
Abstract
Committeri Giorgia, Danilo Bondi, Carlo Sestieri, Ginevra Di Matteo, Claudia Piervincenzi, Christian Doria, Roberto Ruffini, Antonello Baldassarre, Tiziana Pietrangelo, Rosamaria Sepe, Riccardo Navarra, Piero Chiacchiaretta, Antonio Ferretti, and Vittore Verratti. Neuropsychological and neuroimaging correlates of high-altitude hypoxia trekking during the “Gokyo Khumbu/Ama Dablam” expedition. High Alt Med Biol. 23:57–68, 2022.
Background:
Altitude hypoxia exposure may produce cognitive detrimental adaptations and damage to the brain. We aimed at investigating the effects of trekking and hypoxia on neuropsychological and neuroimaging measures.
Methods:
We recruited two balanced groups of healthy adults, trekkers (n = 12, 6 F and 6 M, trekking in altitude hypoxia) and controls (gender- and age-matched), who were tested before (baseline), during (5,000 m, after 9 days of trekking), and after the expedition for state anxiety, depression, verbal fluency, verbal short-term memory, and working memory. Personality and trait anxiety were also assessed at a baseline level. Neuroimaging measures of cerebral perfusion (arterial spin labeling), white-matter microstructural integrity (diffusion tensor imaging), and resting-state functional connectivity (functional magnetic resonance imaging) were assessed before and after the expedition in the group of trekkers.
Results:
At baseline, the trekkers showed lower trait anxiety (p = 0.003) and conscientiousness (p = 0.03) than the control group. State anxiety was lower in the trekkers throughout the study (p < 0.001), and state anxiety and depression decreased at the end of the study in both groups (p = 0.043 and p = 0.007, respectively). Verbal fluency increased at the end of the study in both groups (p < 0.001), whereas verbal short-term memory and working memory performance did not change. No significant differences between before and after the expedition were found for neuroimaging measures.
Conclusions:
We argue that the observed differences in the neuropsychological measures mainly reflect aspecific familiarity and learning effects due to the repeated execution of the same questionnaires and task. The present results thus suggest that detrimental effects on neuropsychological and neuroimaging measures do not necessarily occur as a consequence of short-term exposure to altitude hypoxia up to 5,000 m, especially in the absence of altitude sickness.
Introduction
High-altitude exposure is characterized by hypobaric hypoxia. Among a plethora of effects, it entails several physiological and neuropsychological adaptations (Clark et al., 1983; Caine and Watson, 2000; Virués-Ortega et al., 2004; Wilson et al., 2009; West, 2012; Prete et al., 2020; Bondi et al., 2021) that depend on the pattern and intensity of exposure: extending the exposure over multiple days causes an acclimation, which represents one of the most important components of the human capacity to adapt to high altitude (Da Porto et al., 2009).
Mood and anxiety
Several studies suggest that exposure to altitude hypoxia induces anxiety and mood state disturbances. Humans exposed to high altitude can exhibit symptoms such as breathlessness, palpitations, dizziness, headache, and insomnia, which are the same as those reported during panic attacks or severe anxiety states. Moreover, anxiety states and high-altitude exposure share at least three major clinical manifestations: hyperventilation, suffocation false alarms and cognitive misinterpretations.
Anxiety may or may not be related to acute mountain sickness (AMS). However, those who are more susceptible to AMS tend to show a higher level of anxiety (Missoum et al., 1992). Experimental studies conducted in the hypobaric chamber have shown that hypoxia exposure induces anxiety. Specifically, baseline levels of trait anxiety predict future severe AMS development at high altitude, whereas elevated levels of state anxiety are associated with AMS and its severity (Boos et al., 2018). However, other pieces of evidence disentangle the occurrence of anxiety and AMS, indicating that acute hypoxic physiological anxiety is an altitude-associated neurological disorder characterized by serious manifestations of anxiety and panic, without typical symptoms of AMS (Sracic et al., 2014).
The adverse effects of acute exposure to hypoxia on mood state are aggravated with the increment of altitude (Li et al., 2000). This effect is mediated by the emotional stability (i.e., neuroticism dimension) because individuals with low emotional stability are more sensitive to environmental stress than more stable individuals who face reality (Nicolas et al., 2000). The apparent discrepancy in the evidence may be solved by considering the difference in the level of motivation between volitional exposure to altitude in natural environments and the artificial confinement in hypobaric chambers. While the former triggers high levels of motivation, even under hardly demanding environmental stressors (Karinen and Tuomisto, 2017), hypoxia exacerbates the negative effect of forced confinement on mood (Stavrou et al., 2018).
Cognitive performance
High-altitude hypoxia has been shown to affect a broad spectrum of cognitive functions, with complex processes more susceptible to be markedly impaired. Cognitive impairments can persist several weeks after a high-altitude exposure (Cavaletti et al., 1990). The cognitive functions that are most likely to be affected by hypoxia are attention (Limmer and Platen, 2018), language (Griva et al., 2017), working memory (Yan et al., 2011a), learning (Muthuraju and Pati, 2014), decision-making, and also simple reactivity (Ray et al., 2019). A high degree of variability in the effect of hypoxia on cognitive performance has also been reported across subjects and studies (Griva et al., 2017).
A critical threshold for changes in cognitive responses to hypoxia has been proposed, corresponding to altitudes between 4,000 and 5,000 m (Nelson, 1982). Several authors have shown that acute exposure to altitudes of 4,000 m and above causes alteration in motor, memory, and perceptual tasks (Kennedy et al., 1989; Nelson et al., 1990; Bahrke and Shukitt-Hale, 1993; Virués-Ortega et al., 2004; Nardi et al., 2009). However, cognitive impairment in short-term verbal memory can already emerge at altitudes lower than 4,000 m (Bartholomew et al., 1999). Conversely, other studies demonstrated no significant cognitive impairment at 5,100 m (Harris et al., 2009) and even 6,200 m (Zhang et al., 2012).
Despite some ambiguity in the literature, it seems that short-term exposure with gradual ascents may not entail cognitive impairments below an extremely high threshold. Indeed, acclimation may drive the cognitive adaptations (Zhang et al., 2012; Merz et al., 2013).
Brain structure and function
Cognitive impairment at high altitudes may be due to inefficient neuronal functioning (Yan, 2014). Functional magnetic resonance imaging (fMRI) has demonstrated the detrimental impact of prolonged high-altitude exposure on the activity of the regions of the brain that are involved in verbal and spatial working memory (Yan et al., 2011a, 2011b). Using cerebral perfusion computed tomography, Bao et al. (2017) found a reduction of cerebral blood flow (CBF) in chronic mountain sickness patients, which was seen to be particularly low in the presence of cerebral edema. More recently, combining the use of MRI and diffusion tensor imaging (DTI), Bao's group reported multiple structural changes in chronic mountain sickness patients, such as a decrease of white matter in the right frontal lobe and damage to the left thalamus and the right hippocampus (Bao et al., 2019).
The high sensitivity of the hippocampus and limbic system to hypoxia may be the cause of the dysfunction in learning and memory (Hopkins et al., 1995; Virués-Ortega et al., 2004). Even in the absence of mountain sickness, other regions of the brain are structurally affected by high-altitude hypoxic environments (Hochachka et al., 1999), such as the parietal cortex (Di Paola et al., 2008) and the basal ganglia (Chen et al., 2017) after acute and chronic exposure, respectively. Further reported effects of an extremely high-altitude ascent (over 7,000 m) include cortical atrophy and periventricular hyperintensity lesions (Garrido et al., 1993).
In contrast, Anooshiravani et al. (1999) did not observe the presence of functional and structural, as well as neuropsychological, alterations in a group of climbers after returning from a very high-altitude (over 6,000 m) mountain expedition. When using DTI, however, a number of microstructural compromissions have been detected: a diffused reduction of fiber integrity (corticospinal tracts, corpus callosum, reticular formation of dorsal midbrain, left superior longitudinal fasciculus, right posterior cingulum bundles, and left middle cerebellar peduncle) after a very high-altitude climbing (6,200 m) (Zhang et al., 2012) as well as an increased path length of callosal fibers connecting homotopic visual areas after long-term (2 years) high-altitude exposure (Chen et al., 2016b). Widespread changes in diffusivity measures were also observed in college students after 30 days at 4,300 m together with changes in gray and white matter volume, all reversing after 2 months (Fan et al., 2016).
These impairments in cerebral white matter after very high or long-term exposure may explain some of the previously found cognitive and motor deficits.
Few studies also used resting-state fMRI (Fox and Raichle, 2007) to evaluate the effects of high-altitude exposure on the brain intrinsic (resting state) functional connectivity (FC). Neuroimaging studies have indeed shown that resting-state FC (rs-FC) can be used to identify large-scale resting-state networks that exhibit covariance structures at ultraslow frequencies, and whose topography strongly recapitulates the one observed during active behavior in different domains (e.g., Thomas Yeo et al., 2011). It has been reported that rs-FC can be quickly modified by training in a behaviorally relevant manner (Lewis et al., 2009), thus making it a suitable marker for brain adaptation to high-altitude and extreme environments.
Previous studies focused on long-term (2 years), high-altitude exposure in groups of adults (Chen et al., 2016a, 2016b; Zhang et al., 2017) as well as college freshmen (Chen et al., 2017) who had immigrated to Tibet, finding diffuse changes that correlated with the cognitive loss in working memory and psychomotor function in students (Chen et al., 2017). These were interpreted as central neural compensatory mechanisms underlying respiratory, visual, and cognitive dysfunctions in adults (Chen et al., 2016a, 2016b; Zhang et al., 2017).
Aim of the present study
To our knowledge, no study has examined brain function impairment due to high-altitude hypoxia by conducting a comprehensive neuropsychological and neuroimaging assessment, including structural and FC measures, in a group of both men and women, while also controlling for possible unspecific effects due to the repetition of the tasks and measuring. Therefore, the main aim of the present study was to investigate the selective effects of high-altitude trekking on mood, cognition, and brain structure and function in participants taking part in the “Trek Gokio Kumbu/Amadablam 2012” project. Specifically, we followed an ecological approach to isolate the effect of a long-lasting trekking at hypobaric hypoxia on anxiety, mood, verbal fluency, verbal short-term memory, and working memory, as well as on brain microstructure, CBF, and intrinsic FC.
Materials and Methods
Participants
The experimental group (trekkers) was composed of 12 participants (6 females, 42.1 ± 9.8 years of age, 14.8 ± 3.5 years of schooling), who voluntarily participated in the scientific expedition “Trek Gokio Kumbu/Amadablam 2012”. The control group matched the trekker group, as composed by 12 participants (6 females, 36.6 ± 6.2 years of age, 16.7 ± 1.2 years of schooling). All participants provided written informed consent before participating in the study. They were all in good health, as reported by a medical check. This research was approved by the local Human Ethics Committee (protocol no. 773 COET). No participants suffered from AMS.
Experimental design
The altitude expedition took place over a period of 20 days between November and December 2012. In the trekkers, the neuropsychological battery was administered at three time points: before (Baseline-Chieti), during (Base Camp at 5,000 m), and after (Post-Chieti) the high-altitude trekking. Several results of such expedition have already been published (Mancinelli et al., 2016; Morabito et al., 2016; Tam et al., 2016; Verratti et al., 2016, 2017; Doria et al., 2020). The Big Five Questionnaire (BFQ) and the Trait Anxiety Inventory were only administered at baseline levels. After the expedition, participants of the control group were enrolled and underwent the same neuropsychological evaluations with approximately the same time intervals as the trekkers (Baseline, T1 = During, T2 = Post). The neuroimaging measures were only carried out on the trekkers before and after the expedition.
Trekking plan
The altitude trek lasted 14 days, with 12 days of trekking. Two days were spent at rest in altitude: one for acclimation in Namche Bazar (3,440 m) (day 3 of expedition) and one at the Italian Pyramid (5,050 m) (day 11 of expedition) for experiments. The altimetric profile of the trek is shown in Figure 1. The total distance covered walking was ∼122 km. The total difference in altitude of ascent and descent was 6,953 m (569 m × d−1; range: 86–1,875 m × d−1) and 6,040 m (503 m × d−1; range: 37–1,193 m × d−1), respectively. The walking time was ∼6 hours per day, and the average speed was 0.47 m × s−1. The total number of steps exerted by the participants was 181,319 ± 18,440.
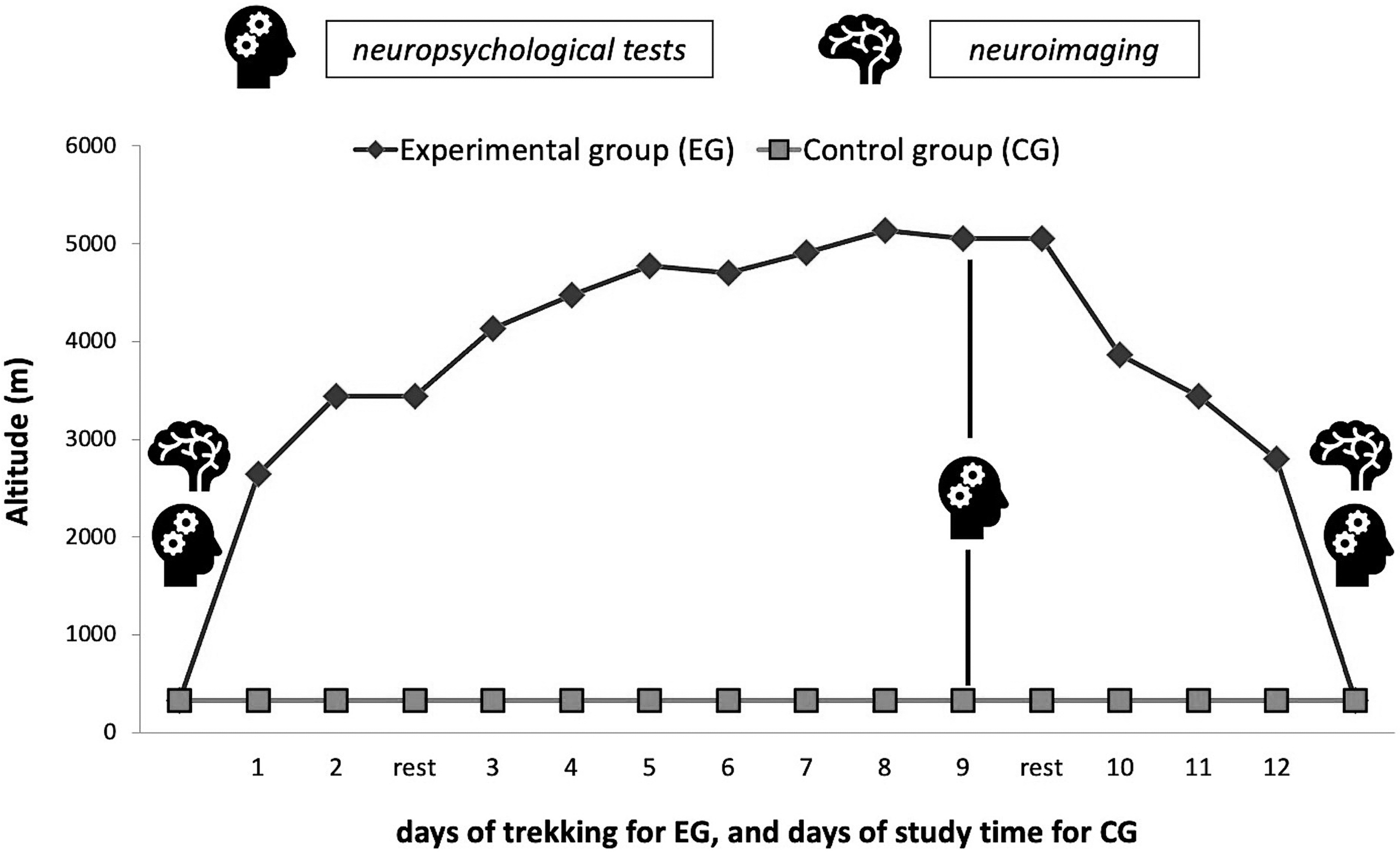
Study design of the expedition. Neuropsychological tests and neuroimaging acquisition before and after the expedition of trekkers, as well as all the neuropsychological tests of the control group, were conducted in Chieti (330 m).
Procedures
Neuropsychological assessment
We administered the following written, verbal, and computerized tests:
BFQ-2 (Caprara et al., 1993; Costa and McCrae, 1995): the test measures five major dimensions of personality, that is, openness, conscientiousness, agreeableness, extraversion, and neuroticism. STAI1-STAI2—State Trait Anxiety Inventory (Spielberger et al., 1970): the inventory measures changes in the amount of state (STAI-1) and trait (STAI-2) anxiety. In STAI-1, the subject must choose one of four alternatives that best reflect “how you feel at this moment”: (1) not at all, (2) a little bit, (3) enough, (4) very much. In STAI-2, the subject must indicate which of the four options “describes how you usually feel”: (1) almost never, (2) sometimes, (3) often, (4) almost always. BDI—Beck Depression Inventory (Beck and Steer, 1993): the inventory quantifies constructs such as depression and dysphoria, asking participants to choose, for each of the twenty items, the alternative that comes closest to how they have been feeling in the last week. Verbal Fluency test: this test evaluates the amplitude of the warehouse vocabulary, the ability to access the lexicon, and the lexical organization. Participants were asked to provide as many words as possible beginning with a given letter within a minute, excluding proper nouns and numbers. We used three sets of three letters across the three sessions: F-A-S (Carlesimo et al., 1995; Zappalà et al., 1995), F-P-L (Novelli et al., 1986; Capitani et al., 1998), and C-P-S (Mondini et al., 2003). Digit Span Forward test (Orsini et al., 1987): this test measures the verbal short-term memory capacity (i.e., span) by asking participants to serially recall a list of numbers previously read by the experimenter. The score corresponds to the length of the longest string of digits correctly reported. 2-Back task (Kirchner, 1958): this task measures the continuous visual working memory performance. Participants monitored the recursive presence of random numbers (0 to 9) on a computer screen and indicated when the current stimulus matched the one presented 2 steps earlier. Each number was presented in gray at the center of a black screen for 1 second and was followed by a 500-millisecond interstimulus interval. Participants had 1 second from the onset of the stimulus to indicate stimulus repetition using the space bar of the computer keyboard. Each session included the presentation of two lists, each comprising 100 stimuli, separated by a break. Different lists of trials were administered across the three sessions. Overall response accuracy was used as the dependent variable.
Neuroimaging measures
DTI enables the measurement of the random movement of water molecules in brain tissues in vivo (Pierpaoli et al., 1996), allowing to detect microstructural changes in white matter before macrostructural changes are apparent. Fractional anisotropy (FA) is a commonly DTI-derived measure used to quantify the degree of anisotropic diffusion within the single voxel; higher FA values are thought to reflect better white matter integrity as a result of greater intravoxel coherence of fiber orientation, axon density and diameter, and/or myelination (Beaulieu et al., 1996; Sen and Basser, 2005; Smith et al., 2007; Caminiti et al., 2013).
n addition to FA, mean diffusivity (MD) reflects the overall magnitude of water diffusion in each measured voxel and it is particularly sensitive to the extracellular volume (Sotak, 2002). In general, low MD reflects a high density of cells and/or extracellular material that impedes the diffusion of water through brain tissue (Mackey et al., 2012). Combined studies of FA and MD offer a potential new field for exploring temporal and spatial aspects of brain changes (O'Dwyer et al., 2011).
DTI data were acquired using a 3T Philips Achieva scanner (Philips Medical Systems, Best, Netherlands) at ITAB—Institute for Advanced Biomedical Technology, Chieti, Italy, with the following parameters: 32 diffusion encoding directions, slice thickness = 2 mm, Repetition Time (TR) = 6,563 ms, Echo Time (TE) = 70 ms, flip angle = 90°, matrix = 112 × 110, Field of View (FOV) = 224 × 120 mm2, b = 0 and 800 s/mm2, 60 axial slices, and voxel size = 1.75 × 1.75 × 2.0 mm3. A 2D fluid-attenuated inversion recovery T2-weighted scan was also used to exclude the presence of small-vessel ischemic disease and other supra- or infratentorial brain lesions.
Arterial spin labeling (ASL) is a noninvasive fMRI technique that uses arterial water as an endogenous intravascular tracer to measure CBF with higher spatial and temporal resolution than other techniques. CBF is defined as the rate of delivery of arterial blood to the capillary beds of a particular mass of tissue. A pulsed ASL sequence with echo-planar imaging and signal targeting with alternating radio frequency was used. The readout was performed with T2*-weighted, single-shot, echo-planar imaging using the following parameters: TR/TE = 4,000 ms/20 ms, 30 tag-control pairs consisting, 19 slices with 3.5 × 3.5 × 6 mm3 resolution, SENSE factor 2.3, label thickness = 130 mm, label gap = 20 mm, label delay = 1,200 ms.
rs-FC by MRI is a noninvasive fMRI technique that investigates the spontaneous intrinsic fluctuations of the blood oxygen-level-dependent (BOLD) signal recorded in the absence of task (Fox and Raichle, 2007). We collected structural and functional MRI images on a Philips Achieva 3T scanner. Structural scans consisted of the following: (1) a sagittal M-PRAGE T1-weighted sequence (TR = 8.14 ms, TE = 3.7 ms, flip angle = 8°, voxel size = 1 × 1 × 1 mm) and (2) a T2-weighted sequence (TR = 3 seconds, TE = 80 ms, flip angle = 90°, voxel size = 0.98 × 1 × 1 mm, 39 slices). Resting-state functional scans were acquired with a gradient echo EPI sequence (TR = 1,914 ms, TE = 25 ms, 39 slices acquired in ascending interleaved order, voxel size = 3.59 × 3.59 × 3.59 mm, 64 × 64 matrix, flip angle = 80°), during which participants were instructed to keep their eyes open in a low luminance environment.
Data analysis
Neuropsychological data
Statistical analyses and plots were carried out using R-based open-source software Jamovi Version 1.2.27.0 (retrieved from https://www.jamovi.org). For the baseline comparisons of the BFQ and the Trait Anxiety Inventory, we first checked the assumption through the Shapiro–Wilk test of normality of distributions, the Levene's test of equality of variances, the Shapiro–Wilk test of normality of residuals, and Q-Q plots. Consequently, we used the independent-sample t-test, with both the Student's and the Bayesian method (Cauchy's distribution with λ = 0.707 was used arbitrarily for a priori distributions, and Bayes factors robustness check was performed). Considering the small sample size, Cohen's d was adjusted to Cohen's dunbiased (Fritz et al., 2012) as follows:
For the comparisons of the State Anxiety Inventory, BDI, Digit Span Forward test, Verbal Fluency Test, and 2-back test, we used a repeated-measures approach. We first checked the normality of distributions with the Shapiro–Wilk test, and then we used the Levene's test of equality of variances and the Mauchly's test of sphericity. After the assumption checks, we used alternatively the ANOVA for repeated-measures test or mixed-model analysis, with the group (trekkers vs. control) as the between factor. Specifically, we used ANOVA for repeated measures in case of balanced designs, and mixed-model analysis in case of missing data (Armstrong, 2017).
Regarding ANOVA for repeated measures, if the sphericity assumption was violated, a correction was applied following the typical recommendation of using the Greenhouse–Geisser method in case of estimated ɛ < 0.75 or the Huynh-Feldt method in case of estimated ɛ > 0.75; post hoc tests were conducted with Tukey correction for multiple comparisons. To further determine whether verbal fluency results were affected by other known variables, we additionally run the analysis with age as a covariate. Regarding mixed-model analysis, participants were considered the random factor and the restricted maximum likelihood estimation method was used; post hoc tests were conducted with Bonferroni correction for multiple comparisons. Partial eta-squared (η2p) and partial omega-squared (ω2p) were used as a measure of effect size (Fritz et al., 2012).
Images were created with the JJStatsPlot module implemented in Jamovi (Patil, 2021; Wickham et al., 2018).
Neuroimaging data
DTI data were preprocessed using different tools from the FMRIB's Diffusion Toolbox, part of FSL (FMRIB's Software Library; www.fmrib.ox.ac.uk/fsl; Smith et al., 2004). All raw diffusion volumes were first corrected for eddy current distortion and head motion using a 12-parameter affine registration to the first no-diffusion weighted volume of each subject, and the gradient directions were rotated accordingly (Leemans and Jones, 2009). Nonbrain tissue was removed from the eddy-corrected images using the Brain Extraction Tool (Smith, 2002) creating a binary mask of the brain. Then, FA and MD scalar maps were estimated at the individual level using the FSL diffusion tensor fitting program tool by fitting a tensor model to the eddy-corrected and brain masked diffusion data. FA and MD data from each participant were further analyzed using the Tract-Based Spatial Statistics (TBSS) (Smith et al., 2006) toolbox available in FSL.
All participants' FA data were aligned into a common space using the nonlinear registration tool FNIRT (Andersson et al., 2007a, 2007b), which uses a b-spline representation of the registration warp field (Rueckert et al., 1999). Next, the mean FA image was calculated and thinned to create a mean FA skeleton, as the centers of all tracts common to the group. Each subject's aligned FA data were then projected onto this skeleton and the resulting data fed into a longitudinal voxelwise statistical analysis (paired t-test). The same procedure was applied to the calculated MD maps, using the individual registration and projection vectors obtained in the FA nonlinear registration and skeletonization stages with TBSS_non_FA tool.
Voxelwise statistical analyses were carried out using permutation-based nonparametric statistics using the FSL Randomize permutation-based program (Nichols and Holmes, 2002) with 10,000 permutations. The statistical threshold was established with a familywise error-corrected p-value (pFWE) <0.05 with multiple comparison correction using threshold-free cluster enhancement (Smith and Nichols, 2009).
Analysis of ASL data was performed using the software Analysis of Functional NeuroImages with a separate preprocessing of tag and control images (Wang et al., 2008; Chiacchiaretta et al., 2018) as follows: (1) motion correction was performed to realign all tag/control time frames to a reference image represented by the first tag/control volume; (2) all the preprocessed images were coregistered to the structural scan; and (3) individual images were warped into the Talairach space for group analysis. Finally, regional CBF maps were obtained from the signal difference between control and tag images as in Tortora et al. (2017), and changes in CBF values after the expedition were evaluated using a paired t-test.
Resting-state functional MRI data were preprocessed by applying a pipeline described in a previous work (Sestieri et al., 2014) by using in-house software functional Interactive Data Language developed at Washington University (St. Louis). Preprocessing of functional BOLD images included the following: motion-correction within and between runs, correction for cross-slice timing differences, resampling into 3 mm isotropic voxels, and warping into 711–2C space, a standardized atlas space (Talairach and Tournoux, 1988; Van Essen, 2005) as well as whole-brain normalization correcting for changes in overall image intensity between BOLD runs.
Furthermore, in preparation for the FC analysis, data were passed through several additional preprocessing steps as follows (Fox et al., 2009): isotropic spatial smoothing 6-mm full-width at half-maximum Gaussian blur in each direction; temporal filtering retaining frequencies below 0.1 Hz; and removal of the following sources of spurious variance through linear regression: (1) head motion (modeled through a six-parameter affine transformation), (2) global brain signal (estimated as the average signal from fixed region in atlas space), (3) signal from a ventricular region of interest, and (4) signal from a region centered in the white matter.
Next, rs-FC was estimated for each subject at each time point (baseline and T2) by computing the Pearson correlation coefficient (r) among fMRI signals extracted by a set of 60 regions of interest (ROI) comprising five networks: default mode (Fox et al., 2005), phonological and semantic language (Vigneau et al., 2006), motor (Hacker et al., 2013), and working memory (Owen et al., 2005) networks. This procedure yielded for each subject an FC matrix in which each cell indicates the strength of FC between each pair of ROI at each time point. Changes in FC magnitude were assessed by means of paired t-test on the FC matrices on each time point.
Results
Neuropsychological data
Baseline: BFQ and STAI-T
Table 1 shows baseline results for both groups. Among the BFQ dimensions, we found a significant difference in conscientiousness, where trekkers scored lower than controls (p = 0.033, dunb = 0.894, BF10 = 2.173). We also found a significant difference in Trait Anxiety, where trekkers scored lower than controls (p = 0.003, dunb = 1.298, BF10 = 11.841).
Comparison of Big Five Questionnaire Dimensions (Openness, Conscientiousness, Agreeableness, Extraversion, and Neuroticism) and Trait Anxiety Inventory Between the Experimental and Control Groups
BF, Bayes factor; C.I., confidence interval; mean diff., mean difference; SE diff., standard errors of differences; dunb, unbiased Cohen's d.
Repeated measures: STAI-S, BDI, Verbal Fluency, Digit Span Forward, 2-back task
Table 2 reports data from the three time points in both groups. STAI results showed a significant effect of group (trekkers lower than controls; F1,20 = 25.393, p < 0.001, η2p = 0.559, ω2p = 0.526) and time (F2,40 = 3.384, p = 0.043, η2p = 0.145, ω2p = 0.100) in State Anxiety (Fig. 2). Post hoc analysis revealed tendencies for a significant reduction of state anxiety at Post relative to both Baseline (pBonferroni = 0.081, mean diff. = 4.000, SE diff. = 1.748) and During (pBonferroni = 0.096, mean diff. = 3.875, SE diff. = 1.748). No other differences or tendencies were observed.
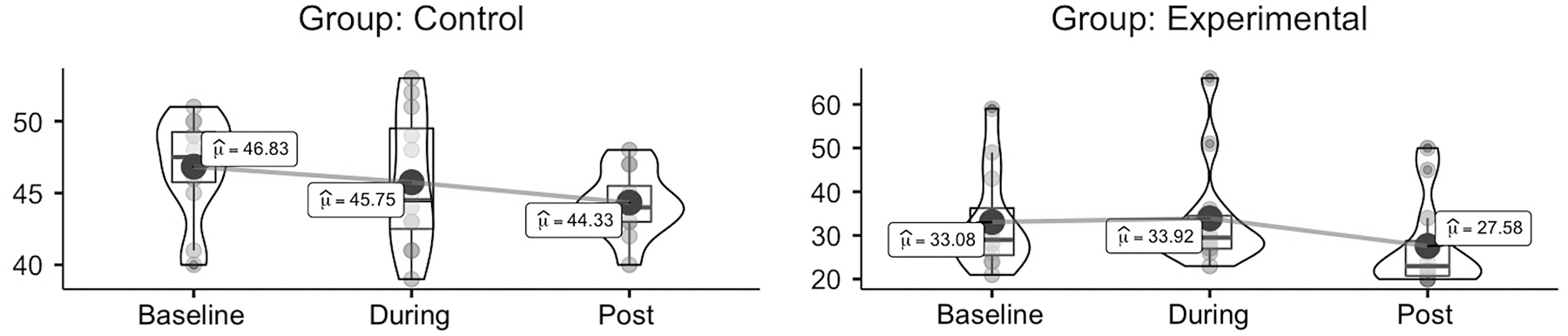
Violin plots of STAI-State Anxiety results, grouped by time (at baseline vs. during vs. after the study period) and split by group (trekkers vs. controls); box plots represent median and interquartile range, dots represent individual values, and big dots represent mean. STAI, State Trait Anxiety Inventory.
Results of the Neuropsychological Tests Across the Three Time Points
BDI, Beck Depression Inventory; STAI-S, State Trait Anxiety Inventory-State.
BDI results showed a significant effect of time (F2,35 = 5.783, p = 0.007, η2p = 0.248, ω2p = 0.201). Post hoc analysis revealed a significant reduction from During to Post (pBonferroni = 0.005, mean diff. = 2.172, SE diff. = 0.639). No other differences or tendencies were observed.
Verbal Fluency results showed significant effect of time (F2,40 = 9.779, p < 0.001, η2p = 0.328, ω2p = 0.290). Post hoc analysis revealed a significant increment of correct responses [ncr] from Baseline to Post (p < 0.001, mean diff. = 7.612 ncr, SE diff. = 1.734 ncr) and, with a lesser extent, from During to Post (p = 0.029, mean diff. = 4.604 ncr, SE diff. = 1.734 ncr). A tendency for a significant time by group interaction was also observed (F2,40 = 3.496, p = 0.063, η2p = 0.149, ω2p = 0.104) (Fig. 3).

Violin plots of Verbal Fluency test results, grouped by time (at baseline vs. during vs. after the study period) and split by group (trekkers vs. controls); box plots represent median and interquartile range, dots represent individual values, and big dots represent mean.
No significant effect was observed regarding the accuracy of the 2-back task and the Digit Span Forward state.
Neuroimaging data
The TBSS analyses did not yield significant differences in FA and MD maps across time points. To verify the results of TBSS, we have also investigated longitudinal FA changes in white matter regions, which showed microstructural changes after an ecological hypoxia exposure in a previous study (Zhang et al., 2012). The following regions of interest (ROIs) were taken from the JHU ICBM-DTI-81 White-Matter Labels atlas included in the FSL distribution: corpus callosum, middle cerebellar peduncle, right and left cingulum, corticospinal tracts, and superior longitudinal fasciculi. For each ROI, mean FA values (from nonzero voxels) were obtained from the skeletonized FA maps, and longitudinal changes were estimated using Wilcoxon signed-rank test (p < 0.05 for null hypothesis rejection). No significant difference was detected in any of the ROIs (see Supplementary Fig. S1).
Similarly, we did not find any significant difference in the whole-brain CBF from the ASL analysis (Fig. 4). Finally, rs-FC analyses did not reveal any significant differences in the strength of rs-FC among the set of ROIs between baseline and post-expedition (see Supplementary Fig. S2).

CBF before and after the altitude expedition in the experimental group, assessed through fMRI with arterial spin labeling technique. CBF, cerebral blood flow; fMRI, functional magnetic resonance imaging.
Discussion
The growing interest in expeditions at high altitudes has meant that an increased number of people have been exposed to the risk of brain structural and functional damage (Caine and Watson, 2000). However, a comprehensive interpretation of the risk of damage to cognitive functions in such an environment remains to be elucidated. Among the topical studies, there have been numerous studies concerning attentional performance and memory skill. The interest in the latest domain is most likely because memory is the first cognitive function to be affected by a lack of oxygen. This is what is found in neurodegenerative diseases, such as Parkinson's and Alzheimer's, and as a substrate of age-dependent physiological pathways: indeed, a hypoxia-dependent pathway of aging processes has been proposed (Di Giulio et al., 2009; Verratti et al., 2009).
Moreover, ecological studies at high altitudes provide an interesting model to study the consequences of cerebral hypoxia in healthy individuals (Wilson et al., 2009). Therefore, the primary target of this study was to investigate the effects of hypoxia on the brain, using neuropsychological assessment and neuroimaging measures.
At a baseline level, the trekker group showed lower values of conscientiousness, state and trait anxiety compared with the control group. Therefore, the trekkers were more prone to face the stressors, and their normal level of emotional stability/neuroticism is also in agreement with this (Nicolas et al., 2000). Notably, state anxiety remained significantly lower in the trekkers compared with the control group throughout the study. The low baseline levels of trait anxiety as well as of state anxiety during the expedition might have contributed to the lack of AMS development at high altitude (Missoum et al., 1992; Boos et al., 2018). On the contrary, the volitional exposure and the succesfull acclimation must have contributed to the generally low levels of both anxiety and depression in the trekkers, which were particularly evident at the end of the expedition.
Our participants demonstrated no significant memory impairment at high altitude (5,050m). Comparing this with previous field studies, Forster (1985) assessed two groups, at sea level and at 4,200 m, detecting the absence of memory deficits in the high-altitude group. In climbers, 75 days after returning from extremely high-altitude exposure without supplementary oxygen, a still significant impairment in associative memory, but not in pure memory, was found (Cavaletti et al., 1990). As suggested by a previous study, the memory load may be the factor driving the memory deficit, since a detrimental effect was only observed in the high-load conditions (Bartholomew et al., 1999). In the abovementioned study, Cavaletti's group also reported a trend toward a significant increase of performance at the Verbal Fluency test, likely due to a learning effect (Cavaletti et al., 1990). In relation to this, one of the main strengths of the current study was the presence of a control group, which performed the same cognitive tasks at equal time points but without hypoxic exposure (see also Fan et al., 2016), thereby allowing us to unmask the effects of learning and task version.
Considering the occurrence of an increased performance both in the controls and trekkers, we argue that such improvement was due to a learning effect. The tendency to significance of the time by group interaction suggests a possible mitigation of the learning effect due to high-altitude exposure. Indeed, while the increment in the number of words was evident already at the second time point for the control group, it was only evident at the third point, thus after the expedition, in the trekkers.
No changes were detected in the accuracy of the working memory task. Therefore, such a cognitive task was not affected by altitude exposure, nor a learning effect was demonstrated. This last finding was already suggested by Jacola et al. (2014), who argued that the testing context does not affect the n-back task in terms of accuracy.
On the contrary, Petiet et al. (1988) demonstrated that simple cognitive skills remain intact after hypoxic exposure, as well as in aging, whereas complex cognitive skills (i.e., abstract reasoning and word finding) are more vulnerable. We partially agree with them, arguing that high-altitude exposure does not entail detrimental effects in short-term memory, updating skills, and phonemic fluency if altitude sickness does not occur. The only detectable consequence was a mitigation of the learning effect for the fluency task. The co-occurrence of altitude sickness and the evaluation of sleep-adaptation to high altitude (Bloch et al., 2015) might help the understanding of the discussed phenomena.
Regarding the neuroimaging data, no change was detected in either structural (white matter integrity) or functional (cerebral perfusion and intrinsic FC) measures.
To our knowledge, the present study is one of the few examinations of the white matter microstructural changes via DTI after an ecological hypoxia exposure in the absence of clinical conditions. Our results are in line with those of a previous voxel-based morphometry study that showed no significant differences in white matter volume in a group of world-class mountain climbers after a single extremely high-altitude expedition (Di Paola et al., 2008). Instead, previous DTI studies detected microstructural compromissions of white matter fibers (Zhang et al., 2012; Chen et al., 2016b; Fan et al., 2016). However, they were conducted after a very high altitude or a longer exposure, thus differing from the experimental conditions of our study (i.e., short-term exposure at high altitude).
Recently, different groups reported a lowering of CBF and/or white matter in patients with chronic mountain sickness and/or high-altitude cerebral edema (Bao et al., 2017, 2019; Hackett et al., 2012; Turner et al., 2021). Interactions between hypoxic mechanism, inflammation, and white matter integrity have been proposed (Chen et al., 2015); moreover, the specific time course of adaptations may require at least 3–4 weeks (for reliable positive changes in FA and MD maps, see Mackey et al., 2012 and Scholz et al., 2009). Therefore, we argue that the short-term hypoxic exposure, in the asbsence of altitude symptoms, did not entail detrimental changes.
Overcoming the absence of adaptation in CBF, we may evoke the challenge posed by altitude exposure on cerebrovascular reactivity, due to the opposed effects of hypoxia-related vasodilation and hypocapnia-related vasoconstriction. Further studies may focus on regional differences in CBF regulation, as demonstrated by Lafave et al. (2019). For what concerns the brain FC at rest, the present data show that short-term exposure does not significantly affect the intrinsic functional architecture of the human brain, unlike previous data available on immigrants after long-term/chronic exposure to high altitude (Chen et al., 2016a, 2016b, 2017; Zhang et al., 2017).
Before concluding, it is worth underlying that the present ecological study, unlike most of previous ones (see Petiet et al., 1988; Fan et al., 2016), included also women in the two groups of participants, and was not designed to unveil mechanistic evidence about either the existence or the nature of causal mechanisms connecting anxiety, mood, cognitive functions, and neuroimaging measures. Beyond the limited sample size, its main limitations rely on the fact that the trekkers performed the second scans 5 days after beginning the descent and the control group did not perform an equivalent trek at sea level, thus not being subjected to the same volume of mechanical load and environmental factors.
To our justification, these lacks were, respectively, due to the time strictly necessary to return home from the expedition and to the extreme difficulty of reproducing environmental factors such as temperature and humidity, considering the high variability of these parameters typical of the ecological study.
Conclusions
High-altitude exposure is an important model for the ecological psychophysiological study of adaptation to extreme environmental stressors such as hypoxia, a very definite phenomenon for its involvement also in neurodegenerative disorders. The present neuropsychological and neuroimaging data extend the topic of neurocognitive correlates of altitude hypoxia by showing that, in the absence of AMS, an altitude trek at 5,000 m does not entail pathological adaptations of brain structure and function, including the intrinsic functional architecture studied for the first time after a nonchronic exposure to high altitude.
With the inclusion of a control group, our ecological study design allowed us to unmask specific learning/familiarity effects, revealing a possible mitigation by hypoxia in the high-load fluency task, which itself deserves further investigation. In a similar vein, further studies implementing longer periods of hypoxic exposure for larger samples of trekkers (both males and females) are required to confirm the present data and better understand the role of gender and exposure duration/elevation beyond that of mountain sickness on the psychophysiological adapatations to high altitude.
Footnotes
Acknowledgments
The authors are grateful to all the Nepalese porters and Sherpas for their essential role in the scientific expedition; brotherly thanks go to the Himalayan guide, Ram-chandra Sapkota. Special thanks to both the researchers and experimental subjects who volunteered to take part in this study.
Authors' Contributions
Conceptualization: G.C. and V.V.; methodology: G.C., G.D.M., C.S., R.R., and V.V.; formal analysis: G.C., D.B., C.S., G.D.M., C.P., C.D., R.R., A.B., T.P., R.S., R.N., P.C., A.F., and V.V.; investigation: G.D.M., C.D., R.R., R.S., and V.V.; resources: G.C., C.S., T.P., and V.V.; data curation: G.C., C.S., and V.V.; writing—original draft preparation: G.C., D.B., C.P., and V.V.; writing—review and editing: G.C., D.B., C.S., G.D.M., C.P., C.D., R.R., A.B., T.P., R.S., R.N., P.C., A.F., and V.V.; visualization: D.B. and V.V.; supervision: G.C. and V.V.; project administration: V.V.; funding acquisition: G.C. and V.V. All authors have read, reviewed, and agreed to the published version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by departmental funds to VV.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.