygen utilization in hypoxia in both mammals and birds. Human lowlanders visiting high altitude over a period of 1–2 months (i.e., the typical duration of an expedition in the Himalayas) undergo several muscular changes, including a reduction in cross-sectional surface area of muscle fibers (Hoppeler et al., 1990; MacDougall et al., 1991), and thus an improvement in oxygen diffusion as the capillary bed serves a smaller muscle volume (Hoppeler and Vogt, 2001). In addition, during a typical expedition to high altitude, the oxidative capacity of lowlanders' skeletal muscle also decreases. Oxidative capacity is the ability to consume oxygen and oxidize substrates, independent of the oxygen availability within the cell. One way this can be reduced is through a smaller volume density of mitochondria, which has been found to occur in human lowlanders who undergo a reduction of up to 30% in the subsarcolemmal or interfibrillar mitochondria following expeditions to high altitude (Hoppeler et al., 1990). Furthermore, several respiratory enzymes, including citrate synthase, cytochrome oxidase (Howald et al., 1990), succinate dehydrogenase, and hexokinase activities, also decrease (Green et al., 1989; MacDougall et al., 1991). This serves to potentially reduce production of harmful reactive oxygen species (Hoppeler et al., 2003). For athletes wishing to incur some of the benefits of exposure to high altitude (i.e., increased oxygen-carrying capacity of the blood) without incurring these muscular costs, a schedule of intermittent hypoxia may be required (reviewed in Dempsey and Morgan, 2015). Total oxygen consumption in human high-altitude residents may be enhanced relative to that in lowlanders through increased oxygen delivery, achieved by greater capillary to muscle fiber ratios (Kayser et al., 1991) and higher rates of oxygen consumption per mitochondria. This may be the result of an enhanced efficiency in adenosine triphosphate (ATP) turnover achieved through coupling of ATP supply and demand pathways (Hoppeler and Vogt, 2001; Hoppeler et al., 2003).
However, birds have greater mass-specific than mammals, and this is supported by up to twice the capillarity in locomotor muscles than mammals (Fig. 2D) (Faraci, 1991; Mathieu-Costello et al., 1992), meaning that oxygen delivery to the muscles is more effective, as diffusion distances are reduced. This advantage is increased further in bar-headed geese, which have significantly higher capillary densities and higher capillary to muscle fiber ratios at the surface of flight muscles, relative to those in low-altitude waterfowl species, facilitating higher levels of oxygen delivery required during high-altitude flights (Scott et al., 2009, 2011).
Furthermore, avian flight muscle has a higher oxidative capacity and greater ability to metabolize fat than mammals (Rosser and George, 1986; for reviews see McGuire and Guglielmo, 2009; Butler, 2016). When mammals are exercising at maximal rates, fatty acids supply ∼15% of the fuel metabolized (Weber et al., 1996). However, in birds, 90% of energy demand may be derived from fatty acids, made possible by a heightened capacity to transport fatty acids through an in increase in fatty acid transport proteins on the membranes of the muscles relative to mammals (Fig. 2E) (reviewed in Guglielmo, 2010). This appears to be an adaptation necessary to sustain the high oxygen demands of flight (Tucker, 1970), rather than a response to hypobaric hypoxia per se. It is yet to be shown if greater fatty acid metabolism is beneficial at high altitudes, given the higher oxidative cost of metabolizing fatty acids; oxidizing lipids becomes increasingly expensive relative to glycogen with increasing altitude (Melzer, 2011; Yap et al., 2018). Enzymes involved in the metabolism of fatty acids were not found to vary across the locomotor muscles of high- and low-altitude populations of torrent duck (Dawson et al., 2016). However, high-altitude populations of deer mice have been found to have greater proportions of type I muscle fibers than low-altitude populations, which may increase their capacity to transport fatty acids (Lui et al., 2015). Furthermore, relative to low-altitude cattle, yaks have been shown to undergo modifications in five key genes in nutrition pathways relating to fatty acid metabolism (Qiu et al., 2012). These studies suggest that fatty acid metabolism may be selected for or enhanced in high-altitude environments in mammals, but it is not yet clear if the same is true in birds, which already have a higher capacity to metabolize fatty acids than mammals (Lui et al., 2015; for review see Storz et al., 2019).
Avian high-altitude species can further increase oxidative capacity at the cellular level. High-altitude resident populations of torrent duck have significantly increased activities of cytochrome oxidase, phosphofructokinase, pyruvate kinase, and malate dehydrogenase in the locomotor muscles relative to those in ducks living at low altitude (Dawson et al., 2016). It is unknown whether these adaptations are common among other high-altitude adapted bird species (respiratory enzyme activity has not been studied in many other species to date), but other adaptations at the cellular level have been described in bar-headed geese. These include a redistribution of mitochondria closer to the capillaries, relative to those in low-altitude waterfowl, which is the opposite response of low-altitude humans acutely exposed to high altitude, who undergo a reduction in subsarcolemmal mitochondria (Hoppeler et al., 1990; Scott et al., 2009, 2011).
Similar adaptations have also been described in high-altitude populations of deer mice, including increased capillarity, a redistribution of mitochondria to reduce the intracellular oxygen diffusion distances, and increased activity of key respiratory enzymes (Lui et al., 2015; Scott et al., 2015a; Mahalingsam et al., 2017; for review see McClelland and Scott, 2018). Therefore, chronic high-altitude hypoxia exposure appears to have acted in both birds and mammals to enhance the oxidative capacity as well as oxygen delivery to skeletal muscle. Such increases in oxidative capacity at the skeletal muscle contribute to greater overall maximum oxygen consumption (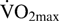 ). At the taxon level, birds appear to be able to support a higher than mammals (Fig. 2), which is beneficial at high altitudes characterized by both reduced oxygen availability and increased thermoregulation demands, given the cold challenge.
). At the taxon level, birds appear to be able to support a higher than mammals (Fig. 2), which is beneficial at high altitudes characterized by both reduced oxygen availability and increased thermoregulation demands, given the cold challenge.
Conclusion
Birds' ability to sustain the high metabolic rates to facilitate the demands of flight likely confer benefits that allow birds to better tolerate hypoxia relative to mammals, including humans. Species and populations of birds and mammals that are regularly exposed to high altitudes have adaptations to enhance oxygen delivery, including greater HVR and greater oxygen-carrying capacity of the blood (reviewed in Butler, 2010; Scott, 2011; Ivy and Scott, 2014; Scott et al., 2015b). More recent research has shown that high altitude can also act as a strong selection pressure for greater oxidative capacity at the cellular level across both birds (Dawson et al., 2016; Yap et al., 2018; Hao et al., 2019) and mammals (Storz, 2007; McClelland and Scott, 2018). Despite these shared features, the advantages of the avian lung, hemoglobin with a high oxygen affinity and insensitivity to hypocapnia, triumph. Recent comparisons of high-altitude adaptations in resident waterfowl in the Andes and migratory bar-headed geese reveal that different mechanisms can serve to increase hypoxia tolerance (Laguë et al., 2017; reviewed in Laguë, 2017). However, there are common features unique to birds that visit high altitudes, which include more effective gas exchange with a greater HVR, larger lungs with higher capillarization, enhanced cardiac output, greater oxygen delivery to skeletal muscle due to increased hemoglobin oxygen affinity, and higher capillary to fiber ratios with increased oxidative capacity. Indeed, on rare occasions, several species of birds have been detected flying at exceptional altitudes (Noel, 1927; Laybourne, 1974; Hawkes et al., 2012; Parr et al., 2017). While extreme high-altitude flight may not represent their typical behavior, the question remains as to how high the best adapted avian species could reach, possibly with the assistance of orographic lift (Bishop et al., 2015).
Footnotes
Authors' Contributions
All authors contributed to the conception of the article; research and first drafting of the article were conducted by N.P. M.W. and L.A.H. contributed to the subsequent writing and editing. All authors have reviewed and approved the article for submission.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
N.P. was supported by a PhD studentship from the University of Exeter.