
Abstract
Murphey, Joshua T., Jennifer L. Temple, and David Hostler. Taste and appetite at altitude: A comprehensive review of sensory and hunger modulation in high-altitude environments. High Alt Med Biol. 26:393–407, 2025.
Introduction:
Individuals living or working at high altitudes typically experience altered taste perceptions and reduced appetite. These changes can lead to nutritional deficiencies, affecting the energy balance and body composition.
Methods:
We conducted a nonsystematic review of PubMed to explore these phenomena and expound on their findings to offer additional insights.
Results:
Changes in taste and perception are common and typically lead to loss of mass. There are limited practical solutions to mitigate these challenges.
Discussion:
Gradual acclimatization and tailored nutritional strategies are required to enhance health and performance in high-altitude environments. This review provides critical insights into the intersection of altitude, nutrition, and health.
Introduction
Living, recreating, and working at moderate to high altitudes impose physiological and psychological challenges that do not occur when engaged in sea level activities. Adaptation to altitude, more commonly referred to as acclimatization, becomes increasingly difficult as elevation increases due to lower partial pressures of oxygen (PO2) in the air. The adverse effects of altitude exposure manifest as high-altitude headache (HAH), acute and chronic mountain sickness (AMS and CMS), high-altitude pulmonary edema (HAPE), and high-altitude cerebral edema (HACE), along with less severe side effects such as ageusia (loss of taste) and anosmia (loss of smell). These adverse effects are exacerbated by increased physical activity save for CMS (Bärtsch et al., 2005; Imray et al., 2010; Toussaint et al., 2021; Turner et al., 2021, 2021). Previous studies have examined the pathophysiology of altitude exposure with varying results. Individual propensities for high-altitude affinity, defined as physiological tolerance to direct exposure to hypobaric conditions, vary considerably. The underlying mechanisms for these disparities remain elusive (Beidleman et al., 2019; Treml et al., 2020; Mallet et al., 2023). As outlined in the Wilderness Medical Society’s guidelines for preventing and treating altitude sickness (2023), the current consensus advocates incremental altitude adjustments over extended durations as the most effective method for acclimatization at moderate to high altitudes (Luks et al., 2023). However, this poses a problem as many climbers, workers, and warfighters may not have the option of slow/incremental altitude exposure.
Altitude acclimatization practices are implemented to prevent altitude sickness. However, they do not blunt the changes to olfaction and taste, leading to decreased hunger and diminished smell perception (Singh et al., 1997; Matu et al., 2017c; Huppertz et al., 2018). These secondary effects of hypobaric exposure, coupled with hypoxic-induced anorexia, result in an energy deficit that ultimately alters energy balance. Various military organizations have studied this phenomenon. Notably, the U.S. Naval School of Aviation Medicine initiated an investigation into the physiological stresses experienced in a low-pressure environment, simulating an ascent to 8,848 m, equivalent to the elevation of Mount Everest. The primary objectives were to determine whether humans could reach such an altitude breathing ambient air and to evaluate if both unacclimated and acclimated individuals could achieve even higher elevations with supplemental oxygen (O2). About 35 years after this, Operation Everest II was launched, aiming not only to emulate the procedures of Operation Everest but also to broaden the scope to examine a wide range of physiological responses to high altitude (Houston, 1997).
Eight male participants were exposed to a progressively greater hypobaria over 40 days during Operation Everest II. Rose et al. (1988) reported on the impact of hypoxia on energy balance, food preferences, and body composition. Participants were provided three meals daily, approximately 3,000 kcal in total, with the option to consume snacks and fluids as desired. Precise measurements were made of all consumed food and liquids (Rose et al., 1988). Daily energy expenditure was assessed through resting metabolic rate and required exercise sessions. The 40-day hypobaric exposure decreased caloric intake by 43%—furthermore, energy expenditure during exercise sessions diminished in correlation with increasing altitude. The average weight loss was 7.4 kg among subjects, which could only partially be reconciled by energy intake and expenditure. It was reported that 67% of the weight loss was attributed to reduced fat-open-access mass regardless of protein intake and despite being fully hydrated during the exposure. The hypoxic anorexia observed among participants was hypothesized to result from insufficient O2 availability, diminished appetite, and constrained exercise capability (Rose et al., 1988; Houston, 1997).
Operation Everest II’s findings paved the way for a collaborative multiagency research initiative: Operation Everest-Comex, also known as Operation Everest III. During this iteration, Westerterp-Plantenga et al. (1999) postulated that the energy deficit observed upon exposure to hypobaric hypoxia resulted from decreased food consumption (Westerterp-Plantenga et al., 1999). This reduction in caloric intake is presumably influenced by alterations in appetite and the subjects’ disposition toward eating in conditions of diminished atmospheric pressure. Participants had the option of consuming food and beverages ad libitum. All consumables were preportioned, and any remnants were measured to ensure accurate energy consumption tracking for each individual. Throughout the 31-day exposure to hypobaric hypoxia, there was a reduction in body mass (−5.0 ± 2.0 kg), a consistent decrease in energy intake (−7.3 ± 1.0 MJ/day), and a drop in energy balance (−4.0 ± 1.7 MJ/day). Participants increased their meal frequency from four to seven instances daily, and there was a notable increase in calories consumed from snacks versus conventional meals. The research team proposed that the observed decrease in food consumption and appetite could be attributed to fluctuations in serum leptin and ghrelin levels due to prolonged hypoxia (Westerterp-Plantenga et al., 1999). These early studies show that energy imbalance at altitude is potentially unavoidable and difficult to limit, which poses a problem for warfighters who must operate in an alpine environment. As energy balance decreases at altitude (Berryman et al., 2018), performance suffers (Matu et al., 2017c; Berryman et al., 2018). Understanding the causes of these secondary effects for individuals required to work at altitude is important so that effective interventions can be identified.
During the early stages of this review, a detailed search strategy was implemented to find appropriate literature sources, and the steps were followed systematically. The research flow diagram (Fig. 1) is provided to show the process followed from identifying 182 potential studies based on database searching. After initial exclusion and deduplication, 135 studies underwent full-text review, from which 98 studies adhered to the inclusion criteria. For this, the flow diagram shows the screening process, allowing only high-quality and relevant research to be included in the review. Highlighted studies are shown in Tables 1–4. This review examines how altitude exposure affects taste, hunger, energy balance, and performance.

Reference flow diagram.
Physiological and Psychological Effects of Altitude and Hypoxia

High altitudea, chronic intermittent hypoxiab, cerebral blood flowc, hypoxic inducible factor-alphad, mu-opioid receptore, delta-opioid receptorf.
High-Altitude Impacts on Appetite and Hunger
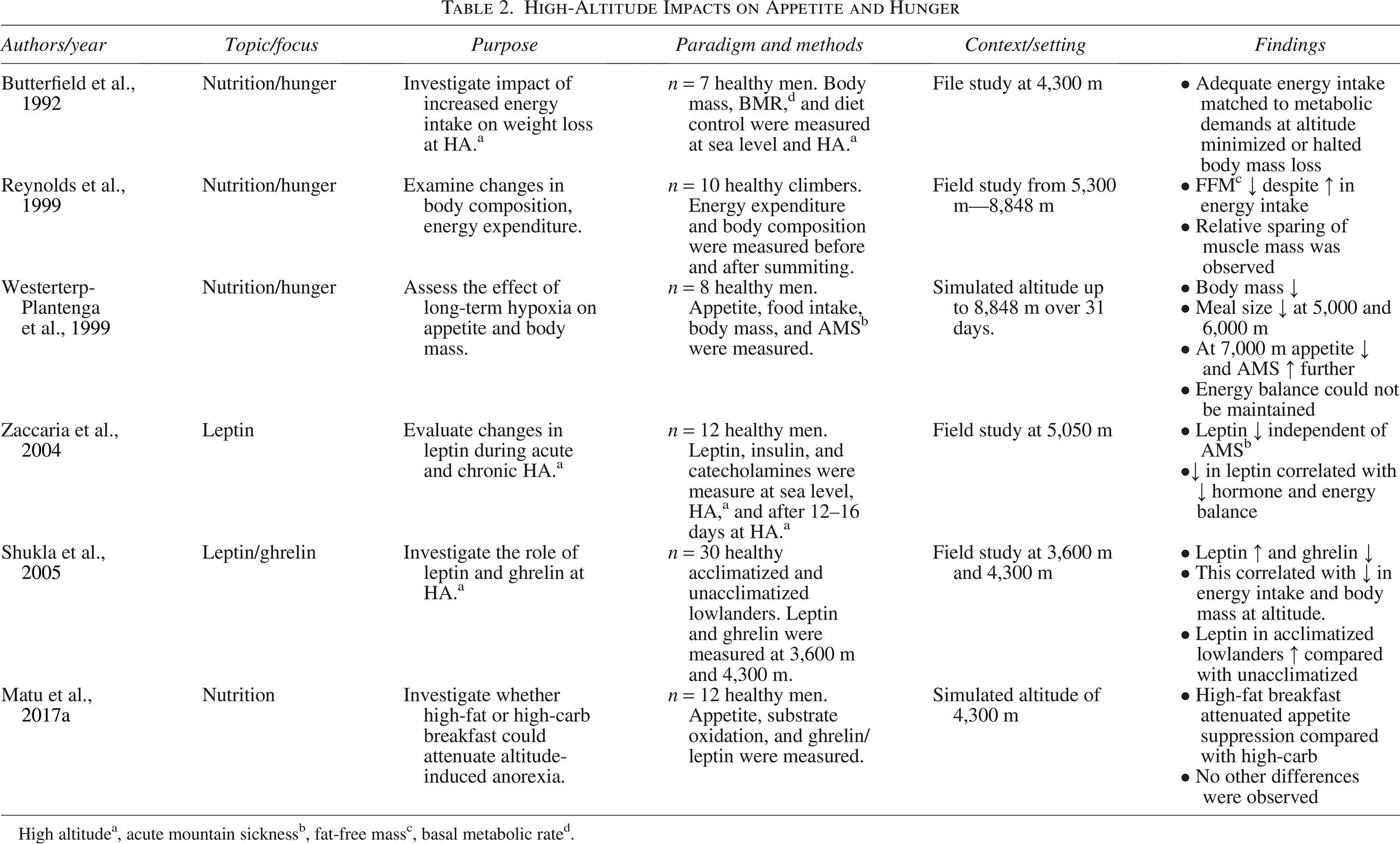
High altitudea, acute mountain sicknessb, fat-open-access massc, basal metabolic rated.
High-Altitude Impacts on Taste and Hormonal Balance

High altitudea, acute mountain sicknessb, fat-open-access massc, thyroid-stimulating hormoned, open-access triiodothyronine, open-access thyroxinee, follicle-stimulating hormonef, luteinizing hormoneg, growth hormoneh, body compositioni.
Physiological Implications of Altered Appetite and Taste at High Altitude
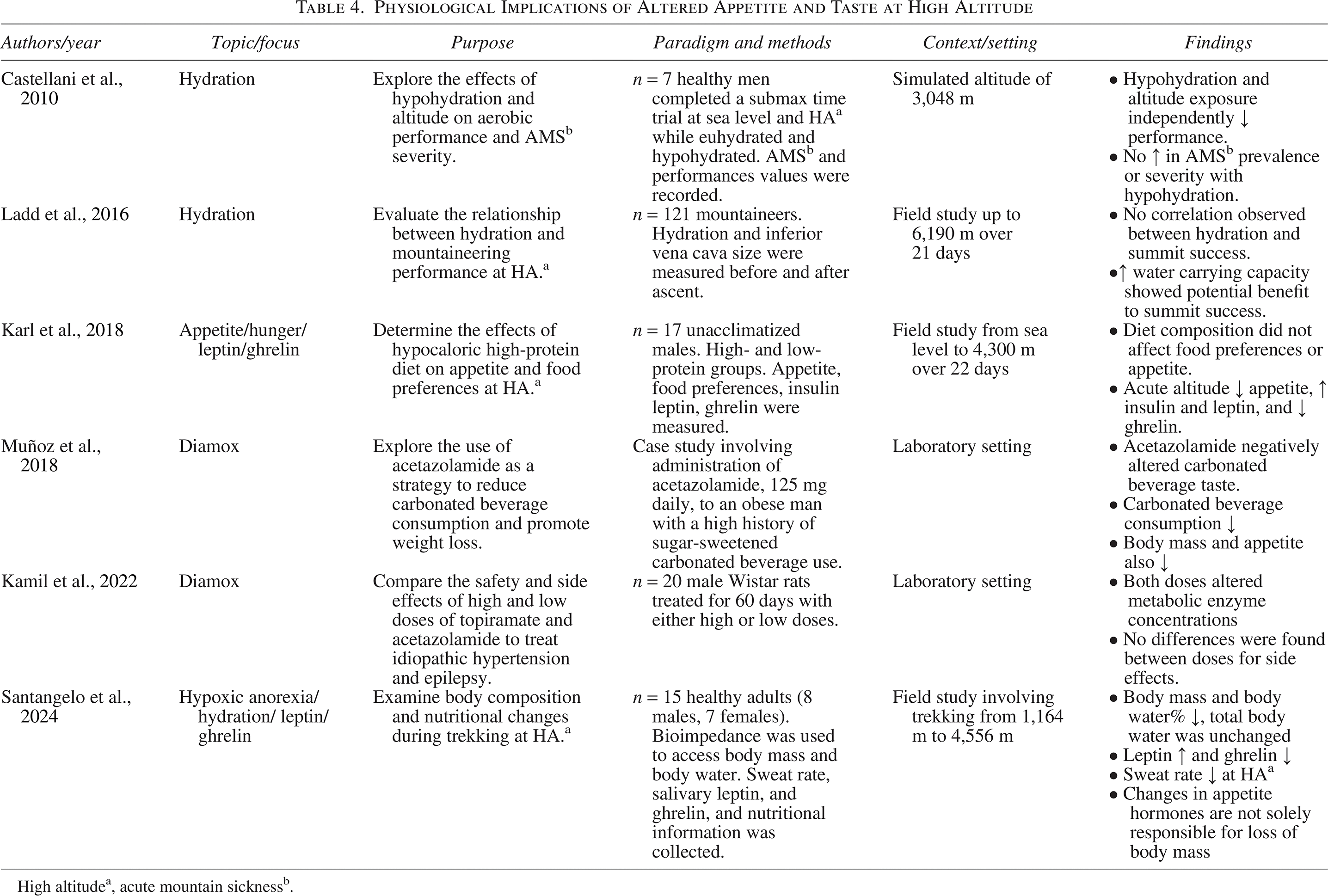
High altitudea, acute mountain sicknessb.
Altitude, Oxygen, and Physiological Responses
Increasing altitude results in a linear decrease in barometric pressure for the first 4,000–5,000 m. After this, the pressure falls exponentially until the earth’s atmosphere is reached. However, elevational definitions are somewhat arbitrary since barometric pressure is also affected by latitude, time, and weather. Moderate altitude is typically defined as 1,500–2,500 m, high altitude as 2,500–4,500 m, very high altitude as 4,500–5,500 m, and extreme altitude is anything above 5,500 m (for reference, Pike’s Peak is 4,302 m, and Mt. Everest is 8,849 m) (Levine and Stray-Gundersen, 1997; Schobersberger et al., 2010; Matu et al., 2017b). Regardless of altitude, the percentage of O2 in the atmosphere is approximately 21%. However, the decrease in atmospheric pressure results in a lower PO2 or, practically speaking, fewer O2 molecules per volume of air. This drop in inspired O2 is also roughly linear until around 5,500 m with a value of 50% less available oxygen than at sea level. Terrestrially, humans can only travel to 8,900 m, the height of Mount Everest. However, with advances in aviation, more people are experiencing short-term exposure to higher altitudes.
Exposure to low PO2 and atmospheric pressure reduces the driving force for oxygen diffusion in the lungs to the blood and lowers levels of arterial oxygen saturation (SaO2). At sea level, atmospheric pressure results in an SaO2 of 97%–99%. Unlike the linear drop in PO2 seen between O2 and barometric pressure, SaO2 falls exponentially, resulting in progressively more severe hypoxia as altitude increases. Although there are slight variances in barometric pressure worldwide, SaO2 can be calculated with some degree of accuracy. This combination of physiology and environment means that individuals living in parts of Asia and South America, primarily Sherpa and Andean, must survive with roughly half the available oxygen compared with those at sea level. This requires considerable physiological adaptations for these cultures to thrive and survive.
To some degree, acute physiological responses to altitude occur in everyone traveling to higher elevations. At sea level, ventilation is driven by the production of carbon dioxide. An increase in cellular respiration from an increased energy demand will result in an increase in carbon dioxide and then an increase in ventilation. At altitude, however, ventilation is driven by the lack of available oxygen and is defined as the hypoxic ventilatory response (HVR) (Powell et al., 1998; Perry, Jonz and Gilmour, 2009). The HVR is seen in two phases: acute and short term. Carotid bodies initiate the increase in ventilation rate and depth for the first five minutes. This is followed by a slow decline over the next 15 minutes. This cyclical rise and fall in ventilation continues over the next 24 hours if hypoxia is sustained after ventilation normalizes (Powell et al., 1998). The HVR can be viewed as the base of the “hypoxic response pyramid,” effectively supporting changes in blood flow, blood compensation, and the central nervous system. Eventually, homeostasis (adaptation) is reached over time.
Chronic HVR is a long-term response to hypoxic environments typically seen in individuals who have adapted to high altitudes (Duffin, 2007). They experience increased angiogenesis, red blood cells, and a shift toward anaerobic metabolism. This adaptation is further supported by the increase in hypoxic inducible factor (HIF-1). HIF-1 is a complex made up of HIF-1α and HIF-1β. Regulation of HIF-1 mainly involves activation and stabilization of HIF-1α as HIF-1β is usually present in cells in a stable form. Under normoxic conditions, HIF-1α is constantly produced and hydroxylated by prolyl hydrolase domain enzymes (PHDs). This hydroxylation marks HIF-1α for degradation by the von Hippel–Lindau protein, ultimately keeping overall levels of HIF-1α low. Under hypoxic conditions, PHD activity decreases as they require oxygen to function, allowing HIF-1α to accumulate and translocate to the nucleus, where it dimerizes with HIF-1β. The HIF-1 complex then binds to hypoxia-responsive elements in the DNA and activates various genes that allow for adaptation to low O2 environments. Genes activated by HIF-1 encode proteins that promote angiogenesis, increase hemoglobin production, enhance the efficiency of O2 transport, and alter metabolism. Chronic exposure to a hypobaric environment can lead to polycythemia and hypoxemia from the overactivation of HIF-1 and HVR. Typically occurring only in populations that live above 2,500 m, the chronic state of hypoxemia results in individuals suffering from CMS, otherwise known as Monge’s disease (Monge and Whittembury, 1976; Powell et al., 1998; Duffin, 2007; Perry, Jonz and Gilmour, 2009).
The link between increased performance at great heights and hypoxic cognitive impairment is more likely due to a blunted sensation of fatigue and pain. Studies on the relationship between pain, fatigue, and hypoxia in humans are scarce. There has been evidence of decreased pain sensation and increased opioid receptors in rats that have experienced chronic hypoxia (Wu et al., 2015), and cognitive studies have shown that climbers and mountaineers show an inflated sense of ability after returning to sea level (Petiet et al., 1988).
Hypoxia elicits divergent circulatory responses systemically and in the pulmonary vasculature. Acutely, systemic vasodilation facilitates the distribution of available oxygen to the skeletal muscle and vital organs. Conversely, vasoconstriction in the pulmonary vasculature exhibits an opposing hemodynamic effect believed to be a response to match the ventilation–perfusion rates (Ainslie and Subudhi, 2014; Wang et al., 2018). Acute pulmonary vasoconstriction ultimately results in pulmonary hypertension, the primary cause of HAPE (Bärtsch et al., 2005). Unlike other organ systems, the heart performs well at altitude. During hypoxic conditions, heart rate and blood pressure elevate to meet the oxygen demands of skeletal muscles and organs. When physical exertion increases, there is a significant rise in cardiac output. With adequate acclimatization, these responses return to prehypoxic levels. Resting heart rate increases moderately and is accompanied by a concomitant reduction in stroke volume (Palatini et al., 1991; Imray et al., 2010; Ke et al., 2017).
Negative Impacts of Hypoxia
The previously reviewed physiological responses play a role in developing HAH, AMS, HACE, HAPE, and CMS. It was once assumed that altitude illnesses represented a spectrum of diseases with increasing severity. However, each has distinct pathologies, requiring multiple medical treatments for patients (Turner et al., 2021). AMS tends to be the primary illness experienced by most when rapidly traveling to high altitudes or beyond their current acclimatization level. As AMS is guaranteed to occur to some degree, this condition is a limiting factor in work and recreation and is often confused with other typical illnesses such as the common flu or cold (Burtscher et al., 2008). This makes a diagnosis of AMS difficult. Once symptoms have progressed enough to impair daily activities, the illness likely has progressed to moderate or severe levels (Imray et al., 2010; Roach et al., 2018). Regardless of the severity, an HAH must be present, as per the Lake Louise Questionnaire (Roach et al., 2018) diagnostic criteria, in combination with other symptoms such as dyspnea, dizziness/lightheadedness, loss of appetite/digestive problems, and fatigue. Progression of AMS can occur within hours or days and depends on how fast and high one travels. Physical exertion, dehydration, and poor nutrition can exacerbate symptoms. The definitive treatment for AMS is descending to a lower altitude, administration of supplementation O2, or using a field hyperbaric chamber such as a Gamow bag. Acetazolamide (Diamox) can be used prophylactically to help prevent or lessen symptoms of AMS. When combined with proper acclimatization techniques, AMS generally resolves and has little to no lasting negative impacts (Hackett et al., 1981; Toussaint et al., 2021).
More severe, high-altitude illnesses can sometimes develop if hypobaric-induced physiological changes go untreated. HAPE is caused by an extended bout of pulmonary hypertension resulting in fluid leakage from the bloodstream into the lungs and alveoli. There is no definitive link between HAPE development and AMS presence. However, it is worth noting that most HAPE cases also exhibit AMS symptoms. HACE represents the potentially most severe form of high-altitude illness. While HAPE may lead to more rapid fatality, the early indicators and symptoms of HACE are similar to those of AMS and may go unnoticed. This may result in HACE not being diagnosed until severe manifestations, such as hallucinations, occur. Previously believed to be a continuum of severe AMS, recent scientific findings have prompted the differentiation of these two conditions based on distinct pathophysiological mechanisms (Hackett et al., 1981; Turner et al., 2021). High-altitude cerebral edema shows a similar etiology to idiopathic intracranial hypertension. However, our understanding of these pathologies remains limited due to the challenges associated with conducting field studies and the low occurrence of cases (Wall, 2017). Treatment for HAPE and HACE is similar to AMS, with the addition of dexamethasone or nifedipine to reduce acute swelling. If not treated, HAPE and HACE are life-threatening.
While research has been done on the recognition and treatment of the various altitude-related illnesses, less attention has been paid to other, nonlife-threatening symptoms associated with travel to high altitude, including loss of taste and appetite, which result in decreased consumption while at altitude. Given the difficulty of maintaining a neutral energy balance at altitude, examining the factors that may impact an individual’s ability to consume enough calories to maintain body weight and perform is essential (Berryman et al., 2018). The degree to which diminished hedonics affects acclimatization is yet to be studied partly because the correlation between taste and altitude has been minimally investigated. Anecdotally, however, there are many mountaineering reports of successful summits, group rescues, and self-rescues supported not only by calorically dense food but also food selections that individuals preferred, lending to the idea that taste may have more to do with a performance at altitude than previously thought.
Effect of High Altitude on Taste and Food Preferences
Taste perception is a complex process that involves multiple physiological and psychological factors. The taste receptor cells on the tongue, roof of the mouth, and throat detect six basic tastes as follows: sweet, sour, salty, bitter, umami (savory), and oleogustus (fat). Papillae comprise multiple taste receptor cells that alternate with supporting cells and resemble “bumps” on the tongue, commonly called taste buds. Papillae can be classified into three distinct subtypes. Fungiform papillae are mushroom-shaped and situated on the front and sides of the tongue, foliate papillae are leaf-shaped and located on the back and sides of the tongue, and circumvallate papillae are circular-shaped and located on the back of the tongue (Davydova et al., 2017). The arrangement of papillae facilitates comprehensive and varied taste perception. Within each taste bud are approximately 50 to 150 taste receptor cells equipped with specialized proteins that preferentially respond to distinct types of molecules, such as sodium chloride (salty taste) and monosodium glutamate (umami taste).
Fat taste detection in humans is closely associated with specific receptors and genetic factors. Liu et al. (2018) highlighted the role of the CD36 receptor, which is particularly sensitive to fatty acids. Variants of this receptor, such as the single-nucleotide polymorphism rs1761667, have been linked to variations in fatty acid sensitivity among individuals. This suggests a genetic basis for the perception of fat taste, which can vary widely between individuals due to differences in receptor expression level (Ackroff and Sclafani, 2014; Liu et al., 2018). Fat perception involves more than taste; it also includes responses from other sensory systems such as smell and chemesthesis. At lower concentrations, fatty acids may not be perceived as having a specific taste quality. Still, as concentrations increase, they become recognizable and can even be unpleasant if they reach suprathreshold levels (Galindo et al., 2012; Keast and Costanzo, 2015).
Once the taste receptors on the tongue are activated, chemical signals are converted to electrical signals and conducted by cranial nerves to the brain stem, where they are then processed and relayed to the gustatory cortex in the temporal lobe. The gustatory cortex processes these signals, giving rise to taste perception. Taste perception is also influenced by other sensory modalities such as olfaction, tactile sensations, and vision. Notably, the sense of smell significantly influences taste perception, as the neural pathways responsible for olfaction and gustation are closely interconnected within the brain (Boesveldt and De Graaf, 2017). In addition, texture and temperature can also affect taste perception. Due to neural plasticity, the brain’s ability to change and adapt in response to these cross-modal interactions can also influence taste perception. For example, repeated exposure to a particular taste can change the neural connections in the brain, leading to an increased or decreased perception of that taste over time (Gillan, 1984; Boesveldt and De Graaf, 2017). This phenomenon, known as taste adaptation, is thought to occur for taste receptors to maintain sensitivity to new or different tastes. Understanding the underlying mechanisms of taste perception can help inform interventions to improve taste perception in those with taste and smell disorders and promote healthy eating habits.
Recent research has also revealed that the physiology of taste perception is more complex than previously thought. Taste receptors are located on the tongue, gastrointestinal tract, and other organs (Young et al., 2009; Gerspach et al., 2011; Foster et al., 2013; Laffitte et al., 2014). In addition, taste receptors have been shown to affect a range of physiological processes beyond taste perception, including immune function, metabolism, and the detection of toxins (Harmon et al., 2021). Several factors, including genetics, age, gender, and the environment, directly influence taste and ultimately impact hunger and appetite. The physiology of taste perception involves a complex interplay of cellular, molecular, and neural processes, and recent research has revealed that taste receptors are involved in a wide range of physiological processes beyond taste perception. Further research is needed to fully elucidate the underlying mechanisms of taste perception and its correlation to hunger and healthy eating habits.
Few studies have examined the impact of high altitude and hypoxia on taste or olfaction. One of the first studies occurred in 1972 when six college-aged women were recruited to examine taste at various altitudes (Maga and Lorenz, 1972). Researchers sought to investigate threshold intensities of sweet, salt, bitter, and sour. After several preliminary threshold taste tests, participants entered a reinforced chamber pressurized initially to sea level and then to 3,048 m. They were instructed to sample concentrations of each taste and to record when they could detect each one. Since all participants had been acclimatized to 1,524 m, the taste concentrations and altitudes of the solution were randomized into two testing groups. The four basic tastes were affected from sea level to 1,524 m but unaltered from 1,524 to 3,048 m, showing to some degree that taste sensation is affected by a decrease in barometric pressure (Maga and Lorenz, 1972). Researchers did not speculate why the participants’ taste was altered, nor did they complete any follow-up studies. This initial research laid the groundwork for subsequent investigations into the impact of altitude exposure on taste perception. However, it is essential to note that much of the existing research on taste and hypoxia predominantly focuses on the effects experienced by air travel passengers rather than individuals working or engaging in recreational activities at high altitudes. During that period, the priority was to provide passengers with enjoyable food rather than catering to the taste preferences of mountaineering expeditions or military operations. It should be acknowledged that most civilian airplanes are depressurized to a simulated altitude of 2,450 m, limiting the generalizability of the findings to higher altitudes or situations involving physical exertion. It was approximately two decades before investigations were undertaken into the altered taste perception and hedonic experiences of individuals living temporarily at high altitudes.
The sense of taste is critical for food selection and enjoyment, regulating food intake, and maintaining energy balance. At high altitudes, various factors can contribute to alterations in taste perception, leading to challenges in maintaining proper nutrition and energy balance. One most commonly reported effects of hypobaria on taste perception is reduced sensitivity to tastes (such as sweet, sour, salty, and bitter) (Maga and Lorenz, 1972; Singh et al., 1997). The exact mechanisms behind these changes are yet to be fully understood, but several possibilities have been suggested. The lower atmospheric pressure at altitude may lead to changes in salivary flow, which can subsequently impact taste bud function. Reduced salivary flow from the cold and dry climate can result in a dry mouth, making it more difficult for taste compounds to dissolve and interact with taste receptors (Muñoz-González et al., 2018; Liu et al., 2021). When combined with hypoxic-induced dehydration and further dehydration from physical activity, the reduced salivary output can make food consumption difficult, resulting in individuals avoiding calorically dense solid food for liquid alternatives (Westerterp and Kayser, 2006; Aeberli et al., 2013; Berryman et al., 2018).
High altitude can cause nasal congestion due to the dry and cold environment, impairing the sense of smell (Huppertz et al., 2018; Mazzatenta et al., 2022). Since the senses of taste and smell are closely linked (Weiffenbach and Bartoshuk, 1992), nasal congestion can further exacerbate the alterations in taste perception experienced at high altitudes. Lastly, the stress response to high altitude, including releasing stress hormones such as cortisol, can also impact taste perception. Acutely, poststress cortisol levels reduce sensitivity to salt and sour sensations, and acute stress, in general, can attenuate sweet taste perception (Al’Absi et al., 2012). However, overall hormonal regulation during ascent and acclimatization to high altitude is highly complex and needs further evaluation. The exact relationship between stress and taste perception is also complex and not fully understood, but it is thought that stress hormones, beyond cortisol, may modulate taste sensitivity and preferences. Von Wolff et al. (2018) and then Keenan et al. (2020) showed marked changes in both secretion and pulsatile frequency during ascent to high altitude for stress-related hormones such as prolactin, thyroid hormone, and cortisol, with levels returning to prehypoxic values upon successful acclimatization (Von Wolff et al., 2018; Keenan et al., 2020). Depending on the degree of altitude exposure, stress response and hormonal release can increase, possibly exacerbating the negative impact on taste. Similar to cortisol, increased thyroid hormone levels have been shown to increase taste thresholds for salt, bitter, and, to some degree, sour (Bhatia et al., 1991; Sebastian et al., 2022). Hormonal processing is likely further impacted by the increase of HIF-1 as HIF-1 is a known suppressor of pituitary hormone synthesis. It is important to note that individuals may experience varying degrees of taste perception changes at high altitudes. Some individuals may be more susceptible to these changes due to genetic differences in taste receptors or other factors related to their physiological responses to altitude.
Effect of High Altitude on Hunger and Appetite
Appetite and hunger are complex processes that involve the interaction of multiple hormones and neural pathways. Two critical hormones involved in appetite are ghrelin and leptin. Ghrelin is produced in the stomach and generally stimulates appetite, while leptin is produced by fat cells and signals the brain to decrease appetite. Ghrelin levels rise before meals and fall after eating (Williams and Mobarhan, 2003). Ghrelin acts on the hypothalamus, which plays a crucial role in regulating hunger and satiety, along with stimulating the release of neuropeptide Y and agouti-related protein, which increase appetite and decrease energy expenditure (Kas et al., 2005). Ghrelin also inhibits the release of proopiomelanocortin (POMC) and cocaine- and amphetamine-regulated transcript, which reduces appetite and increases energy expenditure (Vrang et al., 2002; Younes-Rapozo et al., 2012). Leptin levels, on the contrary, rise as fat stores increase and decrease as fat stores decrease. Leptin acts on the hypothalamus as well, inhibiting the release of neuropeptide Y and agouti-related protein and stimulating the release of POMC and amphetamine-regulated transcript. Leptin also activates the sympathetic nervous system, which increases energy expenditure (Vrang et al., 2002; Izadi et al., 2014). Ghrelin has been shown to increase the secretion of leptin, which may be a mechanism for maintaining energy balance. Leptin, in turn, has been shown to inhibit the release of ghrelin, which may be a mechanism to promote satiety. However, the exact mechanisms by which these hormones interact are still being studied (Williams and Mobarhan, 2003).
Diverse physiological and environmental factors can perturb the intrinsic hunger experienced at high altitudes, thereby eliciting alteration in appetite and subsequent food consumption. One possible explanation is the alteration of hormone levels, such as ghrelin and leptin, which directly impact hunger. Hunger, in turn, influences food consumption and energy balance. As mentioned earlier, the hormone ghrelin plays a central role in stimulating hunger, and its levels have been found to fluctuate greatly at high altitudes (Williams and Mobarhan, 2003). Ghrelin is present in humans in acylated and unacylated forms. Desacyl-ghrelin, commonly called unacylated ghrelin, had long been considered an inert degradation of acylated ghrelin. However, recent studies have shown unacylated ghrelin to have its own receptors and play a role in inhibiting acylated ghrelin, inducing a negative energy balance by decreasing food intake and delaying gastric emptying. Acylated ghrelin is the dominant form found in humans and has the inverse effect of unacylated ghrelin. It promotes hypertrophy in cytosolic lipid droplets in adipose tissue and the liver, ultimately facilitating a positive energy balance.
Both are 28 amino acid peptides primarily secreted by the stomach and small intestine (Asakawa et al., 2005; Toshinai et al., 2006). Prior investigations have demonstrated a decline in acylated and salivary ghrelin levels with increasing altitude (Shukla et al., 2005; Riedl et al., 2012; Matu et al., 2017c; Santangelo et al., 2024). Santangelo et al. (2024) examined 12 healthy adults (6 males, 6 females) during a trek to 4,556 m over 4 days. Food intake was recorded, and energy intake was calculated. Body mass, body water, sweat rate, hydration, and salivary leptin and ghrelin were measured before, during, and after the sojourn. Significant alterations were observed in body mass and body water percentage. However, the absolute water level remained unchanged, indicating that the reduction in body mass was not specifically due to water loss. A reduction in sweat rate was also observed at higher altitudes. Lastly, salivary leptin increased and ghrelin decreased, ultimately leading to dysregulated appetites that likely contributed to reduced body mass. To date, no studies have reported on unacylated ghrelin levels (Riedl et al., 2012; Matu et al., 2017c, 2017a; Santangelo et al., 2024). Hypoxia in and of itself can acutely blunt ghrelin levels due to systemic blood redistribution and lower liver blood flow, decreasing ghrelin acylation (Debevec, 2017). At the same time, other studies have reported no discernible alterations compared with sea level, and a minority have observed an elevation (Shukla et al., 2005; Riedl et al., 2012; Wasse et al., 2012; Matu et al., 2017c, 2017a).
Hypoxia is an unavoidable feature of high-altitude environments and has been shown to inhibit hunger and appetite (Butterfield et al., 1992; Reynolds et al., 1999; Westerterp and Kayser, 2006; Wing-Gaia, 2014; Berryman et al., 2018). Some studies suggest hypoxia may suppress appetite by altering the central nervous system’s sensitivity to ghrelin and other appetite-regulating hormones (Hornbein et al., 1989; Strader and Woods, 2005; Schwartz, 2006). This may explain how hunger and appetite could be impacted without changes in circulating ghrelin. Interestingly, the link between reduced appetite and ghrelin is not limited to hypobaric exposure. Normobaric hypoxia also reduces ghrelin levels compared with sea level when the hypoxic gas composition is altitude-matched (Chroboczek et al., 2021). Conversely, leptin tends to exhibit an inverse relationship to ghrelin under hypoxic conditions (Tschöp et al., 1998; Shukla et al., 2005). Since HIF-1α is expressed in POMC neurons, high levels of HIF-1 in hypoxic environments may influence how these neurons respond to leptin, as secreted leptin will ultimately bind to POMC neurons in the hypothalamus (Gaspar and Velloso, 2018). However, similar to the heterogeneous reports on ghrelin level changes at altitude, studies indicate varied patterns of leptin levels, including increases, decreases, and no change (Vats et al., 2004; Zaccaria et al., 2004). Based on these findings, ghrelin and leptin are not solely responsible for altered hunger under hypoxic conditions. The suppression of hunger, increased resting metabolic rate, and increased energy expenditure at altitude all contribute to the hypoxia-induced anorexia phenomenon (Berryman et al., 2018).
Appetite is also changed at altitude. Although appetite and hunger regulation are related, they are not the same. Hunger is a biological or physiological need to eat driven by changes in blood metabolites. Whereas appetite is a psychological drive to eat that can be altered by hunger, it can also occur without hunger. As discussed earlier, critical appetite-regulating hormones, such as ghrelin and leptin, are affected by high altitudes (Tschöp et al., 1998; Shukla et al., 2005). In addition to these hormones, other appetite-regulating factors, such as cholecystokinin (CCK) and peptide YY (PYY), may also alter appetite at high altitudes (Matu et al., 2017c). CCK is a peptide hormone produced in the small intestine that contributes to satiety by promoting the release of digestive enzymes and bile, delaying gastric emptying, and acting on the central nervous system. Studies have reported increased CCK levels at high altitudes, possibly contributing to reduced appetite and food intake (Matu et al., 2017c). PYY is another peptide hormone released from the gastrointestinal tract in response to food intake, and it plays a role in promoting satiety. Wasse et al. (2012) showed that PYY levels were elevated at high altitudes, potentially contributing to appetite suppression in individuals exposed to high-altitude conditions (Wasse et al., 2012).
Factors such as dehydration and gastrointestinal discomfort at altitude can further impact appetite and food intake. These factors, combined with the changes in appetite and hormonal regulation of hunger, make maintaining energy balance particularly challenging for individuals at high altitudes. Whether there is a drive to eat at altitude, the combined effects of hypoxia and overall fatigue significantly impact individuals’ desire to eat.
The impact of altitude on appetite is a multifaceted phenomenon encompassing intricate interactions among diverse physiological, hormonal, and environmental factors. These modifications in appetite can engender a diminished desire for food and subsequent reductions in dietary intake, presenting significant obstacles to sustaining energy balance and fostering optimal well-being among individuals exposed to high-altitude circumstances (Westerterp-Plantenga et al., 1999; Berryman et al., 2018). The consistently diminished appetite reported in these previous studies is likely due to the cumulative effects of taste disruptions, hypoxia, and additional stressors. Enhancing our comprehension of the elements influencing changes in hunger at high altitudes and formulating strategies to mitigate their effects are paramount to optimizing health and performance in high-altitude environments.
Implications for Nutrition and Health at High Altitude
Given the challenges of altitude-induced changes in taste perception, hunger, and appetite, exploring potential strategies for mitigating these effects is essential. Addressing these issues can help maintain energy balance, optimize nutritional status, and improve overall performance at high altitudes. As mentioned earlier, evidence suggests that the negative energy balance induced by altitude primarily stems from the resultant anorexia rather than solely from limited food availability or other altitude-associated challenges such as load carriage management, food preparation constraints, water scarcity, and environmental hazards. The precise altitude threshold where anorexia becomes evident is not known and may vary among individuals. As these past studies have shown, hypoxic anorexia may be unavoidable even under the best circumstances, regardless of how high one goes (Matu et al., 2017a; Berryman et al., 2018; Karl et al., 2018; Santangelo et al., 2024). Major & Doucet (2004) and Matu et al. (2017a) both reported a threshold between 2,000 and 2,500 m. However, Dünnwald et al. (2019) observed a loss of body mass altitudes as low as 1,500 m. Santangelo et al. (2024) also saw a reduction of body mass over 4 days coupled with changes in appetite hormones but surmised that altered appetite and satiation are not directly linked to hypoxic anorexia. Hence, it can be surmised that the interplay of one or multiple factors could potentially restrict energy intake through direct anorexigenic effects, challenges associated with food preparation and consumption, altered taste preferences, or any of these combined (Major and Doucet, 2004; Matu et al., 2017c; Dünnwald et al., 2019; Santangelo et al., 2024).
One approach to counteract the altered taste perception at high altitudes is to enhance the flavor of foods by using more intense seasonings and spices (Singh et al., 2000, 1997; Al’Absi et al., 2012; Boesveldt and De Graaf, 2017). This approach can improve the sensory appeal of meals, stimulate food consumption, and alleviate the appetite-suppressing effects associated with high-altitude exposure. In addition, opting for energy-dense and nutrient-rich food choices can effectively fulfill individuals’ energy and nutritional demands at high altitudes (Shukla et al., 2005; Harmon et al., 2021). This may involve choosing foods high in healthy fats, complex carbohydrates, and high-quality proteins and ensuring an adequate intake of vitamins and minerals. Singh et al. (1997) completed one of the only studies examining the effect of hypoxic stress on hedonics. Investigators showed that providing a highly palatable carbohydrate-rich diet could attenuate hypoxic anorexia and the resulting body mass loss (Singh et al., 1997). Other studies have also demonstrated a switch in food preference to carbohydrates. Matu et al. (2017b) showed when given a high-fat breakfast, appetite suppression was tempered in high-altitude sojourners, particularly during exercise (Matu et al., 2017a). Despite subjects maintaining their food consumption at altitude, there was a measurable negative impact on body mass.
As discussed earlier, oral fatty acid sensitivity can be individually unique. Individuals less sensitive to fatty acids tend to consume less fat and have lower energy intake overall. Conversely, more sensitive people may need to regulate their fat intake more effectively, leading to higher fat consumption and larger daily calorie consumption. To date, no studies have completely mitigated the effects of hypoxic anorexia. Drawing from research on populations residing at high altitudes and observations of Himalayan mountaineers, the energy balance can eventually be reestablished. The pivotal inquiry remains: how much sustained exposure to altitude is required to negate anorexic effects? Furthermore, what other physiological ramifications may arise from extended altitude exposure? These questions are yet to be elucidated and warrant further investigation in future studies (Kayser, 1992; Beall, 2002; Shukla et al., 2005; Aeberli et al., 2013).
These previous studies show that the total amount of calories one consumes is more important than the proportion of macronutrients in one’s diet. As a result, future research should concentrate on implementing ways to recognize those with an innate inclination toward consuming foods high in calories and fats. By identifying individuals who generally avoid fats, it would be possible to provide them with improved recommendations for preventing an excessive caloric deficit, particularly during high-altitude work or recreational activities (Keast and Costanzo, 2015; Matu et al., 2017a).
Irregular meal timing can be an issue due to the nature of the environment, the availability of adequate camping locations, and the duration of the overall exposure time. Consuming smaller, more frequent meals throughout the day, rather than fewer large meals, could help maintain the energy balance and promote appetite at high altitudes (Westerterp-Plantenga et al., 1999). This approach can also aid in preventing gastrointestinal discomfort, which can further suppress appetite. Along with meal frequency, euhydration is critical at high altitudes, as hypohydration has the potential to dampen appetite and give rise to gastrointestinal complications. Ensuring sufficient fluid consumption is vital for upholding optimal bodily functions and potentially alleviating the impact of altitude on hunger and appetite (Castellani et al., 2010; Ladd et al., 2016).
When possible, proper acclimatization to high altitude can minimize the physiological challenges associated with altitude exposure, including the impact on taste perception and hunger. Gradual ascent and allowing time for the body to adjust to the new environment can help reduce the severity of these effects (Roach and Hackett, 2001; Mallet et al., 2023). Some evidence suggests that medications such as Diamox can also impact taste perception and appetite (Kamil et al., 2022). Diamox aids in acclimatization by inhibiting carbonic anhydrase and facilitating the excretion of bicarbonate, effectively counteracting the respiratory alkalosis that occurs at altitude (Becker, 1955). Nevertheless, the diuretic function of Diamox poses challenges in maintaining optimal hydration levels, compromising euhydration.
A prominent side effect of carbonic anhydrase inhibitors is appetite suppression. Studies have indicated that Diamox can induce metabolic alterations at high and low concentrations at altitude (Muñoz et al., 2018; Kamil et al., 2022). In some cases, nutritional supplementation may be necessary to ensure adequate intake of essential nutrients, mainly when food availability and variety are limited (Butterfield et al., 1992; Butterfield, 1999). Supplements should be carefully considered and tailored to individual needs and specific high-altitude conditions. Very few studies have been conducted on the effectiveness of supplementation to mitigate the effects of hypoxic anorexia or enhance performance under hypoxic conditions. One of the primary challenges associated with high-altitude performance is regulating hypoxic acid/base balance. To address this issue, research studies should investigate the effectiveness of various supplements, including iron, nitrate (such as beetroot), sodium bicarbonate, citrate, and beta-alanine. By conducting comprehensive studies on these supplements, we can better understand their potential to enhance performance at high altitudes (Cerretelli and Samaja, 2003; Peeling et al., 2018). The benefits of these supplements have shown positive results at sea level, but their effectiveness at altitude is yet to be revealed (Perut et al., 2020; Grgic et al., 2021; Ostfeld and Hoffman, 2023; Schulte et al., 2024). Several strategies can be used to mitigate the negative impacts of altitude on taste perception and hunger, with the ultimate goal of helping individuals maintain energy balance, optimize nutrition, and improve performance at high altitudes.
Conclusion
This review underscores the importance of evaluating gustation under hypobaric hypoxic conditions. The physiology of hunger and taste perception involves intricate interplays of physiological and psychological factors. Appetite and hunger are complex processes influenced by hormonal and neural pathways. While changes in ghrelin and leptin have been implicated in feelings of hunger and appetite, it is imperative to acknowledge that hypoxic-induced anorexia is not solely attributed to these hormones. Moreover, high-altitude exposure can modify taste perception, characterized by reduced sensitivity to different tastes, impaired salivary flow, nasal congestion, and stress responses. These alterations vary among individuals and can challenge achieving proper nutrition and maintaining energy balance. These variables are significant for individuals operating in alpine environments, such as warfighters and search and rescue personnel. Given the necessity to transport all nutrition supplies, food selection becomes crucial in ensuring energy balance. As previously stated, maintaining the caloric status becomes challenging; ensuring adequate macro and micronutrient intake through enjoyable food choices is critical to mitigating the risk of lean body mass reduction.
As this review has shown, understanding the complex interaction between nutritional balance and exposure to high altitude needs further investigation. Future research should focus on studies that include taste perception and dietary interventions as primary aims, as these outcomes have traditionally been secondary variables. Studies should focus on controlling food intake from a macronutrient perspective, and foods should be tailored to the subjects’ personal preferences to increase the likelihood of maintaining adequate caloric consumption.
Comprehending the underlying physiological mechanisms governing hunger, taste perception, and their interactions with high-altitude exposure is imperative for fostering healthy eating habits and optimizing well-being in such demanding conditions. Further research is needed to fully elucidate these mechanisms and develop targeted interventions to address the distinctive nutritional and sensory challenges of high-altitude environments.
Authors’ Contributions
J.T.M.: Conceptualization (lead), writing—original draft (lead), and writing—review and editing (lead). J.L.T.: Writing—review and editing (supporting). D.H.: Supervision (lead) and writing—review and editing (equal).
Footnotes
Author Disclosure Statement
The authors have no financial conflict of interest.
Funding Information
The author(s) received no financial support for the research, authorship, and/or publication of this article.