
Abstract
Doherty, Connor J., Jou-Chung Chang, Benjamin P. Thompson, Erik R. Swenson, Glen E. Foster, and Paolo B. Dominelli. The impact of acetazolamide and methazolamide on exercise performance in normoxia and hypoxia. High Alt Med Biol. 24:7–18, 2023.—Carbonic anhydrase (CA) inhibitors are commonly prescribed for acute mountain sickness (AMS). In this review, we sought to examine how two CA inhibitors, acetazolamide (AZ) and methazolamide (MZ), affect exercise performance in normoxia and hypoxia. First, we briefly describe the role of CA inhibition in facilitating the increase in ventilation and arterial oxygenation in preventing and treating AMS. Next, we detail how AZ affects exercise performance in normoxia and hypoxia and this is followed by a discussion on MZ. We emphasize that the overarching focus of the review is how the two drugs potentially affect exercise performance, rather than their ability to prevent/treat AMS per se, their interrelationship will be discussed. Overall, we suggest that AZ hinders exercise performance in normoxia, but may be beneficial in hypoxia. Based upon head-to-head studies of AZ and MZ in humans on diaphragmatic and locomotor strength in normoxia, MZ may be a better CA inhibitor when exercise performance is crucial at high altitude.
Introduction
As mountain tourism increases in popularity over the years, acute mountain sickness (AMS) among high-altitude tourists will increase, often due to rapid ascent and the lack of acclimatization to altitude. Acetazolamide (AZ) is a common prophylactic medication used to reduce the occurrence and severity of AMS by augmenting the hypoxic ventilatory response (HVR) and facilitating acclimatization to altitude. While using AZ for prophylaxis of AMS is unquestionably beneficial (Bärtsch and Swenson, 2013), it may have negative consequences on exercise performance (Dominelli et al, 2018; Garske et al, 2003; Stager et al, 1990). In addition, the drug's impact on exercise performance varies with dosing and altitude. A recent review found that AZ reduced exercise performance at sea level, but the impact of AZ on exercise performance in hypoxia remained unclear (Posch et al, 2018).
Methazolamide (MZ), an alternative carbonic anhydrase (CA) inhibitor to AZ, is also efficacious in treating AMS but may not hinder exercise performance to a similar degree. However, there is a lack of prospective studies comparing these two CA inhibitors head-to-head with respect to exercise performance at high-altitude, making definitive conclusions difficult. The purpose of this review is to compare the effects of AZ and MZ on exercise performance at sea level (i.e., normoxia) and high altitude (i.e., hypoxia). We focus on exercise performance outcomes (e.g., time to complete a set distance) and physiological variables directly related to performance (e.g., maximal oxygen uptake [V̇O2max]) along with the physiological mechanisms by which AZ and MZ may affect exercise performance.
Hypoxic Ventilatory Response
Before detailing AZ or MZ effects on exercise performance, it is informative to describe why an individual would take a CA inhibitor to prevent or treat AMS. The complete description of AMS and/or the impact of hypoxia itself on exercise is beyond the scope of this review and is extensively covered elsewhere (Bärtsch and Swenson, 2013; Luks and Hopkins, 2014). As such, we will provide a brief overview focusing on the physiological challenges of hypoxia and its effects on exercise performance.
Without an adequate acclimatization period, AMS can develop following rapid ascent to high altitude (e.g., >2,500 m). At high altitudes, the lower barometric pressures result in a lower inspired partial pressure of oxygen (PIO2), which leads to decreased alveolar oxygen tension, arterial oxygenation (PaO2), and a lower arterial oxyhemoglobin saturation (SPO2) and arterial oxygen content (CaO2) (Bärtsch and Swenson, 2013; Swenson, 2016). The reduction in CaO2 is ultimately what hinders exercise performance with decrements in V̇O2max observed at altitudes as low as 580 m above sea level (Gore et al, 1996).
One of the first physiological reflex mechanisms triggered at high altitude to defend against reductions in PaO2 and tissue oxygenation is increasing ventilation (V̇E) (i.e., HVR). In response to arterial hypoxemia, the chemically sensitive carotid bodies detect a drop in PaO2 and relay afferent feedback to the respiratory centers to increase alveolar ventilation (Teppema and Dahan, 2010). The increased V̇E improves PaO2 but lowers arterial carbon dioxide tension (PaCO2). Carbon dioxide plays an important role in blood pH through the reversible reaction, rapidly catalyzed by CA, that converts water and CO2 to bicarbonate (HCO3−) and hydrogen ions (H+) in a reversible manner [Eq. (1) below].
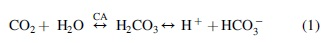
As a consequence of decreased PaCO2, blood pH increases and blunts the initially higher ventilatory drive, secondary to the reduced stimulation of central and peripheral chemoreceptors. This decrease in V̇E occurs 5–20 minutes after exposure to hypoxia and attenuates the initial rise in PaO2 with HVR. Following chronic exposure to high altitude, respiratory alkalosis, and its suppression of ventilation is partially corrected by renal bicarbonate excretion, among other physiological adaptations to altitude. Increased renal bicarbonate excretion increases H+ concentration in the blood and tissues, which improves central and peripheral chemoreceptor stimulation of ventilation and increases PaO2 and CaO2. However, a greater ventilation will further reduce PaCO2, limiting the role of central chemoreceptors in mediating the increases in ventilation.
Pharmacological Inhibition of CA
CA isoenzymes are found throughout the body and are responsible for catalyzing the reversible hydration of CO2 and dehydration of bicarbonate [Eq. (1)]. In blood, bicarbonate acts as a buffer to help maintain systemic pH (Davenport, 1974). In the lungs, CA catalyzes the reverse reaction of Equation (1), dehydrating bicarbonate into water and CO2 with the latter expired with ventilation (Klocke, 1978). At the kidneys, CA mediates the reabsorption of bicarbonate by converting it to CO2 and water for diffusion into renal tubular cells, then converting it back to bicarbonate for export into renal venous blood (Maren, 1967). In healthy individuals at sea level, bicarbonate is entirely reabsorbed and returned to the circulation to maintain blood pH homeostasis (Davenport, 1974), unless an alkalemic state exists.
At high altitude, the elevated blood pH caused by HVR is corrected by increasing renal bicarbonate excretion over several days. Prophylactic treatment with a CA inhibitor hastens this correction of blood pH to altitude by immediately accelerating bicarbonate excretion. Inhibiting renal bicarbonate reabsorption produces a mild metabolic acidosis, which counteracts the respiratory alkalosis caused by HVR (Davenport, 1974; Swenson, 2016). The return of blood pH to approximately prehypoxic exposure levels almost restores hypoxic ventilatory drive, increases V̇E and improves alveolar and arterial oxygenation (Leaf and Goldfarb, 2007; Swenson, 2014). The improved arterial oxygenation with pharmacological CA inhibition is considered the primary mechanism in reducing AMS (Richalet et al, 2012; Swenson, 2014).
While CA inhibition is efficacious in preventing/treating AMS, in a small study of 13 subjects, it did not prevent to a statistically significant extent other high-altitude illness such as pulmonary edema when susceptible climbers ascended to 4,559 m (Berger et al, 2022). However, further research is required to determine whether CA inhibition truly reduces the severity or incidence of developing pulmonary edema.
Differences in CA Inhibitor Dosing
The pharmacodynamics and pharmacokinetics of MZ and AZ differ due to an additional methyl group added to the thiadiazole ring in AZ (Fig. 1). The methyl group leads to greater lipophilicity and reduced plasma-binding capacity compared to AZ (Maren, 1967). The increased lipophilic properties of MZ promote greater distribution throughout the body, including greater localization in brain tissue (Maren, 1967). Due to their physical-chemical differences, smaller doses of MZ relative to AZ (e.g., oral dose of 200–250 mg/day for MZ vs. 500–750 mg/day for AZ) produce similar levels of CA inhibition. Human studies directly comparing AZ and MZ at clinically relevant doses found similar increases in arterial oxygenation, but greater blood pH reductions with AZ (Boulet et al, 2018; Dominelli et al, 2018; Teppema et al, 2020).
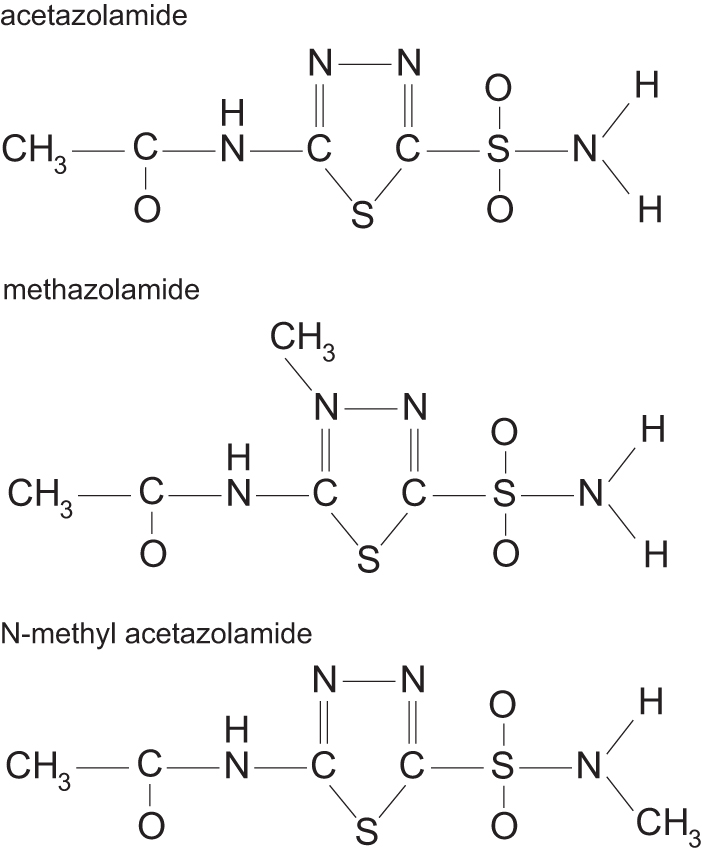
Chemical structures of AZ, MZ, and N-methylacetazolamide. AZ, acetazolamide; MZ, methazolamide.
The greater acidosis observed with AZ may be explained by differences in pharmacokinetics, resulting in the administration of doses with unequal potency with respect to intracellular CA inhibition. Since the more lipophobic AZ binds to plasma proteins to a greater degree, AZ may accumulate more in the kidneys, increasing renal CA inhibition. In support of this, AZ reduces plasma [K+] to a greater extent than MZ, most likely due to an increased diuretic effect (Teppema et al, 2020). It is possible that the induced metabolic acidosis could influence exercise performance (Ferguson et al, 2018). However, work by others does not find a significant effect of hyperchloremic and lactic acidosis on diaphragm muscle function, suggesting the impact on locomotive muscle performance may be minimal (Michelet et al, 2015; Yanos et al, 1993).
When treating AMS, targeting renal CA is important to produce the desired increase in ventilation and oxygenation without any undue side effects (Swenson, 2016). Due to the organic acid concentrating capacity of the kidneys, a low dose of AZ (1–5 mg/kg) is all that is required to effectively inhibit renal CA (Swenson, 1998). Low doses of AZ will also inhibit membrane-bound CA located on vascular endothelium, leading to a slight rise in tissue PCO2 and stimulating ventilation via peripheral chemoreceptors (Klocke, 1997). Increases in cerebral blood flow secondary to the rise in PCO2 minimizes the role of central chemoreceptors in increasing ventilation due to greater washout of CO2 from the brain.
Greater doses of AZ are required to penetrate and fully inhibit CA isoenzymes (I and II) of erythrocytes (>15 mg/kg) and to cross the blood brain barrier (>20 mg/kg), directly affecting the central nervous system (Carrion et al, 2001; Swenson, 1998); both of which are much greater than the effective dose to treat AMS (Kayser et al, 2012; Swenson, 2014). Unfortunately, there is considerably less work to determine the ideal dosing range to treat AMS with MZ. Similarities in physical-chemical properties between AZ and MZ (Fig. 1) suggest that they could share similar mechanisms of action. If MZ has a similar mechanism of action as AZ, it is reasonable to speculate that only a low dose of MZ is required to selectively inhibit renal CA enzymes and increasing dosage will lead to erythrocyte and central nervous system CA inhibition.
It is unclear whether differences in CA inhibitor dosing affect hypoxic and high-altitude exercise performance. For example, the same dose of AZ (i.e., oral 500 mg/day) has shown improvements in submaximal exercise performance (Bradwell et al, 1986), no impact on time trial performance (Bradbury et al, 2020), and decrements in maximal and submaximal exercise performance (Stager et al, 1990) (Fig. 2, Tables 1 and 2). Thus, other aspects other than dose (e.g., differences in study design) are likely responsible for any differences in performance, which may even include non-CA inhibiting actions (Swenson, 2016).
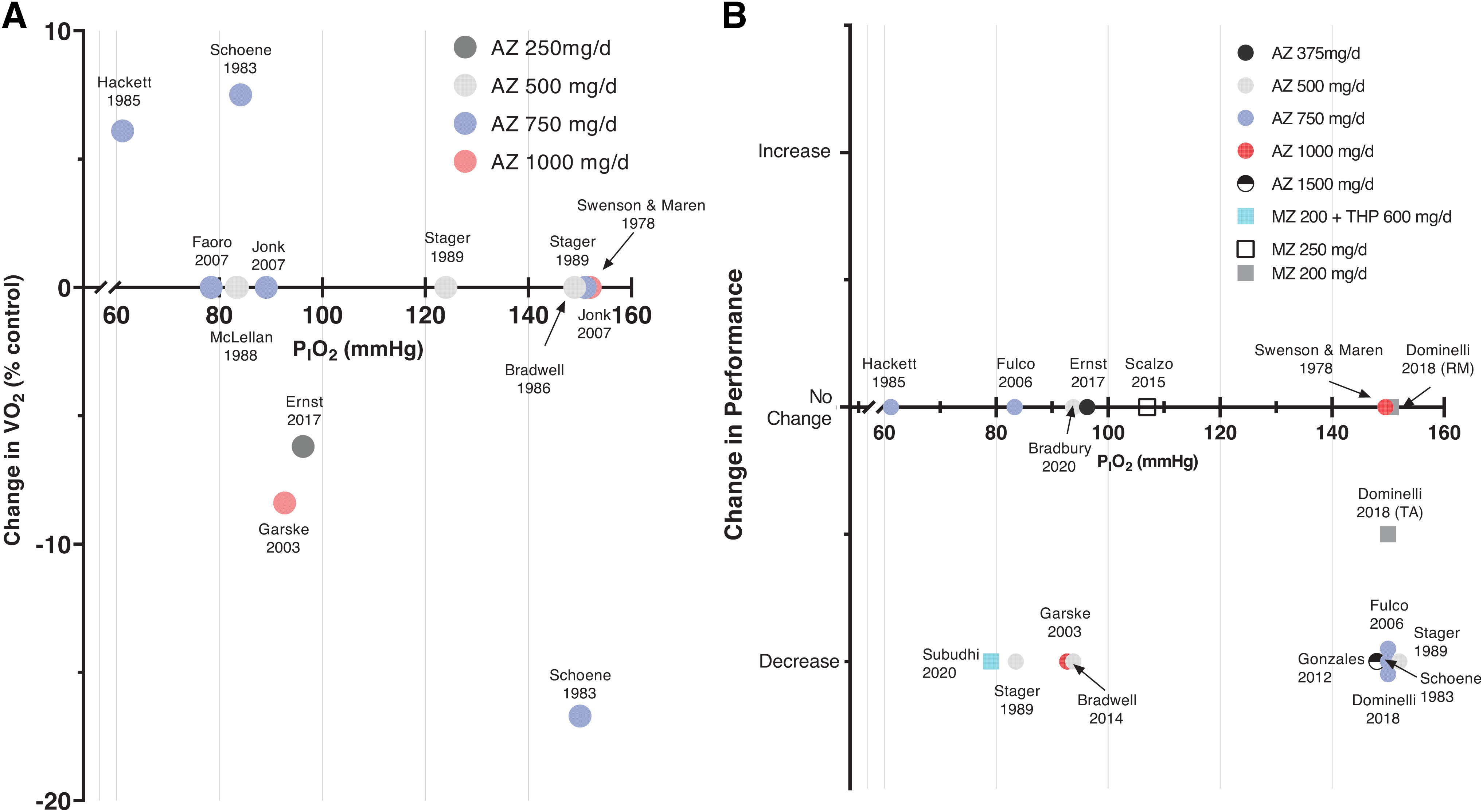
Summary of studies assessing the effect of AZ on maximal oxygen uptake (V̇O2max) and performance outcomes (i.e., Time to exhaustion, exercise duration, maximal power output).
Maximal Exercise Responses of Acetazolamide Compared to Placebo
b.i.d., twice a day; CE, cycle ergometer; DBP, diastolic blood pressure; HH, hypobaric hypoxia; HR, heart rate; LA, blood lactate concentration; NH, normobaric hypoxia; o.d.; once a day; PaCO2, partial pressure of carbon dioxide; PaO2, arterial oxygenation; PO, power output; Q, cardiac output; RER, respiratory exchange ratio; RPE, rate of perceived exertion; RPE-B, perceived dyspnea; RPE-L, perceived leg discomfort; SBP, systolic blood pressure; SM, semirecumbent; SpO2, oxyhemoglobin saturation; Tmax, maximum exercise time; t.i.d., three times a day; TM, treadmill; V̇E, minute ventilation; V̇CO2, carbon dioxide output; V̇O2, oxygen uptake; WR, work rate.
Submaximal Performance Outcomes of Acetazolamide Compared to Placebo
TT, time trial.
AZ Impact on Normoxic Exercise Performance
Exercise performance is influenced by a multitude of factors. Due to CA inhibition, AZ affects arterial oxygenation, blood and cellular pH, and respiratory muscle metaboreflex; all of which can either impair or improve performance to some extent. A common finding, however, is that in normoxic conditions, AZ significantly reduces exercise capacity compared to placebo [Fig. 2A, B, Tables 1 and 2; (Fulco et al, 2006; Gonzales and Scheuermann, 2013; Schoene et al, 1983; Stager et al, 1990; Swenson and Maren, 1978)]. Below, we describe the likely physiological mechanisms behind this finding.
First, while AZ increases PaO2 due to greater ventilation, exercise performance decrements are observed in normoxic conditions (Tables 1, 2 and Fig. 2A). Indeed, studies have demonstrated both reductions in V̇O2max (Schoene et al, 1983) and shorter time to exhaustion (Stager et al, 1990) with AZ relative to placebo. With increased ventilation, improved pulmonary gas exchange, and arterial oxygenation in normoxia (Jonk et al, 2007), it might be hypothesized that CA inhibitors are beneficial to exercise performance by improving oxygen delivery to the muscles. In normoxia, however, hemoglobin is typically fully saturated with oxygen, thus any further increase in alveolar oxygen pressure and PaO2 does not meaningfully improve CaO2.
Second, muscle fatigue may be associated with changes in acid-base muscle cell status induced by AZ. AZ could potentially reduce exercise performance by impairing bicarbonate buffering of blood pH. CA inhibition with AZ significantly lowers blood pH, and plasma bicarbonate concentration (Fulco et al, 2006; Jonk et al, 2007; Stager et al, 1990; Swenson and Maren, 1978) both at rest and during exercise. Lower blood lactate concentrations with AZ intake can be explained by more intracellular retention of H+ and lactate in the muscle (Becker and Deitmer, 2008). One study found inducing metabolic alkalosis with bicarbonate supplementation increased total work performed during supramaximal exercise (Messonnier et al, 2007).
The greatest improvements in exercise performance with bicarbonate were observed in subjects with lower levels of proteins (monocarboxylate transporter −1, CA II, CA III, and Na+/H+ exchanger) mediating muscle cell pH regulation and transport of lactate/protons, suggesting that acid-base balance in skeletal muscle plays a role in muscle fatigue (Messonnier et al, 2007). However, other work suggests that the extent of fatigue development is not proportional to reductions in blood pH (Westerblad, 2016). There are also no differences in plasma bicarbonate levels in moderate and heavy intensities (>75% V̇O2max) (Stager et al, 1990), suggesting that decreased buffering capacity does not significantly affect performance.
With increasing exercise intensity, greater amounts of metabolites, such as lactate and H+, are produced. While the accumulation of metabolites coincides with the onset of muscle fatigue, the rate of perceived exertion is not altered with changes in blood lactate (Miller et al, 2002). Likewise, there are no changes in V̇O2max with AZ despite a 50% reduction in peak blood lactate increases (Jonk et al, 2007). Altogether, muscle tissue acid-base status appears to be an important contributor to exercise performance while the role of blood acid-base status remains unclear.
Third, distribution of cardiac output and blood flow is mediated by sympathetic outflow, which involves certain metabo- and mechanoreflexes (Mitchell and Schmidt, 2011). For example, increased airway resistance is accompanied by decreased blood flow and increased vascular resistance in the limbs as a result of respiratory muscle metaboreflex (St Croix et al, 2000). As AZ increases V̇E for a given work rate in normoxia, a plausible mechanism for performance decrement may be the redistribution of cardiac output away from active locomotor muscles to respiratory muscles. At a given submaximal exercise intensity where there is no change in V̇O2, increased V̇E coincides with a decrease in time to exhaustion (Fulco et al, 2006; Stager et al, 1990).
However, leg V̇O2 and blood flow were observed to be similar during exercise with AZ compared to placebo (Jonk et al, 2007), which suggests that the increased V̇E with AZ is insufficient to affect blood flow to locomotor muscles due to the respiratory muscle metaboreflex.
AZ Impact on Hypoxic Exercise Performance
While AZ negatively affects exercise in normoxia, the results are inconclusive with hypoxic exercise (Fig. 2, Tables 1 and 2). As indicated above, exercise performance is multifaceted and is influenced by psychosomatic factors. For example, perceived exertion during hypoxic exercise can be negatively influenced by the presence of AMS symptoms, such as headache, dizziness, and overall fatigue (Bärtsch and Swenson, 2013). AZ could potentially improve performance in hypoxia due to AMS symptom attenuation, rather than through physiological or biochemical alterations of performance per se. However, at a given exercise intensity in hypoxia, perceived leg discomfort increases with AZ compared to placebo (Garske et al, 2003). Thus, while AMS symptoms may be lessened, other exercise-limiting symptoms may be exacerbated.
From a physiological perspective, hypoxic exposure alone increases V̇E in the attempt to raise CaO2, but AZ causes a further 10%–20% increase in V̇E compared to placebo, which additionally increases SpO2 (Bradbury et al, 2020; Fulco et al, 2006; Garske et al, 2003; Hackett et al, 1985; Jonk et al, 2007; Schoene et al, 1983). Likewise, in instances of no increase in V̇E, there is also no change in SpO2 (Stager et al, 1990). Although these findings are consistent across most studies on AZ, their impact on performance outcomes is more variable.
While increased ventilation can increase V̇O2 by raising SpO2 and CaO2, this is not a direct relationship. With higher peak V̇E and SpO2 during maximal exercise, numerous studies report no significant differences in V̇O2 max (Bradbury et al, 2020; Faoro et al, 2007; Hackett et al, 1985; Jonk et al, 2007), others an increase in V̇O2 max (7.5%) (Schoene et al, 1983), or decrease (8.7%) (Garske et al, 2003) (Fig. 2A). The decrease in V̇O2 max may be due to higher dosage of AZ than the others (i.e., 1,000 mg/day vs. 500–750 mg/day) (Garske et al, 2003). A higher AZ dose is more likely to critically inhibit intracellular CA in red blood cells and tissues, increasing local CO2 retention, whereas a lower dose will only target CA in kidneys and vascular endothelium (Swenson, 1998).
The improvement in V̇O2 at high altitude may be attributed to potentially greater increases in CaO2 for a given V̇E increase than at lower altitude due to the shape and steeper slope of the oxyhemoglobin dissociation curve, over which gas exchange occurs in the lung and tissues at high altitude compared to low altitude. We caution, however, our rationale as follows does not account for metabolic changes that impact the shape of the dissociation curve. For instance, at a PIO2 of 84 mmHg, resting SpO2 is around 66% (Schoene et al, 1983), while resting SpO2 is 79% at a PIO2 of 94 mmHg (Bradbury et al, 2020). At a SpO2 of 66%, PaO2 falls on the steeper portion of the oxyhemoglobin dissociation curve, where relatively minor increases in PaO2 will increase SpO2 and thus CaO2 to a greater degree, due to relatively higher oxyhemoglobin affinity.
In contrast, at a SpO2 of 79%, oxyhemoglobin affinity is relatively less on the same sigmoidal oxyhemoglobin dissociation curve, thus a similar change in PaO2 brings about less improvements in CaO2. Furthermore, higher altitudes will lead to a lower PaCO2 from increased V̇E, causing a relative left-shift on the oxyhemoglobin dissociation curve. Thus, for a similar magnitude of change in PaO2, there is greater binding affinity of oxygen to hemoglobin at higher altitudes, but at the cost of lesser ease of off-loading to the tissues. At submaximal intensities, many studies observe an increase in V̇E and SpO2 at any given work rate, but there are no differences in submaximal V̇O2 between AZ and placebo (Fulco et al, 2006; Garske et al, 2003; Jonk et al, 2007; McLellan et al, 1988).
In part, this may be due to partial red cell CA inhibition slowing the rate at which the Bohr effect occurs during capillary transit to yield greater O2 offloading (Maren and Swenson, 1980). Similarly, at the level of the active muscles, there are no differences in leg V̇O2 despite a 10% increase in V̇E (Jonk et al, 2007). These results further suggest that increases in V̇E arising from CA inhibition do not translate to increases in V̇O2. Another potential explanation is that the magnitude of SpO2 increases is insufficient to improve V̇O2. Specifically, to affect V̇O2, there needs to be at least a ∼3% change in SpO2 (Harms et al, 2000).
However, it is important to note that V̇O2max is not the definitive predictor of exercise performance. For instance, despite no difference in time to exhaustion compared to placebo, V̇O2max can increase by 7.5% (Schoene et al, 1983) or decrease by 6% (Ernst et al, 2017) with AZ intake. Conversely, with no significant changes in V̇O2max, time to exhaustion can decrease by 26% (Stager et al, 1990) or stay the same (Bradbury et al, 2020). A possible explanation for changes in V̇O2max but no changes in time to exhaustion may be that higher ventilation requires greater blood flow and total oxygen delivery to respiratory muscles (Harms et al, 1997). The disparity between V̇O2 and performance can be partially attributed to the dissociation between oxygenation and performance.
Although all participants taking AZ showed an increase in resting SpO2, ∼56% of participants were unable to complete the exercise protocol (Bradwell et al, 2014). Thus, while maximal oxygen uptake is an important indicator of aerobic capacity, these findings suggest that O2 delivery and consumption cannot fully account for the changes in exercise performance at altitude caused by AZ.
While increased V̇E due to AZ is beneficial to arterial oxygenation, it can be a possible decrement by increasing perceived dyspnea (Hackett et al, 1985). However, this mechanism during exercise is not fully supported. During hypercapnic ventilatory response testing, AZ reduces dyspnea perception for a given V̇E compared to placebo (Garske et al, 2003). For a given exercise intensity, AZ increases V̇E by 16%, but does not increase perceived dyspnea (Garske et al, 2003). The observed dissociation between perceived dyspnea and V̇E may be explained by AZ's ability to affect neural afferent transmission (Brechue et al, 1997). As perceived exertion is also attributed to afferent feedback from active muscles, impairing transmission from afferent receptors will reduce sensory information and alter perceived exertion.
While there are no differences in maximal V̇E and end-exercise dyspnea, there is a 10% decrease in exercise duration with AZ (Garske et al, 2003). Similarly, where there are no significant differences in V̇E, time to exhaustion is reduced by 26% in AZ compared to placebo (Stager et al, 1990). Contrary to the original hypothesis, a drop in exercise performance is still present without significant changes in V̇E and/or perceived dyspnea.
A decrease in V̇CO2 and/or increase in the V̇E/V̇CO2 ratio is also observed with AZ during exercise (Bradbury et al, 2020; Bradwell et al, 2014; Bradwell et al, 1986; Faoro et al, 2007; Garske et al, 2003; Jonk et al, 2007; McLellan et al, 1988; Stager et al, 1990). As a result of HCO3− excretion, reduced buffering capacity may impair CO2 transport, as indicated by decreased V̇CO2 for a given work rate (Garske et al, 2003; Jonk et al, 2007; McLellan et al, 1988). In addition, inhibition of red blood cell CA is found to increase CO2 retention in tissues (Garske et al, 2003), but this is unlikely at therapeutic doses of AZ (Swenson and Maren, 1978).
In one study, during steady state exercise, participants taking AZ had similar V̇CO2 compared to placebo, but there was a significantly slower rise in V̇CO2 during the transition to higher work rates (Scheuermann et al, 1999). This observation may explain the reduction of V̇CO2 in the studies that applied a ramp or incremental exercise protocol. With shorter increments between increases in work rate, AZ does not allow V̇CO2 at the lungs to reach a “true” steady state value due to CO2 retention and delayed CO2 transport from tissues.
Thus far, the mechanism of impaired CO2 transport and buffering capacity on performance is unknown and requires greater investigation, but presently a reduction in V̇CO2 is consistent across different altitudes and doses of AZ. Another explanation for the reduction in V̇CO2 could be a change in fuel utilization by AZ from carbohydrate to fat as shown by Hirahara et al (1995) in resting subjects. However, whether this is the case during exercise was not investigated in that study.
MZ and Exercise Performance
Studies examining the impact of MZ on exercise performance in humans can be found in Table 3. In one study, the impact of MZ, theophylline, and a combination of MZ and theophylline on a 12.5 km cycling time trial in normoxia and hypoxia (PIO2 ∼105 mmHg) was examined in men (Scalzo et al, 2015). Theophylline is a general phosphodiesterase inhibitor and an adenosine receptor blocker, and when combined with MZ, has been suggested to improve exercise performance (Radiloff et al, 2012). The change in time trial performance from normoxia to hypoxia was not statistically different between the individual treatments of MZ and theophylline compared to the control condition.
Submaximal Performance Outcomes of Methazolamide Compared to Placebo
Methazolamide.
Theophylline.
AZ, acetazolamide; MZ, methazolamide.
However, combined MZ and theophylline treatment significantly attenuated the decrease in time trial performance from normoxia to hypoxia (37 seconds) relative to the control condition (180 seconds). The improvement in performance was attributed to the combined effects of MZ improving arterial oxygenation and theophylline improving systemic circulation by its actions on adenosine receptors (Dayan et al, 2016; Leaf and Goldfarb, 2007). However, there are some limitations with the study by Scalzo et al (2015). First, despite matching for age, body composition, and V̇O2max between experimental conditions, this was not a randomized crossover-controlled trial study design. Each experimental condition group was composed of a different set of individuals, making it difficult to solely attribute the study findings to the effect of the drugs. Second, no measures of V̇E or blood hemoglobin concentration were reported, making it difficult to elucidate the mechanism in which oxygenation was improved.
In a follow-up study, Subudhi et al (2021) performed a repeated measures crossover study and examined a time trial performance in hypobaric hypoxia (PIO2 ∼80 mmHg) with and without combined MZ and theophylline treatment. In contrast to the study by Scalzo et al (2015), all participants completed the 12.5 km with longer times (+51 seconds) with the combined drug treatment compared to control, despite significant improvements in arterial oxygenation (SpO2 +4%). In addition, with MZ and theophylline, greater heart rates were attained for a given power output compared to the control condition, suggesting greater levels of fatigue (Subudhi et al, 2021). Unfortunately, the individual effect of each drug was not investigated, making it difficult to discern the mechanism for reduced exercise performance and increased fatigability.
To directly investigate the impact of MZ and AZ on the fatigability of respiratory and locomotor muscles, another study examined muscle function following isolated small muscle group fatiguing exercise in normoxia (Dominelli et al, 2018). Individuals completed fatiguing dorsiflexion exercise and inspiratory resistance exercise with and without MZ or AZ. Treatment with MZ resulted in less impairment of baseline muscle function compared to AZ. Three minutes postexercise, tibialis anterior fatigue was significantly greater with AZ (∼65% of baseline twitch torque response) compared to MZ and placebo (∼78% and ∼71% of baseline twitch torque response, respectively) (Dominelli et al, 2018). Likewise, respiratory muscle fatigue was greater with the AZ treatment compared to MZ and placebo after 3 minutes of exercise (∼83%, ∼86%, and ∼92% of baseline transdiaphragmatic pressure, respectively).
Although the study provides objective evidence that muscle function is better preserved during fatiguing exercise with MZ, exercise was only performed in isolated small muscle-mass tissues, making the findings difficult to extrapolate to whole body exercise. In addition, with the focus on determining the oxygen-independent mechanisms limiting muscle fatigue development, the testing protocol was limited to normoxic conditions. During exercise at sea level, SpO2 remains high (∼95%–98%), thus the increased V̇E with CA inhibition will not appreciably affect arterial oxygenation. However, during exercise in hypoxia, reduced arterial oxygenation will lead to greater development of muscle fatigue (Amann et al, 2006). It remains to be investigated if the differences in muscle fatigue development with the drug will persist during exercise in hypoxia.
Secondary Effects of AZ and MZ
Both AZ and MZ have secondary effects outside of their primary function to inhibit CA that could influence exercise performance. Hypoxia exposure increases pulmonary vascular resistance and pulmonary artery pressure (PAP) in humans and other mammals due to hypoxic pulmonary vasoconstriction (HPV) (Swenson, 2013). Several studies in animals and humans have demonstrated that AZ attenuates HPV (Boulet et al, 2018; Höhne et al, 2007; Pickerodt et al, 2019; Pickerodt et al, 2014; Teppema et al, 2007; Teppema and Swenson, 2015; Tremblay et al, 2015). Animal studies further suggest that the reduction in HPV is independent of CA inhibition (Höhne et al, 2007; Pickerodt et al, 2014; Shimoda et al, 2007).
In unanesthetized dogs exposed to acute hypoxia, N-methyl-AZ , a non-CA inhibiting AZ analog (Fig. 1), reduced PAP (Pickerodt et al, 2014). MZ appears to share this action as pulmonary vascular sensitivity to hypoxia was lessened similarly to AZ in humans at rest (Boulet et al, 2018) and in dogs (Pickerodt et al, 2014). However, reducing HPV does not necessarily improve exercise performance as individuals given sildenafil, a known pulmonary vasodilator and HPV inhibitor, did not produce any improvement in completing a 16.1 km time trial during hypoxia (Carter et al, 2019).
AZ and MZ also have similar effects on the control of breathing (Teppema et al, 2020). Both AZ and MZ left shift the hypercapnic ventilatory response, while the slope of the relationship between the hypoxic sensitivity and H+ concentration is similar between placebo, AZ, and MZ (Teppema et al, 2020). Compared to rest, HVR is greater during submaximal cycling exercise (Constantini et al, 2021). However, the increased gain in HVR during exercise was not correlated with changes in 10-km time trial performance from normoxia to hypoxia; suggesting that shifts in control of breathing from rest to exercise has little impact on exercise performance (Constantini et al, 2021). Further insight into how AZ and MZ affect exercise performance in normoxia and hypoxia could be gleaned in future studies examining the impact of metabolic acidosis produced by other means (e.g., ingestion of ammonium chloride).
AZ increased fatigue in both respiratory and locomotor muscles compared to control, but MZ attenuated respiratory muscle fatigue relative to AZ resulting in similar levels of locomotor muscle fatigue as control (Dominelli et al, 2018). In addition, studies in rodents found greater preservation of muscle tension with MZ than AZ following fatiguing muscle stimulation (Côté et al, 1989; Frémont et al, 1991). In a mechanism independent of CA inhibition, MZ may affect muscle function differently because, unlike AZ, it is not an effective big potassiu (BK) channel opener (Tricarico et al, 2013; Tricarico et al, 2004). BK channels are found in the T-tubules of skeletal myocytes and play a critical role in maintaining cellular electrochemical gradient and electrolyte homeostasis. Previous work has shown that increased BK channel activity will decrease muscle force during continuous contractions (Kristensen et al, 2006).
Both hypoxic exposure and high-intensity exercise will increase reactive oxygen species (ROS) production (Bailey et al, 2009; Powers et al, 2020). Oxidative stress can be beneficial by promoting muscle hypertrophy (Powers et al, 2020), but high levels of ROS can result in muscle contractile dysfunction and fatigue (Powers et al, 2011). Increased ROS in hypoxia may also play a role in the development of AMS (Bailey et al, 2009). There is some evidence to suggest that AZ and MZ have antioxidant properties, helping to scavenge ROS in normoxia and hypoxia (Bailey et al, 2012; Prouillac et al, 2009; Shimoda et al, 2021). In mice, MZ reduced oxidative stress and cellular apoptosis by inhibiting caspase-3 (Li et al, 2016).
Others found MZ, but not AZ, upregulated nuclear-related factor-2, a regulatory transcription factor for the production of antioxidant proteins (Lisk et al, 2013). Recent work in humans found AZ to have an improved nonenzymatic plasma antioxidant capacity, but not MZ (Boulet et al, 2018). However, clear conclusions on the antioxidative properties of AZ and MZ from this study should be taken with caution as a difference in small plasma antioxidants did not influence overall markers of oxidative stress or the attenuation of HPV (Boulet et al, 2018).
Future Directions
The impact of AZ on exercise performance in hypoxia is not fully understood as reflected by inconsistent findings (Tables 1 and 2). One possible explanation for the variability in exercise performance could be the differences in methodology. At different levels of hypoxia (i.e., PiO2 of 70 mmHg vs. 100 mmHg), the impact of CA inhibition on exercise performance will differ (Fig. 2A, B). Based on where gas exchange occurs on the oxyhemoglobin dissociation curve at lower altitudes, the improvement in arterial oxygenation with CA inhibition is likely smaller than that at higher altitudes. Thus, the deleterious effects of AZ on exercise performance observed in normoxia may be more prevalent at lower altitudes, compared to higher altitudes.
In addition, the diuretic impact of CA inhibition may have performance implications. A benefit of reduced extracellular and plasma volume is an increased hematocrit, thus improving oxygen carrying capacity (Nepal et al, 2012). However, blood volume loss can adversely impact exercise performance due to reductions in cardiac output and potential dehydration (Barr, 1999). More recent studies have addressed this confounding factor by implementing hydration protocols and measuring urine-specific gravity and urine volume (Ernst et al, 2017; Faoro et al, 2007). While dehydration was intentionally addressed, the results were still inconclusive as V̇O2max decreased with mild hypoxia (Ernst et al, 2017), while there were no differences in maximal V̇O2 and power output at higher altitudes (Faoro et al, 2007).
AZ is the most widely prescribed CA inhibitor to prevent and treat AMS. Conversely, very few studies have examined the use of MZ in preventing and treating AMS during expeditions to high altitudes. Despite this paucity of studies, MZ appears to share similar mechanisms of action as AZ in preventing and treating AMS, suggesting that it could be equally effective in preventing AMS (Forster, 1982; Wright et al, 1994). While MZ is known to be more potent at equimolar dosing, more studies examining the use of MZ at preventing and treating AMS are warranted to confirm its efficacy and its critical dosing levels.
Conclusions
Several conclusions can be drawn from this review evaluating the effect of AZ and MZ on exercise performance. First, AZ hinders exercise performance in normoxia. Such a result is expected because in healthy humans, hemoglobin is typically fully saturated with oxygen during sea level exercise and increasing ventilation via partially compensated metabolic acidosis serves little benefit and may obligate a greater diversion of cardiac output to the nonlocomotor respiratory muscles. Second, the impact of AZ on hypoxic exercise performance is equivocal and can be influenced by multiple factors. AZ appears to bring more benefit to performance at higher altitudes compared to lower altitudes.
Third, MZ appears to have similar prophylactic benefits for AMS and there is indirect evidence that it may result in less exercise performance decrements than AZ. Overall, we suggest that MZ may be a better alternative CA inhibitor when exercise performance is crucial at high altitude, however, this hypothesis still awaits direct experimental testing in a head-to-head randomized controlled crossover studies.
Footnotes
Authors' Contributions
All authors contributed to the drafting, reviewing, and editing of the article. All authors approved the final submission.
Author Disclosure Statement
No competing financial interest exists.
Funding Information
No funding was received for this article.