
Abstract
Simmonds, Michael J., Surendran Sabapathy, and Jean-Marc Hero. Rate-pressure product responses to static contractions performed at various altitudes. High Alt Med Biol. 22: 166–173, 2021.
Background:
Adventure tourism has led to an unprecedented number of individuals being exposed to altitude, including those with subclinical cardiometabolic disorders. The disproportionate hemodynamic challenge associated with small-muscle static activities is potentially dangerous at altitude as these may compound the risk for cardiac events. We thus examined the cardiovascular response to, and during recovery from, static exercise performed at altitude.
Methods:
Eighteen individuals completed this study at three altitudes (sea level; ∼1,500 m; ∼3,000 m) in central Nepal. At each altitude, individuals performed two handgrip contractions for 2 minutes at the same intensity (30% maximal voluntary contraction [MVC]), with two distinct recovery periods: during control recovery was completed quietly at rest, while during ischemic challenge recovery was conducted with a cuff occluding the upper limb.
Results:
Oxygen saturation decreased during ascent to 1,500 m (−2%) and 3,000 m (−8%), compared with sea level. Handgrip MVC was not affected by altitude, although heart rate at rest (∼70 beat/min), during static exercise (range ∼90–95 beat/min), and during recovery in both conditions (each ∼70 beat/min) was significantly increased by ∼15% at 3,000 m, but not 1,500 m. The magnitude of the muscle metaboreflex during recovery from static exercise was unaffected by altitude; however, the rate-pressure product was significantly elevated by ∼10% during and following static exercise at 3,000 m.
Conclusions:
A significant increase in the rate-pressure product during static exercise was observed at altitude, which persisted during recovery. Individuals at risk for cardiac events should use awareness of static contractions while at altitude, especially considering that stress induced by static exercise is additive to that of dynamic activities such as hiking.
Introduction
Ascent to altitude is associated with various stressors (e.g., cold exposure; dehydration) that may be easily managed with careful preparation, while others (e.g., hypoxia) require adaptive responses within the organism. Adaptive responses by the physiological systems typically involve two phases: acute responses may last days-to-several weeks, and then chronic adaptations (i.e., acclimatization) may be observed (for more details, see Bartsch and Gibbs, 2007). Nonacclimatized individuals, upon sojourn to high altitude, tend to demonstrate signs of sympathoexcitation, including transient increases in systemic blood pressure (Rhodes et al., 2011; Schultz et al., 2014) and augmented sympathetic nerve activity (Duplain et al., 1999).
This rise in blood pressure is altitude dependent and the net result of contrasting effects of vasoconstriction resultant from increased sympathetic tone and low arterial PO2 inducing relaxation of the resistance vessels (i.e., hypoxic vasodilation); after an initial balance of these two competing stimuli during the first weeks of adaptation, a gradual increase in vascular resistance occurs that reflects continued “sympathetic dominance.” This sympathetic dominance and the combined effects of increased systolic blood pressure and heart rate indicate an increased myocardial demand during short-term visits to altitude; an effect that may persist for at least 7 weeks (Hansen and Sander, 2003; Lundby et al., 2018). An estimate of myocardial work may be simply determined using the rate-pressure product (systolic blood pressure × heart rate), given this index provides a proxy dependent upon the frequency of cardiac contraction and also the force against which the myocardium must contract (Nelson et al., 1974). Limited evidence has demonstrated that during equivalent submaximal efforts, this index of myocardial work significantly increased by over 200% at altitude, when compared with sea level (Nishihara et al., 1998).
The cardiovascular response to static work at sea level is proportional to relative intensity—specifically, the percentage of maximal voluntary contraction (MVC)—and also the mass of contracting muscle (Mitchell et al., 1980). Moreover, relative to the modest muscle mass recruited, and the comparatively low work rates (e.g., 5%–30% MVC), there is a considerable increase in systemic blood pressure, in addition to heart rate, that may be observed until task failure (i.e., end exercise) (Lind and McNicol, 1967; Hietanen, 1984). Lind and McNicol (1967) specifically demonstrated that the cardiac stress associated with static contractions is additive to that induced by dynamic exercise; when static handgrip contractions were performed at 30% MVC in individuals walking on a treadmill, the chronotropic and pressor responses increased by ∼50% over-and-above that induced by treadmill walking alone. These data support that an exceptional rate-pressure product, and thus myocardial demand, is induced during static exercise relative to the total work being performed. Furthermore, static work limits local blood flow in the active limb due to the associated extravascular pressures and thus increases metabolite accumulation—which is a potent stimulant of the exercise “muscle metaboreflex” (Alam and Smirk, 1937; Boushel, 2010). While elevated sympathetic response to static work is well-described at sea level, limited studies have been completed at altitude, where an elevated baseline sympathetic tone may be observed owing to reduced arterial PO2. On the contrary, more is known regarding dynamic exercise responses while at altitude; simulated and actual exposure to altitude demonstrates that the sympathetic dominance observed at rest is compounded with an exaggerated sympathetic response to moderate exercise (Woods et al., 2017), thus presenting a potential risk for cardiac events. This observation is concerning, given that static exercise produces a disproportionate sympathetic response relative to the muscle mass involved when compared with dynamic exercise—even at sea level (Hietanen, 1984).
Given that many tasks during mountaineering involve isometric muscle tasks—for example, use of walking poles and load carriage—the cardiovascular response to isometric muscle contractions is more than a scientific curiosity, particularly given that static contractions evoke an additive stress response over-and-above basal responses during dynamic exercise (Lind and McNicol, 1967). The burgeoning tourism industry associated with summit tours/hiking (Horiuchi et al., 2017) infers that a greater number of individuals likely to be at elevated risk of cardiovascular disorders will be exposed to the combined effects of environmental (e.g., hypoxia, cold exposure) and exercise stressors, which independently increase sympathetic tone. Given that the cardiovascular responses to isometric muscle tasks at altitude are poorly described, we aimed to study the effects of a dose/response in moderate altitude exposure (∼0/∼1,500/∼3,000 m) on the cardiovascular response to a standardized isometric exercise effort in previously altitude-naive individuals.
Materials and Methods
Subjects
Twenty-three volunteers (13 women) participated in the present study after providing written informed consent: 18 participants (23 ± 4 years; 10 women) completed the study at all three altitudes and are reported hereafter. Reasons for withdrawal from the study by the five noncompleting volunteers centered around preference to conduct other activities once at altitude that precluded their participation during the study window. Participants were selected from volunteers who were partaking in a 4-week conservation field-study throughout Nepal that included multinight sojourns at elevations of interest (i.e., <100 m; 1,500 m; 3,000 m). Participants ascended into the Nepalese Himalayas via walking expedition and achieved all elevations without assistance. Before being accepted as participants, volunteers were screened via a brief clinical examination and interview to determine suitability for the present study. Participants were free from diagnosed metabolic, respiratory, neurological, and cardiovascular disorders, and were not using medications known to influence the cardiovascular responses to exercise (e.g., β-adrenergic antagonists). Current health status was assessed at the onset of each experimental session. The experimental procedures were reviewed and approved by the Griffith University Human Research Ethics Committee.
Experimental overview
The present experimental procedures were conducted at least 7 days after international travel, and all participants were free of acute illness on each testing session. The testing sites were chosen a priori based on published mountain elevations within the Nepalese Annapurna mountain range and Chitwan National Park. At the onset of each experimental testing session, the elevation (m) of each testing site was confirmed via an electronic altimeter (Ambit2; Suunto Oy, Finland). Resting measurements of heart rate, blood pressure, and peripheral capillary oxygen saturation (SpO2; %) were recorded while seated and rested at each testing site. Handgrip strength testing was subsequently conducted while in the seated position; this measurement was used to characterize the participant's handgrip strength, and was also used to provide an individualized relative-intensity during subsequent sustained handgrip contraction experiments.
Experimental testing sessions
All testing sessions were scheduled and completed at 07:30 ± 01:30 hours, and participants avoided exercise and caffeine-containing products for 12 hours before testing. Each testing session was conducted within 12 hours of reaching the specific elevation (i.e., 1,500 and 3,000 m) and after an overnight sleep. Each experimental testing session commenced with a brief explanation of the study protocols while the participant was seated. Resting SpO2 was recorded via a portable pulse oximeter (Onyx Vantage 9590; Nonin Medical, Inc.) that was attached to the second digit of the participant's nondominant hand. Heart rate was recorded in the resting position from a heart rate monitor chest-strap (Ambit2; Suunto Oy) and was confirmed with pulse rate that was concurrently displayed on the pulse oximeter: all heart rate measurements reported were recorded using the heart rate monitor. Blood pressure was then obtained manually via auscultation of the brachial artery of the nondominant limb using a calibrated sphygmomanometer (Tycos DS58; Welch Allyn) and stethoscope (Littman Classic II S.E.; 3M Health Care): all blood pressure investigations were conducted and recorded by the same experienced investigator (M.J.S.) and reported hereafter as mean arterial pressure (MAP), determined using the equation: MAP = diastolic blood pressure +1/3 × (pulse pressure).
Handgrip strength was determined using a calibrated dynamometer (Jamar Hydraulic Hand Dynamometer; Sammons Preston Rolyan). Participants were provided a demonstration on the use of the instrument by the investigators, before the appropriate hand-size settings were determined for each individual. Once the participant had an opportunity to become accustomed to the feel of the dynamometer, the posture of the participant was corrected to ensure that feet were placed apart on the ground, a straight back was maintained, and the elbow of the dominant/active limb was flexed at 90°. The investigators then vigorously encouraged the participant to provide maximal effort for ∼3 seconds during a handgrip contraction. The highest force produced during this effort was immediately recorded and the participant was provided ∼30–60 seconds of rest, before the maximal handgrip contraction was repeated on further two occasions. The peak force produced during the three consecutive handgrip strength tests was considered the MVC.
The participants were then provided a description of the experimental protocols and instructed to avoid breath holding or Valsalva manoeuvers during the static contractions. Participants were also coached on the requirement for relying on auditory feedback from a research assistant for providing real-time adjustment of the instantaneous force output that was to be used during the static muscle contraction: this method of feedback was required given that the dial of the handgrip dynamometer is oriented away from the participant and toward the investigators. Once participants were able to demonstrate an ability to maintain a steady submaximal force output as directed by the research assistant for at least 10 seconds, the experimental protocols were commenced.
Immediately before each experimental test, the participant's posture was corrected as per during the determination of MVC, and participants were reminded to maintain normal breathing throughout the test. The two experimental trials involved participants conducting (i) a sustained submaximal handgrip muscle contraction at 30% MVC for 2 minutes, and then quiet rest for 2 minutes following cessation of muscle contraction (“CON” trial); and (ii) an identical sustained muscle contraction per CON; however, at 1:55 minutes of the 2-minute muscle contraction, a sphygmomanometer cuff was inflated to >80 mm Hg above systolic blood pressure at the level of bicep brachii of the active limb—on average, the cuff was inflated to ∼220 mmHg. The cuff remained inflated for the 2-minute recovery period (i.e., ischemic recovery; ISC) for this trial. The cuff inflation during recovery of the ISC trials facilitated investigation into the so-called muscle metaboreflex at various altitudes; classic works support that both feedforward innervation and metabolite stimulation of afferent nerves local to the exercising muscle, each determines the cardiovascular response to exercise (Alam and Smirk, 1937). This method thus provided insights into whether the feedforward and/or feedback responses were altered by sojourn to altitude. Between repeated experimental trials—that is, CON and ISC—recovery was provided to facilitate blood pressure and heart rate returning to resting values, which was confirmed before the second trial. Throughout the experimental trials, beat-by-beat heart rate was recorded continuously and blood pressure was measured via auscultation of the brachial artery at minute 1 and 2 of the sustained muscle contraction, and also at minute 1 and 2 of the recovery period.
Statistical analysis
Results are reported as mean ± standard error for each of the two trials and three different conditions. Normality of the data was tested using the D'Agostino and Pearson omnibus normality test (Prism, Release 6.03; GraphPad Software, Inc.). Data for each condition were compared using a multifactorial analysis of variance with repeated measures to determine whether significant differences in the means existed, and linear regression was performed to examine relationships between specific dependent variables (Prism, Release 6.03; GraphPad Software, Inc.). Significance was determined at an alpha level of 0.05.
Results
The individual and mean SpO2 responses to acute exposure to altitude are illustrated in Figure 1. All participants presented with SpO2 measurements within the normal range for healthy individuals while at sea level (99.1% ± 0.7%). The sojourn to altitude significantly decreased SpO2 in a nonlinear manner (F = 206, p < 0.0001) when compared with sea level; an overnight visit to 1,500 m resulted in a −1.8% ± 1.0% decrease in SpO2 (p < 0.001), while a much larger decrement (−8.3% ± 2.0%) was observed at 3,000 m (p < 0.0001). The SpO2 measured at 3,000 m was significantly lower than that observed at 1,500 m (p < 0.0001).

Oxygen saturation (SpO2) of arterial blood measured while at rest during exposure to SL, and at 1,500 and 3,000 m altitude for 18 volunteers. Individual responses are plotted in gray, while the pooled sample data are represented in black. aSignificantly different to SL; b3,000 m significantly different to 1,500 m (both at least p < 0.001). SL, sea level.
The peak handgrip force obtained during the three MVC trials is presented in Figure 2. No significant difference was detected in forearm MVC at any altitude. No significant relationship was detected between SpO2 while at rest and forearm MVC force production at 3,000 m (r = −0.17, p = 0.20), nor was a significant relationship observed in the decrement in SpO2 (Δ−8.3% ± 2.0%) and delta forearm MVC force production (Δ0.5 ± 3.0 kgf) measured at 3,000 m and sea level (r = −0.06, p = 0.81).
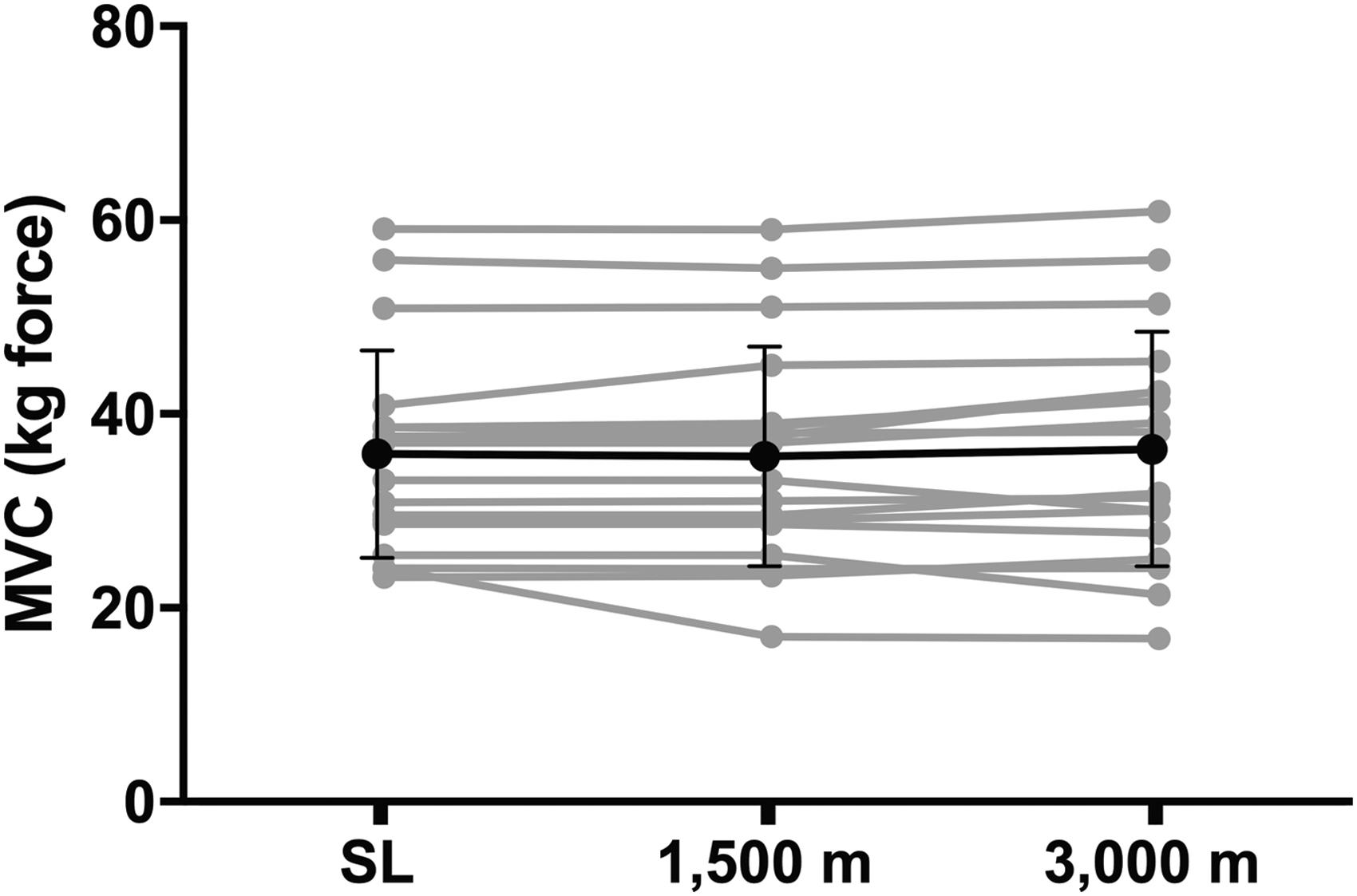
Forearm MVC force production while at SL and following overnight exposure to 3,000 m altitude for 18 volunteers. Individual responses are plotted in gray, while the pooled sample data are represented in black. MVC, maximal voluntary contraction.
Heart rate response
The heart rate responses to the 2-minute static handgrip contractions, and the subsequent recovery period without (CON) or with an occlusive cuff (ISC), at sea level, 1,500, and 3,000 m, are presented in Figure 3. The general pattern of heart rate response to static exercise (Fig. 3, upper) was consistent for all trials, although at different magnitudes: the 2-minute static handgrip contraction significantly increased heart rate. Moreover, a similar trend in the decrease of heart rate during the recovery period was observed for all trials: heart rate decreased significantly during recovery from the 2-minute static handgrip contraction.

Heart rate response to forearm static exercise and during recovery, while at SL, and following overnight exposure to 1,500 and 3,000 m altitude (upper) for 18 volunteers. Static exercise significantly increased heart rate (middle), and recovery decreased heart rate at all three altitudes. Forearm occlusion (ISC) during recovery did not affect heart rate (lower). Overnight exposure to 3,000 m increased heart rate at all time points, irrespective of ISC or CON trials, when compared with SL. Three thousand meters significantly elevated to SL: *p < 0.05; ***p < 0.001. aSignificantly different to SL; b3,000 m significantly different to 1,500 m. Note: data are tightly clustered at each altitude, particularly at 3,000 m—thus the open and filled triangles are almost superimposed. CON, control; ISC, ischemic recovery.
A main effect for altitude was detected (F = 19.8, p < 0.0001), where heart rate was significantly increased at 3,000 m when compared with 1,500 m, and sea level. During the CON trials, exposure to 3,000 m significantly increased heart rate at rest (p = 0.02), at 60 seconds (p < 0.0001), and 120 seconds (p < 0.0001) of static handgrip contraction, and after 60 seconds (p < 0.0001) and 120 seconds (p < 0.0001) of recovery, when compared with sea level. For the ISC trials, heart rate was significantly elevated at 3,000 m compared with sea level measured at rest (p = 0.02), at 60 seconds (p < 0.0001) and 120 seconds (p < 0.0001) of static handgrip contraction, and after 60 seconds (p < 0.0001) and 120 seconds (p < 0.0001) of recovery while the arm cuff was inflated to induce upper-limb ischemia.
The magnitude of change (i.e., Δchange) in heart rate from rest to the end of the 2-minute static handgrip contraction (Fig. 3, middle) was significantly increased due to the sojourn to altitude (F = 10.3, p < 0.001). The delta change in heart rate measured after 2 minutes of static handgrip exercise compared with rest was significantly increased at 1,500 m (p = 0.04) and 3,000 m (p < 0.0001) when compared with sea level. Moreover, performing the 2-minute static handgrip contraction at 3,000 m induced a significantly increased delta change in heart rate (end exercise—rest) when compared with 1,500 m (p = 0.01).
The decrease in heart rate (i.e., Δchange) observed during the recovery period compared with end of exercise (i.e., 2-minute recovery—end exercise) was not significantly different after exposure to altitude, when compared with sea level (Fig. 3, lower; p = 0.35).
Mean arterial pressure
The MAP response to static exercise (Fig. 4) shared a similar trend; there was a significant increase from rest after 60 seconds (p < 0.0001) of handgrip exercise, with a further increase by 120 seconds (p < 0.0001). However, during recovery from static exercise, a divergent response could be observed; MAP either: (i) decreased rapidly toward resting levels within 60 seconds of terminating exercise during CON; or (ii) decreased gradually, remaining elevated when compared with resting levels even after a 120-second recovery with ISC. Interestingly, altitude exposure did not influence the magnitude of increase in MAP during static exercise (Fig. 4, middle) nor influence the dampened decrease in MAP observed during recovery for the ISC (Fig. 4, lower).
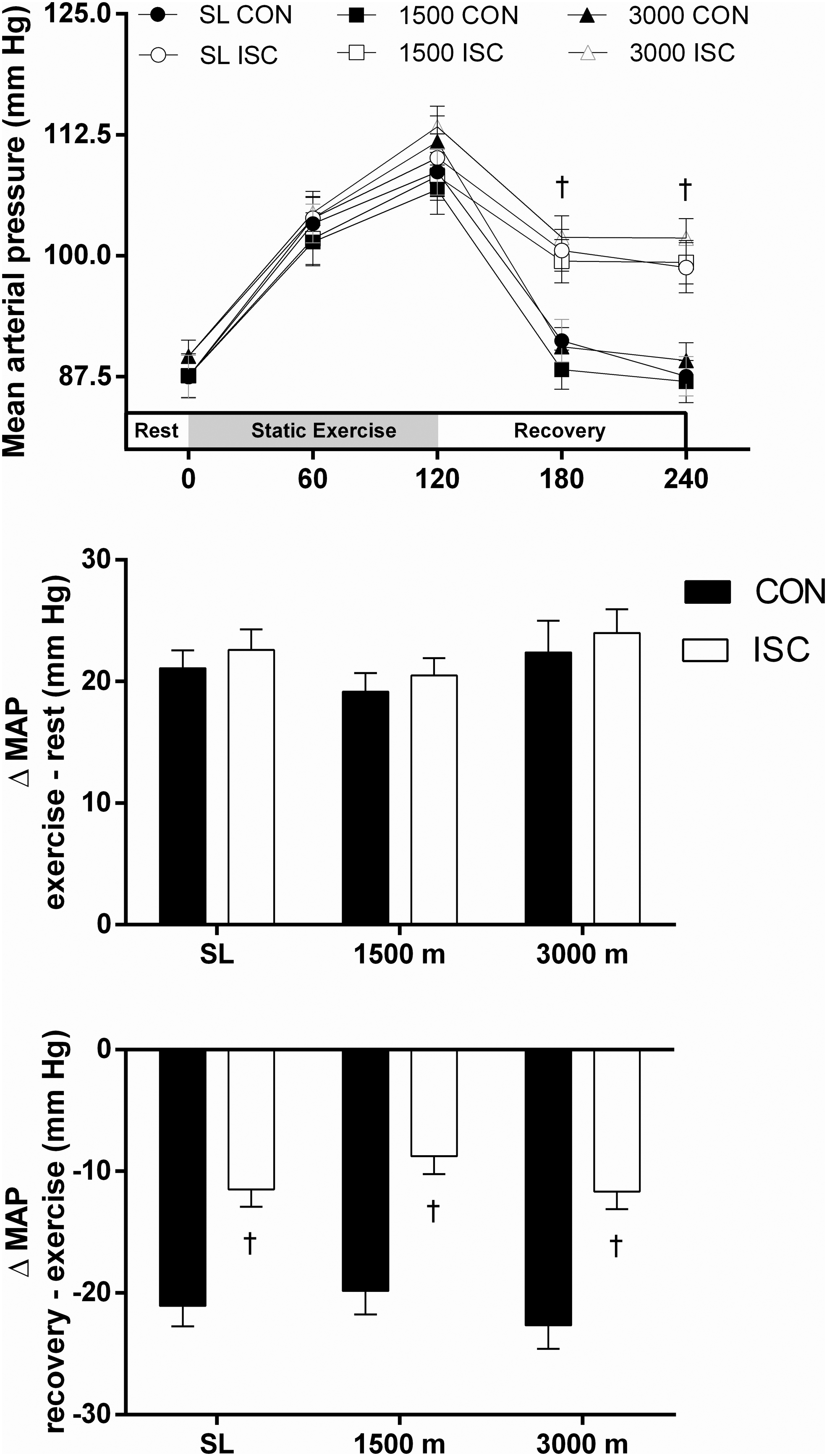
MAP response to forearm static exercise and during recovery, while at SL, and following overnight exposure to 1,500 and 3,000 m altitude for 18 volunteers. Static exercise significantly increased MAP, and recovery decreased MAP at all three altitudes (upper). The increase in MAP was not affected by altitude (middle). Forearm occlusion (ISC) during recovery significantly dampened the decrease in MAP at all three altitudes (upper and lower). †ISC significantly different to CON. MAP, mean arterial pressure.
Rate-pressure product
The rate-pressure product response to static handgrip exercise, with and without ischemic forearm occlusion during recovery, is presented in Figure 5. For all conditions, exercise progressively increased the rate-pressure product with exercise duration, and thus, myocardial demand during static exercise increased significantly after 60 seconds (p < 0.001) and 120 seconds (p < 0.001), when compared with rest. The rate-pressure product then progressively decreased during recovery for all conditions, although both altitude and ISC affected the time course of recovery.

The rate-pressure product response to static handgrip exercise without (CON; left) and with ischemic forearm occlusion (ISC; right) during recovery, while at SL, and at 1,500 and 3,000 m altitude for 18 volunteers. For clarity, significant differences discussed in-text.
Exposure to 1,500 m had minimal effect on the mean response for the rate-pressure product when compared with sea level; however, it was particularly evident that exposure to 3,000 m had a profound effect on the rate-pressure product, in both CON and ISC. At most time points, exposure to 3,000 m significantly increased the rate-pressure product, as evident through significant differences observed after 60 seconds (p < 0.001) and 120 seconds (p < 0.001) of static exercise when compared with both 1,500 m and sea level, for both CON and ISC.
With respect to altitude, recovery from static exercise was rapid in CON when performed at sea level, where no difference in rate-pressure product could be detected after 60 seconds of recovery (p = 0.472), when compared with rest. Overnight exposure to altitude, however, prolonged the recovery period; the rate-pressure product had not returned to basal levels even after 120 seconds of recovery at 1,500 m (p < 0.01) and 3,000 m (p < 0.0001). With respect to ISC, it was found that recovery from exercise was not complete even after 120 seconds of recovery at any altitude, including sea level, in the presence of ischemic occlusion; ISC resulted in a substantially elevated rate-pressure product at all altitudes after 60 seconds (all p < 0.001) and 120 seconds (all p < 0.001), when compared with rest. The response during recovery for the ISC trial was elevated at 3,000 m, even when compared with 1,500 m, after 60 seconds (p < 0.001) and 120 seconds (p < 0.001).
Discussion
While exposure to altitude was, historically, relatively restricted to those living within proximity of isolated mountain ranges with inherited adaptations to cope with the extreme environment, a burgeoning market for adventure tourism has led to an unprecedented number of nonacclimatized individuals experiencing the acute effects of altitude. We observed that SpO2 decreased in a nonlinear manner when measured at rest during a stepwise sojourn to moderate altitude in nonacclimatized individuals; specifically, exposure to 3,000 m had a proportionally larger effect on blood oxygen saturation when compared with 1,500 m (∼8% vs. ∼2% reduction, respectively). This decrease in SpO2 during acute altitude exposure coincided with several pertinent findings: (i) handgrip MVC was not affected at altitude, when compared with sea level; (ii) heart rate was significantly elevated at rest, during static handgrip exercise, and during recovery from static exercise performed at 3,000 m, but not 1,500 m; (iii) MAP was not affected at rest, during static exercise, or during recovery from static exercise; (iv) the magnitude of the “muscle metaboreflex” (discussed later) during recovery from static exercise was unaffected; and (v) the rate-pressure product was significantly elevated during and following static exercise. Collectively these data support that there is a critical altitude that leads to a dramatic decrease in blood SpO2 and manifests several responses indicative of sympathetic dominance. In the present study, the rate-pressure product—an indirect measure of myocardial demand—was significantly elevated during static exercise, which persisted during the recovery period from this activity. Given that static exercise is regularly performed during daily activities, and indeed hiking, the additive cardiovascular stress induced by isometric contractions should be considered during sojourns to altitude.
Acute exposure to moderate altitude induced a stepwise decrease in blood oxygen saturation, as expected, in the present study, where SpO2 decreased by ∼2% at 1,500 m and ∼8% at 3,000 m, when compared with sea level. This classic observation can be attributed to the associated reduced driving pressure (i.e., barometric pressure) for pulmonary gas exchange. The magnitude of decrease in SpO2 measured at 1,500 m was modest when compared with previous findings (Tucker et al., 1984) in an hypobaric chamber, whereas the sojourn to 3,000 m induced a typical response for laboratory (Tucker et al., 1984) and environmental (Karinen et al., 2010) exposure to altitude. Furthermore, the individualized data presented in Figure 1 indicate that all participants experienced SpO2 responses within the reported range for young individuals (Rojas-Camayo et al., 2018).
Acute exposure to 3,000 m, but not 1,500 m, induced a significant increase in heart rate at rest, and also an altered chronotropic response during and following static exercise. This classic heightened chronotropic response cannot be explained by an altered intensity of static exercise at altitude; all conditions were performed at the same relative intensity, which was confirmed on the morning of each test. The consistency in the upward shift in the chronotropic response at all time points measured at 3,000 m supports a systemic modulator of heart rate. Elevated circulating concentrations of epinephrine and norepinephrine, known modulators of sympathetic tone, are consistently observed during acute exposure to hypoxia (Kontos and Lower, 1969). Furthermore, Busch et al. (2020) observed a significant increase in muscle sympathetic nerve output during exercise and also in ischemic recovery from handgrip exercise, comparable with the current study; these data clearly support that sympathetic reactivity is heightened during acute exposure to altitude.
Altitude did not affect MAP (Fig. 4), which confirms recent reports (Baertschi et al., 2016; Busch et al., 2020). Baertschi et al. (2016) demonstrated that MAP was not affected at 4,200 m, and was only significantly increased (although modestly) when individuals were exposed to 6,000 m. The working hypothesis for this observation was that any small increase in systolic blood pressure (via sympathetic tone) was offset entirely by dampened diastolic pressure (via hypoxic vasodilation), thus diminishing changes in MAP. Recent findings by Simpson et al. (2019) support this position; they suggested that lowlanders may offset heighted sympathetic nerve output when at altitude via humoral vasoactive compounds and modulation of vascular sensitivity. The competing effects of sympathetic innervation and hypoxic vasodilation (which may be both endothelium dependent and independent) lead to a complex systemic blood pressure response during acute exposure to altitude (Mazzeo et al., 1991). Intriguingly, the “muscle metaboreflex” (for review, see Boushel, 2010) to ischemic challenge during recovery from static exercise was unaffected by altitude (Fig. 4) supporting that any altitude-induced challenge to central and peripheral regulation of MAP was entirely overcome.
Perhaps the most profound observation of the present study was that the rate-pressure product was significantly increased at 3,000 m, particularly during and following static exercise. The rate pressure product is a reliable noninvasive and highly accessible index of myocardial demand, and generally agrees with findings using advanced medical imaging (Ansari et al., 2012). Static contractions performed while at altitude therefore present as a particularly unassuming, yet potentially hazardous, challenge if performed at high intensities and/or for extended durations in combination with dynamic exercise. In light of the concurrent decreased arterial oxygen content (SpO2), the sympathetic dominance and associated increase in myocardial oxygen requirements (per rate-pressure product) infer additional risk for myocardial ischemia during acute altitude exposure. This insight has real implications, given many tasks performed at altitude include prolonged static contractions, such as load carriage and even the use of walking poles. Currently, the risk of myocardial ischemia during acute exposure to altitude is unclear, although a recent case study details an individual who presented with a classic ischemic electrocardiogram during moderate exercise performed at 3,260 m, despite completing a negative cardiopulmonary exercise test (i.e., showed no signs of ischemic electrocardiogram) only days earlier at sea level (Caravita et al., 2014). It is thus clear that the extreme environment encountered during rapid ascent to even moderate altitude induces an unexpected burden that may present risk for those with poor cardiometabolic health (Hainsworth and Drinkhill, 2007).
The present study also demonstrated that ischemic occlusion of the active limb during recovery from static exercise blunted the postexercise fall in blood pressure. This is a well-described phenomenon that occurs due to partitioning of the central and peripheral stimuli that promote the exercise pressor response (Gandevia and Hobbs, 1990); while descending drive from the motor cortex ceases upon termination of exercise, the metabolite-rich blood that stimulates Group III/IV afferent feedback for modulating hemodynamics is “trapped” by the extravascular pressure induced by the sphygmomanometer (Seals, 1989). This situation leads to an elevated blood pressure until the metabolites are cleared following the removal of extravascular pressure. Our present study builds upon these findings, indicating that this response is not influenced by traversing to moderate altitude, which was unexpected given it was hypothesized that the already lower oxygen tension within peripheral blood at altitude may have exacerbated the Group III/IV afferent sensitivity. These findings highlight the robust defence of MAP, even in the face of environmental challenges.
In conclusion, our data indicate a need for consideration when conducting even seemingly mild activities involving static contractions at moderate-to-high altitude, especially when performed in tandem with dynamic exercise. Upon acute exposure to 3,000 m, our participants presented with substantial desaturation of blood oxygen levels, and an exaggerated chronotropic response to static exercise. The associated increase in myocardial demand, as inferred by the rate-pressure product, during and following static exercise at 3,000 m is a particular concern for individuals who may be at risk for cardiac events, including the increasingly numerous proportion of the population that fits the “metabolic syndrome” and yet remains subclinical for cardiac disease. Further verification of this thesis would be of value, particularly if direct measurements of electrical and/or biochemical markers of cardiac myocyte dysfunction were shown to increase due to the substantial myocardial load of isometric tasks while at altitude. It follows that clinicians providing suitability recommendations for adventure tourism involving altitude exposure, and indeed tourist themselves, should be educated on the risk of static exercise while at altitude, particularly given that even small muscle mass contractions induce a disproportionately large hemodynamic challenge. It is reasonable, for example, to suggest that professionals involved in designing acclimatization protocols should extend their service to guide patients/clients of the potential hazards of static exercise performed in extreme environments.
Footnotes
Acknowledgments
Special thanks to Narinder Virdee who organized and managed the logistics of the expedition, without whom this study could never have happened. Thanks to our Nepalese colleagues who assisted with the collaborative teaching program (Tribhuvan University): Prof Krishna K. Shrestha and Prof Khadga Basnet. We also thank Lal P. Gurung, Project Director for the Anapurna Conservation Area Project, who supported our expedition. Finally, we thank the volunteers who managed to successfully reach each checkpoint, and who also made the journey rewarding.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was procured for this research.