
Abstract
Hepatic gene transfer is envisioned as a substitute for protein replacement therapies, many of which are derived from blood products. Thus, the target populations may have a high prevalence of blood-borne pathogens, such as hepatitis C virus (HCV). We sought to determine whether the safety of recombinant adeno-associated virus serotype 2 (rAAV2) would be altered by preexisting HCV infection. Doses of approximately 1 × 1013 vector genomes of an rAAV2-chimpanzee α1-antitrypsin (rAAV2-cAAT) vector were injected into the portal vein of each of three HCV genome-positive (HCV+) chimpanzees and three HCV-negative (HCV−) controls. Acute safety studies were performed up to 90 days after vector administration, along with analyses of the peripheral blood and liver tissue for rAAV2-cAAT genomes. Vector genome copy numbers in blood and liver tissue were similar in both groups. All animals demonstrated increases in liver and muscle enzyme levels after the pretreatment liver biopsy (5 days before vector injection) and after the vector injection. However, HCV+ animals demonstrated a substantially greater rise in aspartate aminotransferase, alanine aminotransferase, and creatinine phosphokinase values than HCV− animals. Histopathology demonstrated abnormal lipid accumulation (steatosis) in the hepatocytes of HCV+ animals, both before and after vector injection. These data indicate an increased susceptibility to subclinical liver toxicity from portal vein injection of rAAV2 in the presence of HCV infection.
Introduction
The only well-established model for HCV infection is in chimpanzees (Bukh, 2004). Several studies have demonstrated that chimpanzees can become chronically infected with HCV. Chronically infected animals demonstrate HCV seropositivity, persistence of HCV genomes in their peripheral blood, and a variety of mild liver abnormalities (Dash et al., 2001; Poduri, 2003; Bukh, 2004; Fernandez et al., 2004; Katayama et al., 2004; Major et al., 2004). Chimpanzees also represent a suitable model to mimic human responses to recombinant viral vectors because they are so closely related to humans genetically. In the current study, the advantages of the chimpanzee model were exploited to address the question of whether HCV infection predisposes to rAAV2-mediated liver toxicity. Groups of HCV+ and HCV− chimpanzees were each injected with doses of 1 × 1013 vector genomes (VG) of an rAAV2-chimpanzee AAT-expressing vector. The chimpanzee AAT gene sequence was used to avoid adaptive immune responses to the transgene product, which could have affected the results. The data presented here suggest an important effect of HCV infection status on the level of liver enzyme elevation after liver-directed rAAV2 vector delivery.
Materials and Methods
Vector preparation
The rAAV-hAAT vector, previously described elsewhere (Xu et al., 2001; Song et al., 2001, 2002), was modified by insertion of the chimpanzee AAT cDNA, which was amplified by polymerase chain reaction (PCR) from a chimpanzee liver cDNA pool, using the same technique previously used by our group to clone the rhesus AAT cDNA (Song et al., 2002).
Vectors were packaged and purified by a standard co-transfection technique in HEK-293 cells followed by conventional chromatographic techniques (Zolotukhin et al., 2002). Cells were harvested and resuspended in buffer, and each sample was prepared by a single pass through a micro-fluidizer after endonuclease (Benzonase; Merck, Indianapolis, IN) treatment. The sample was applied to a heparin affinity column, washed, and eluted and the vector-containing fraction was collected. The eluted sample was adjusted to 1.1 M NaCl via the addition of concentrated sodium chloride and then passed through a hydrophobic interaction column, and the virus-containing flow-through was collected. The sample was diluted, using sterile water for injection, applied to a column containing sulfopropyl cation-exchange resin, and washed, and the virus was eluted.
Animal care approvals and oversight
This study was conducted in accordance with all applicable animal care guidelines at an AAALAC (Association for Assessment and Accreditation of Laboratory Animal Care International)-accredited facility, and was preapproved by the University of Louisiana at Lafayette Institutional Animal Care and Use Committee. All animals were housed with appropriate social stimulation and veterinary oversight.
Portal vein injection under fluoroscopic guidance
The subjects were placed on the procedure table and general anesthesia was induced.
The right hemiabdomen was prepared and draped in a sterile fashion. The liver was localized by both fluoroscopy and ultrasound. Using these modalities, an access site was chosen. Local anesthesia was administered followed by a dermatotomy.
A 21-gauge needle was advanced into the peripheral liver parenchyma. Carbon dioxide was then administered directly into the liver parenchyma to visualize the portal vein. The needle was then advanced into the portal vein and its position was confirmed with iodinated contrast and withdrawal of venous blood. The specimen was delivered into the portal vein followed by a saline flush. The final position of the needle was confirmed with a contrast injection. The needle was removed and hemostasis was achieved by manual compression.
Monitoring of clinical laboratory findings
Blood was drawn by venipuncture at each of the following time points relative to vector injection (defining the vector injection day as day 0): days –7, –5, 0, 1, 7, 14, and 90. At each of these time points, the following assays were performed in compliance with University of Louisiana at Lafayette–New Iberia Research Center standard operating procedures: complete blood counts, serum electrolytes (including sodium, potassium, chloride, bicarbonate, blood urea nitrogen, creatinine, and glucose), liver enzymes (including aspartate aminotransferase [AST], alanine aminotransferase [ALT], alkaline phosphatase [ALP], bilirubin, and γ-glutamyltransferase [GGT]), and creatine phosphokinase (CPK). In addition, blood was drawn on days –7, 14, and 90 for anti-AAV2 and anti-AAT antibodies by enzyme-linked immunosorbent assay (ELISA) and for antigen-specific lymphocyte proliferation against AAV2 and AAT antigens, using published techniques (Brantly et al., 2006).
Liver biopsy procedures, tissue processing, and analysis
Liver biopsies were collected in a sterile manner, using a Jamshidi Menghini 16-gauge liver biopsy needle (Cardinal Health, Dublin, OH). A 3-cm core of liver tissue was collected. Liver biopsies were fixed in 10% neutral buffered for malin and processed and embedded in paraffin. Sections (4 μm) were cut and stained with hematoxylin and eosin. Histopathology review was performed blinded to HCV status and samples were scored for infiltrates (portal tract and parenchymal regions), fibrosis, and apoptosis. The grading scale was as follows: 0, none; 1, mild, focal; 2, mild, multi-focal; 3, moderate, multifocal; 4, severe, multifocal. Macrovesicular and microvesicular steatosis were scored separately by estimating the percentage of positive cells according to the following scale: 0, none; 1, <5%; 2, 5–9%; 3, ≥10%. The presence of stellate cells and hemosiderosis was also noted.
Quantitative real-time PCR of blood and liver tissue
Genomic DNA (gDNA) was extracted from blood and liver tissue according to the manufacturer's protocol (DNeasy tissue kit; Qiagen, Valencia, CA). Resulting DNA concentrations from the extraction procedure were determined with a biophotometer (model 6131; Eppendorf, Hamburg, Germany). One microgram of extracted gDNA was used in all quantitative PCRs according to a previously used protocol (Song et al., 2002) and reaction conditions (recommended by Applied Biosystems, Foster City, CA) included 50 cycles of 94.8°C for 40 sec, 37.8°C for 2 min, 55.8°C for 4 min, and 68.8°C for 30 sec. Primer pairs were designed to the chicken β-actin promoter as described (Donsante et al., 2001) and standard curves were established by spike-in concentrations of a plasmid DNA containing the same promoter (Song et al., 2002). DNA samples were assayed in triplicate. The third replicate was spiked with cytomegalovirus (CMV) enhancer/chicken β-actin DNA at a ratio of 100 copies/μg of gDNA. If at least 40 copies of the spike-in DNA were detected, the DNA sample was considered acceptable for reporting vector DNA copies.
Serum cytokine determination
Assessments of cytokine profiles from the serum were performed with a commercially available multiplex kit (mouse multi-cytokine detection system; Bio-Rad, Hercules, CA) and the Bio-Plex suspension array system (Bio-Rad). Simultaneous measurement of several cytokines was performed. All assays were performed according to the manufacturer's protocols. Cytokine concentrations were determined with Bio-Plex software with four-parameter data analysis. The sensitivity of the assay is less than 10 pg/ml and has a range of 0.2–32,000 pg/ml with an inter- and intraassay coefficient of variation less than 10%.
Results
Portal vein injections and clinical status
Radiographic contrast material was mixed with each dose of 1 × 1013 VG of rAAV2-chimpanzee α1-antitrypsin (rAAV2-cAAT). All portal vein injections were confirmed by fluoroscopic visualization (Fig. 1). All animals tolerated all procedures well, with no clinical evidence of bleeding, hypotension, or respiratory distress. Each animal was monitored clinically for 90 days after vector administration. Animals were then released to participate in other studies.
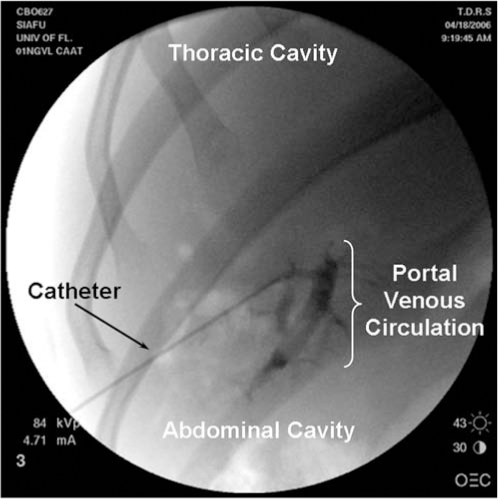
Fluoroscopic delivery of rAAV2-cAAT to the portal vein of chimpanzees. Doses of 1 × 1013 VG of rAAV2-cAAT mixed with nonionic radiographic contrast agent were injected via a transabdominally introduced needle into the portal circulation. The filling of the portal venous system is demonstrated by the fluoroscopic image.
Presence of vector genomes in blood and liver tissue
Vector genome copies were present in the peripheral blood of all animals within the first 7 days after vector injection, but not afterward (Fig. 2A and B). Copy numbers of vector genomes were similar between the two groups regardless of HCV status, with the highest levels at approximately 1 × 104 VG per microgram of cell DNA. Vector genomes were also present in the liver tissue of both groups, again with similar levels between the groups (Fig. 2C and D). However, in the case of liver tissue, vector genomes persisted in some animals at a low level at 90 days after vector injection. Genome copy number peaked in liver tissue at day 7, with values up to the 104 to 105 copies per microgram, and then declined at 90 days in all animals.
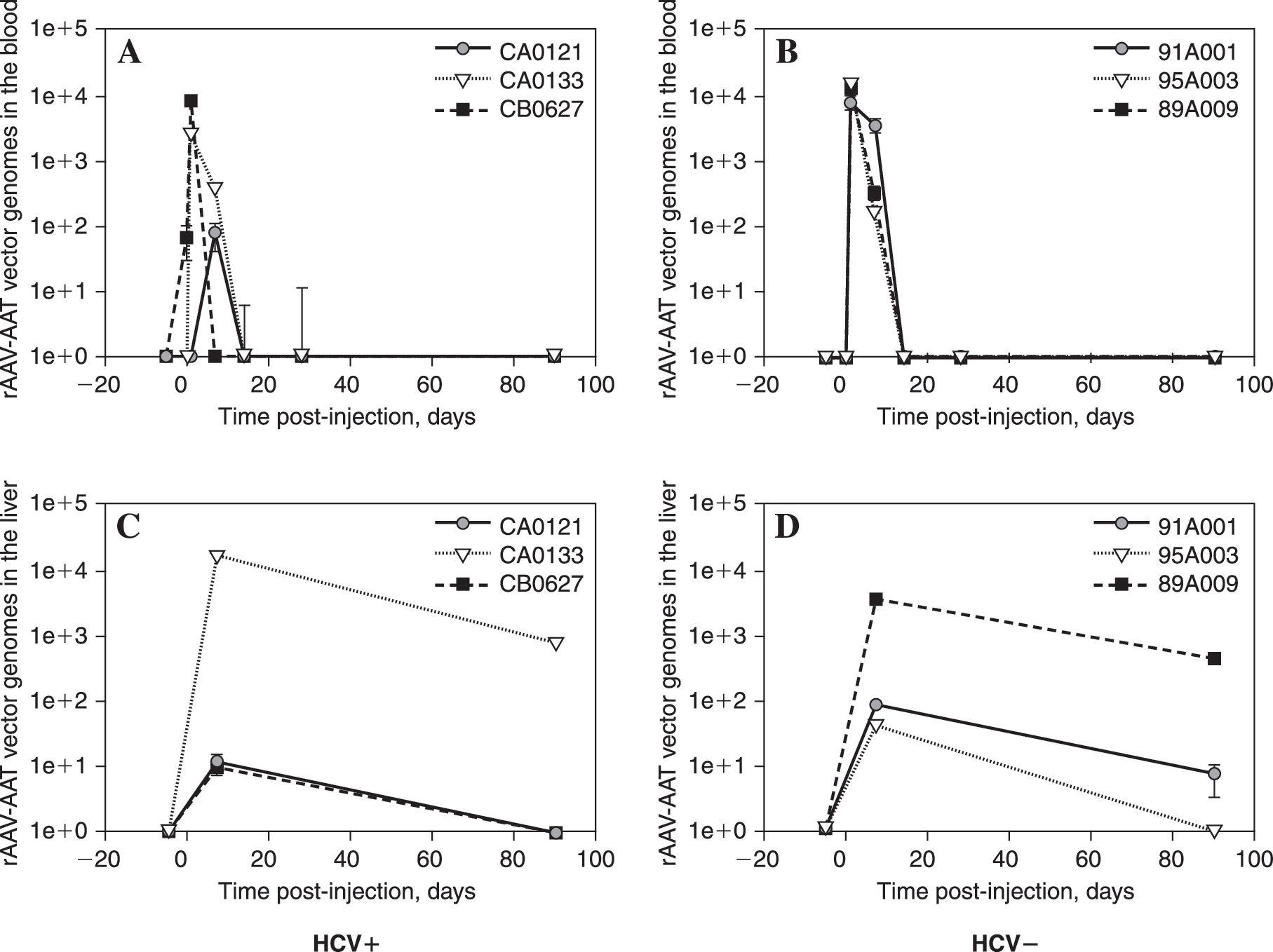
Quantitative PCR indicates the presence of vector genomes within the first 7 days postinjection in the blood and low-grade persistence of vector genomes at 90 days in the liver. DNA was extracted from the peripheral blood and liver biopsy samples and 1 μg of each was analyzed by real-time quantitative PCR for the presence of rAAV2-cAAT vector genomes. Samples were assayed in duplicate and the error bars indicate the range for each time point for each individual animal. The identification of each animal is indicated in the inset legend. (
Elevation of hepatocellular enzymes after liver biopsy or vector injection
The levels of AST increased after both the pretreatment liver biopsy and vector injection in all animals (Fig. 3C and D). The absolute levels of AST and the incremental increases were greater in the HCV+ animals than in the HCV− animals, indicating an increase in susceptibility to hepatocellular injury after liver manipulation regardless of whether or not vector was involved. ALT is a more liver-specific indicator of hepatocellular toxicity. ALT elevations showed a similar pattern, in that elevations were greater in HCV+ animals and occurred both after the day –5 biopsies and after the day 0 vector injections (Fig. 3A and B). Interestingly, HCV− animals showed slight changes in ALT. Two of three HCV+ animals also had increased ALT levels on day 90. Although there is no significant effect of rAAV transduction on HCV infection as determined by TaqMan titers, the two animals with the highest titers were the animals with the sustained ALT elevation (Fig. 3A and G). No animals showed clinical or biochemical evidence of jaundice, coagulopathy, electrolyte disturbance, or other clinically significant toxicity.
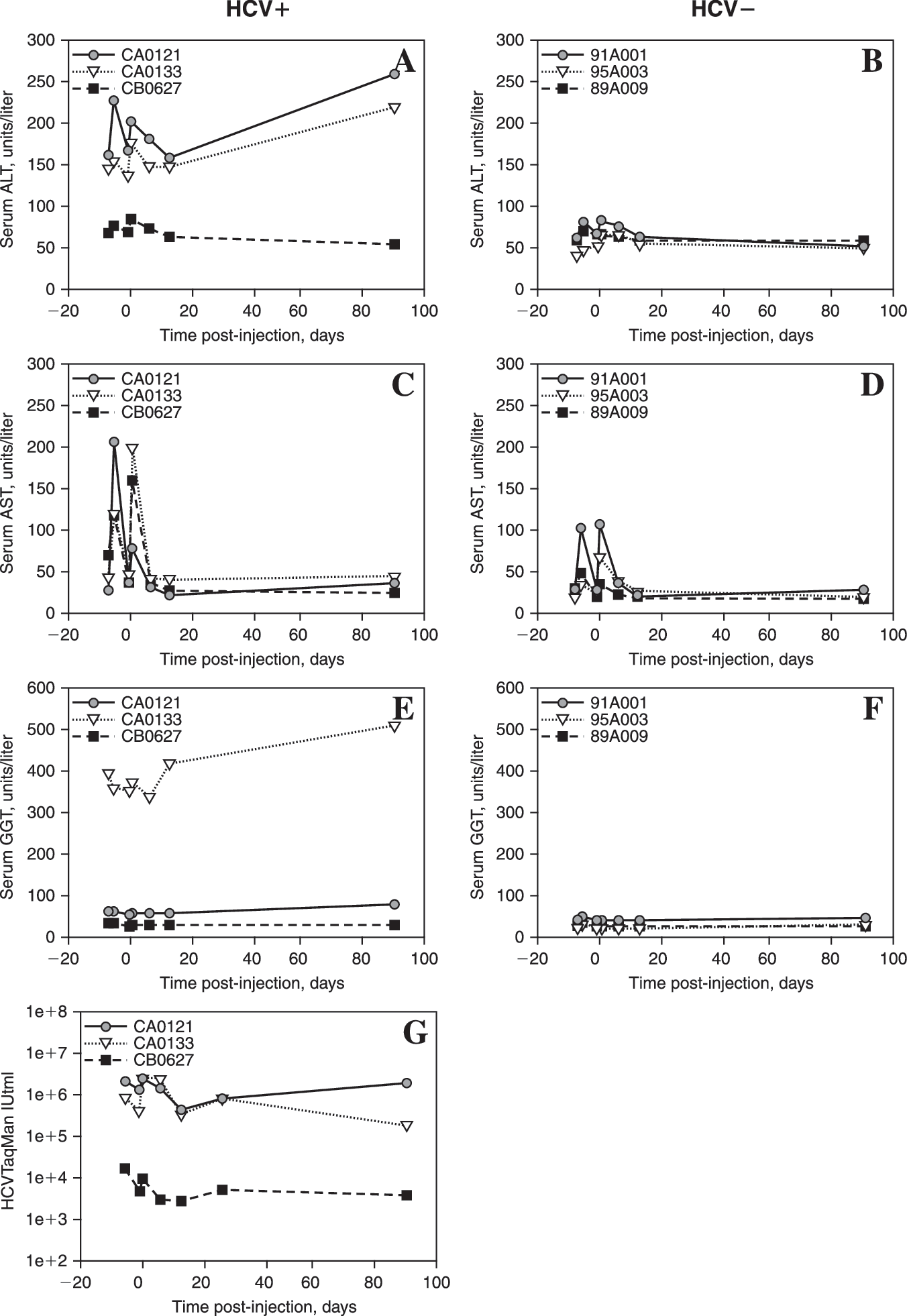
Serum liver enzyme levels after portal vein injection of 1 × 1013 VG of rAAV2-chAAT into HCV+ versus HCV− chimps. (
Other serum enzymes
Enzyme indicators of obstructive liver disease were not affected by the procedures. Serum ALP levels were normal in all animals (data not shown), whereas GGT showed persistent elevation in one HCV+ animal, but no changes in any other animals (Fig. 3E and F). Unexpectedly, CPK, a muscle enzyme, showed a pattern of increase similar to AST and ALT, with elevations noted after both biopsies and portal vein injections and a greater elevation in the HCV+ animals (Fig. 4).
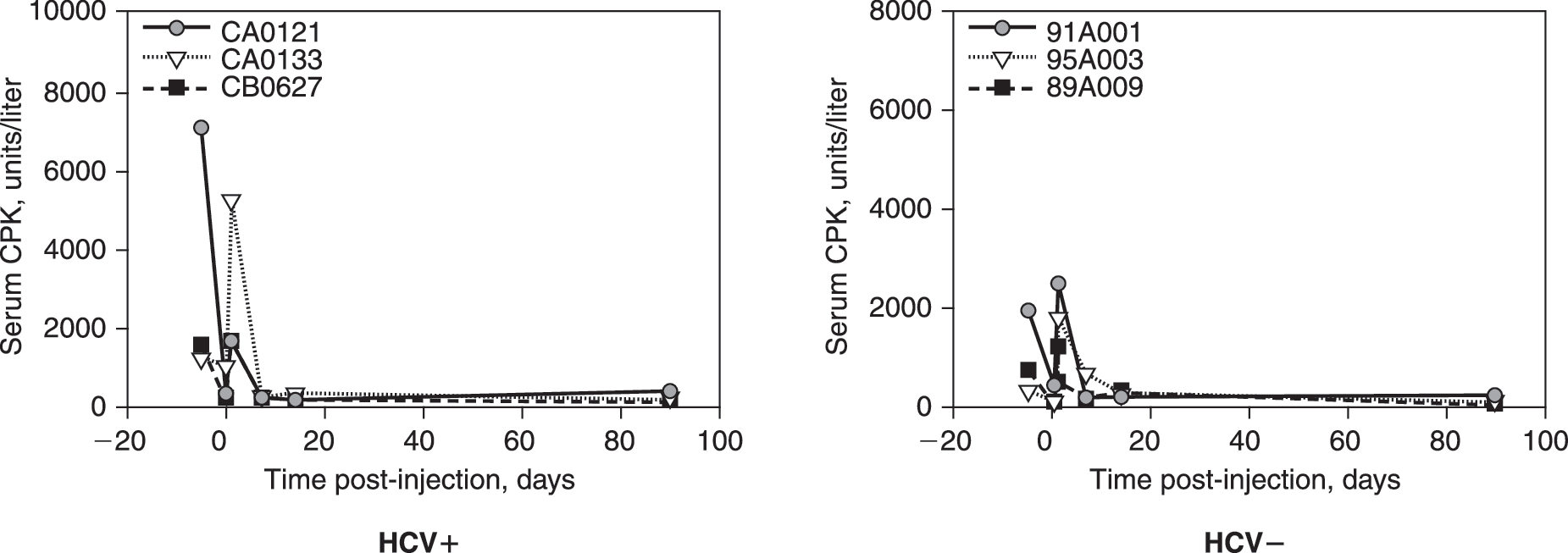
Serum CPK after portal vein injection of 1 × 1013 VG of rAAV2-chAAT into HCV+ versus HCV− chimps. Serum from each animal was analyzed for CPK at each indicated time point. The identification of each animal is indicated in the inset legend. Normal range for CPK enzyme in healthy chimpanzees is 0–792 U/liter.
Immune responses to rAAV2 capsid and AAT transgene product
All vector-treated chimpanzees underwent serum analysis for humoral immune responses, including assays for anti-AAV2 and anti-AAT antibodies by ELISA. Each chimpanzee demonstrated a greater than half-log rise in anti-AAV2 antibody levels after vector injection (Fig. 5A and B). There were no noticeable differences between the HCV+ and HCV− animals. None of the animals showed greater than background signal in the anti-AAT antibody assay at any time point (data not shown). Serum cytokine analysis on days –5, 0, 1, 7, 14, 28, and 90 for interleukin (IL)-6, tumor necrosis factor (TNF)-α, IL-1β, IL-2, IL-8, monocyte chemotactic protein (MCP)-1, and macrophage inflammatory protein (MIP)-1β did not reveal any differences between groups (data not shown).

Anti-AAV2 antibody titers and antigen-specific lymphocyte proliferation against AAV capsid after portal vein injection of 1 × 1013 VG of rAAV2-chAAT into HCV+ versus HCV− chimps. (
Antigen-specific lymphocyte proliferation assays were used as an indicator of cell-mediated immune responses, with a stimulation index of 2 (i.e., a 2-fold increase in tritiated thymidine incorporation) being used as a cutoff for a positive result. No such responses were observed either in response to AAV2 antigen (Fig. 5C and D) or in response to AAT antigen (data not shown).
Liver histopathology
Histopathologic analysis of liver biopsy samples showed a number of abnormalities, most of which were mild and not associated with the HCV status of the chimpanzees (Table 1). More notably, however, lipid accumulation (steatosis) was prominent in all HCV+ animals at all time points, including before vector injection (Fig. 6A). Semiquantitative scoring of steatosis indicated that this finding was seen exclusively in the HCV+ animals (Fig. 6B). There was a slight, nonsignificant downward trend over time. Time points spanning the time of vector injection, that is, from day –5 to day +7, did not reveal any relationship between the presence of steatosis and vector injection. The other incidental abnormalities, including focal hemosiderosis and focal areas of mild fibrosis, were seen sporadically in both HCV− and HCV− groups (Fig. 6C and D). There were no consistent differences from before to after vector treatment in either group. In addition, liver sections were examined for evidence of apoptosis, and none was noted in either group.
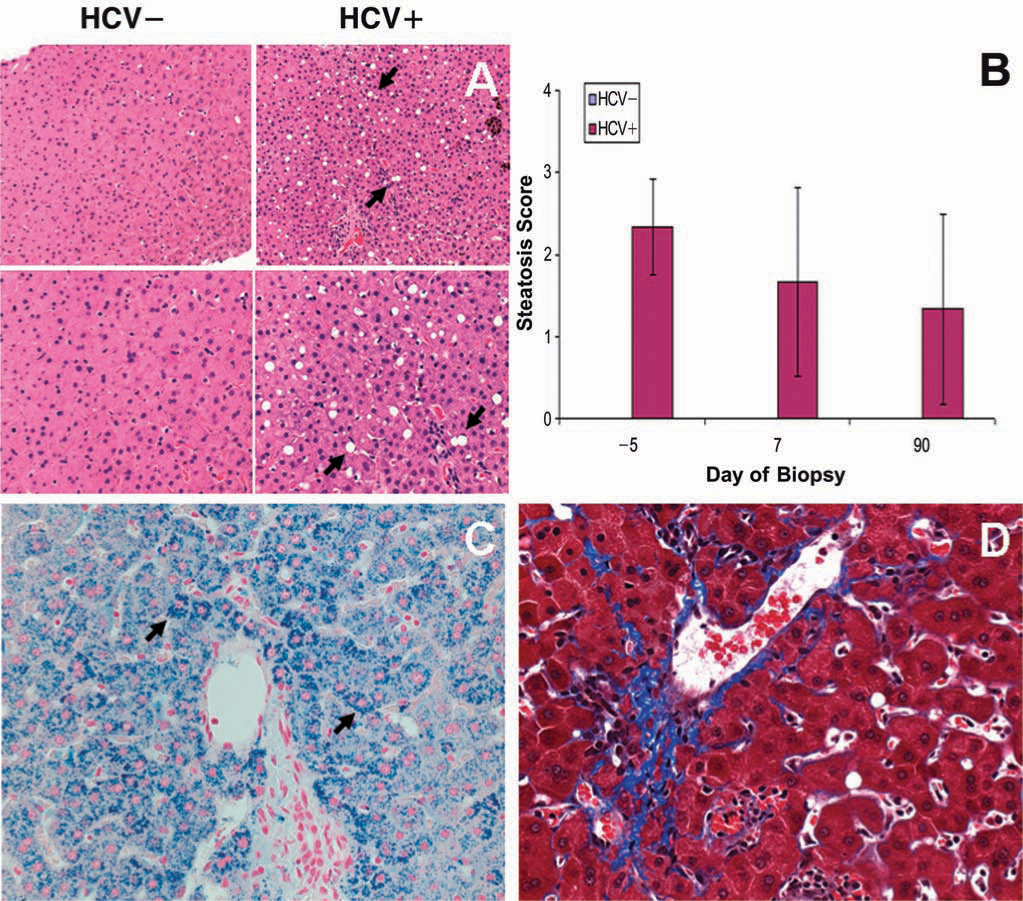
Steatosis, hemosiderosis, and fibrosis in liver biopsy samples from HCV+ versus HCV− chimpanzees. (
Discussion
The data presented here confirm that preexisting HCV infection can predispose chimpanzees to transient elevations of AST, ALT, and CPK or, as was the case for two HCV+ chimpanzees, persistent elevations in ALT. Serum enzyme elevations were seen after both liver biopsies and portal vein injections of vector. These elevations were not associated with clinical evidence of liver failure or muscle toxicity, but could have been interpreted as being possibly or probably related to the gene therapy vector, if they were encountered in the setting of a clinical trial. This possibility should be kept in mind when interpreting safety data generated in HCV-infected individuals.
The cellular origin of the enzyme elevations is not entirely clear. AST can originate from either hepatocytes or from muscle. The presence of elevation of the more liver-specific enzyme, ALT, and the muscle-specific enzyme, CPK, would seem to indicate that subclinical insults to both hepatocytes and muscle were occurring with liver manipulations. Although the two HCV+ chimpanzees with the highest HCV titers had a sustained increase in serum ALT levels as compared with baseline, it should be noted that even at baseline those levels were already outside the normal range (36–68 units/liter). Liver biopsy analysis demonstrated abnormal lipid accumulation in HCV+ animals, reinforcing the probability that at least some portion of the AST and ALT elevation was hepatic in origin. Even so, one cannot rule out some effect from the general inhalant anesthesia with isoflurane, which is used for both procedures.
Another key issue is the potential effect of adaptive immune responses to either the AAV2 capsid or to the trans-gene product. Our studies indicated that no detectable lymphocyte proliferation responses were present to either AAT or AAV2. However, a response cannot be ruled out as it has been shown that this assay is not as sensitive as an interferon (IFN)-γ enzyme-linked immunospot assay. As expected, we did not observe a response to the endogenous chimpanzee AAT either. However, anti-AAV2 antibodies were present, and could theoretically have contributed. The fact that elevations of the enzymes were seen even after the pretreatment liver biopsy argues against any form of adaptive response, however. Another potential factor would be greater innate responses from inflammatory cells that might be expected to reside in the livers of the HCV+ animals in higher numbers. Histopathologic findings did not suggest that large numbers of inflammatory cells were present, however.
The more probable explanation for these findings is that HCV-infected hepatocytes are more susceptible to injury when exposed to potentially injurious stimuli. Indeed, there seems to be an HCV titer-dependent effect on ALT serum levels, in which the two chimpanzees with the highest ALT levels exhibited a higher and sustained rise in ALT serum levels whereas the chimpanzee with the lower HCV titer had only a transient increase in ALT level after viral vector delivery. The presence of steatosis in the liver of the HCV+ animals, although not specific, is consistent with some form of hepatocyte distress as a primary problem. Furthermore, the susceptibility of the liver to demonstrate biochemical markers of injury after either biopsy or viral vector exposure is also consistent with this hypothesis.
The implication of these findings for future clinical trials of rAAV2-mediated gene delivery to the liver is unclear. In the hemophilia B trial discussed above, the serum enzyme abnormalities did not correlate with clinically significant organ dysfunction (Manno et al., 2006). However, it is particularly worrisome if one were to use rAAV2 to treat disorders such as AAT deficiency; many of these patients have some underlying liver disease. Importantly, it is not known whether other rAAV serotypes that are more efficient for liver transduction, such as AAV8, will also elicit these enzyme elevations. The HCV-infected chimpanzee model could be appropriate for studies with these other serotypes to address this question in future studies. The possibility that the vector infusion procedure itself causes an insult to the liver that, in combination with the underlying HCV disease, and hence may be responsible for the rise in the liver function test results cannot be ruled out. Despite this, the potential for an adverse effect in populations with underlying HCV liver disease remains real. In addition to testing alternative serotypes, this model can be used in future to determine and tease out how much of the adverse effect is related to the vector or infusion procedure.
Footnotes
Acknowledgments
The authors thank the personnel of the Powell Gene Therapy Center's NGVL Toxicology Core Laboratory and Vector Core Laboratory for vital technical assistance. This work was supported by grants from the NIDDK (DK58327), the NCRR (RR16586), and the NHLBI (HL69877 and HL59412).
Author Disclosure Statement
No competing financial interests or potential conflict of interests exist for any of the authors listed on this publication.