
Abstract
Pancreatic ductal adenocarcinoma (PDAC) is still the fourth leading cause of cancer-related deaths in Western countries, with increasing incidence. Neither effective prognostic markers nor therapies exist for this cancer. MicroRNAs are potent inhibitors of protein translation, and aberrantly expressed in many cancers. Because let-7 microRNA targets the K-ras oncogene, we aimed to characterize let-7 expression and function in PDAC in vitro and in vivo. Let-7 expression was quantified by real-time RT-PCR from resected tumors and matching adjacent tissue, and in endoscopic ultrasound-guided fine needle aspiration material from patients with PDAC. Let-7 is detected by reverse transcription in situ PCR in a PDAC tissue microarray. PDAC-derived cells were transfected with plasmid-based synthetic microRNAs or by lentiviral transduction, in vitro and in vivo. Let-7 microRNA expression is strongly reduced in PDAC samples, as compared with adjacent tissue. Let-7 is present in normal acinar pancreatic cells, and lost in poorly differentiated cancer samples. In addition, let-7 expression was repressed in patients with PDAC not eligible for surgery. Restoring let-7 levels in cancer-derived cell lines strongly inhibits cell proliferation, K-ras expression, and mitogen-activated protein kinase activation, but fails to impede tumor growth progression after intratumoral gene transfer or after implantation of Capan-1 cells stably overexpressing let-7 microRNA. We describe here for the first time the extensive loss of expression of let-7 in PDAC. In addition, this study provides the initial steps for a microRNA replacement therapy for this cancer.
Introduction
The proto-oncogene Kirsten-ras (K-ras) is mutated in more than 80% of PDACs and thus is considered a potential molecular marker and target for PDAC treatment (Costentin et al., 2002). Interestingly, let-7 microRNA inhibits K-Ras protein expression in human cancer-derived cells (Morris and McManus, 2005). In Caenorhabditis elegans, let-7 controls cell cycle exit timing and differentiation (Johnson et al., 2005). In humans, let-7 coding regions map to regions altered or deleted in human tumors (Calin et al., 2004). A deregulation of let-7 is associated with the genesis and maintenance of the lymphoma phenotype in Burkitt lymphoma cells (Sampson et al., 2007). In addition, let-7 represses the high-mobility group-2 (HMGA2) oncogene (Mayr et al., 2007), to inhibit lung cancer cell-derived proliferation (Takamizawa et al., 2004; Lee and Dutta, 2007). Johnson and colleagues demonstrated that let-7 alters several signaling pathways in cancer-derived cells, such as mitotic signaling, cell cycling, angiogenesis, and cell adhesion and migration (Johnson et al., 2007). Considering that the current cancer treatment regimens are directed toward single targets (including let-7-targeted mRNAs) the therapeutic benefit of let-7, and of microRNAs in general, may be greater. Enthusiasm for the therapeutic development of let-7 is heightened by the fact that let-7 is a naturally occurring molecule with biological implications in cancer, which supports the concept of microRNA replacement therapy. Therefore, this study was aimed at characterizing let-7 microRNA expression in PDAC and at determining whether restoring let-7 expression in PDAC-derived cells inhibits cell proliferation and tumor progression.
Materials and Methods
Cell lines
Capan-1, Capan-2, BxPC-3, MIA PaCa-2, and PANC-1 cell lines derived from human PDACs were obtained from the American Type Culture Collection (Manassas, VA) and cultured as previously described (Torrisani et al., 2001). Human HPNE (human pancreatic nestin-positive) cells derived from pancreatic ducts were obtained from M. Ouellette (University of Nebraska Medical Center, Omaha, NE) and were grown as described (Lee et al., 2003). All media were supplemented with amphotericin B (Fungizone; Invitrogen, Cergy Pontoise, France), antibiotics (Invitrogen), and anti-Mycoplasma Plasmocin reagent (InvivoGen, Toulouse, France).
Clinical samples
Pancreatic tissue samples were collected from pancreatic surgery patients, in accordance with the policies and practices of the facility's ethics committee at the Centres Hospitaliers Universitaires (CHU) of Toulouse and Bordeaux, and Cancéropôle Grand Sud-Ouest (Table 1). All patients gave their informed written consent. Histopathology faculty selected cancerous pancreatic tissue with matched adjacent tissue.
Abbreviations: PDAC, pancreatic ductal adenocarcinoma; UICC, international Union against Cancer (Geneva, Switzerland).
Samples were matched with adjacent normal tissue. Tissue sections 13 to 20 were used for in situ detection of let-7.
UICC classification.
Endoscopic ultrasound (EUS)-guided fine needle aspiration (FNA) was performed in patients ineligible for surgery because of advanced disease or who were in poor health, using a linear ultrasound endoscope (Olympus America [Center Valley, PA] and Pentax Medical [Montvale, NJ]) and an EUSN1 22G needle (Cook Medical, Bloomington, IN). The protocol was approved by the Ethics Committee of Midi-Pyrénées CPPRB-1. All patients gave their informed written consent before EUS. FNA tissue material was transferred into Dubosq-Brazil medium or formalin. Residual cellular material was collected in RNAlater (Qiagen, Courtaboeuf, France) and stored at −20°C. Twelve patients diagnosed with PDAC after histopathological analysis of microbiopsies were included at CHU Toulouse (Table 2: female, n = 5; male, n = 7; mean age, 65 ± 2.6 years).
Abbreviations: EUS, endoscopic ultrasound; FNA, fine needle aspiration; id, inner diameter; TNM, cancer staging system (T, tumor size; N, lymph node spread; M, metastasis).
Grade I, well differentiated; grade II, moderately differentiated; grade III, poorly differentiated.
Let-7a coding sequence was cloned into the psiSTRIKE U6 hairpin cloning system (Promega, Charbonnières-les-Bains, France), to obtain psiSTRIKE(Let-7). As control, a small nuclear RNA (siRNA) targeting firefly luciferase (Luc) was cloned into the psiSTRIKE U6 hairpin cloning system (Promega) to generate psiSTRIKE(Luc). Plasmids were verified by sequencing (MilleGen, Labége, France). pmaxGFP vector was purchased from Amaxa (Walkersville, MD). pT7A5(RFP) vector was previously described (Cordelier et al., 2003).
Oligonucleotides 5′-tgctgtgaggtagtaggttgtatagttgttttggccactgactgacaactataccctactacctca-3′ and 5′-cctgtgaggtagtagggtatagttgtcagtcagtggccaaaacaactatacaacctactacctcac-3′ were annealed and cloned into pcDNA-6.2-GW/emGFP-miR vector (Invitrogen) to obtain pcDNA6.2-GW/emGFP-miR-Let-7 vector. pcDNA6.2-GW/emGFP-miR-Neg vector (Invitrogen) was used as control. GFP-miR-Let-7 and GFP-miR-Neg cassettes were cloned into pDONR221 (Invitrogen), using the BP Clonase reaction (Invitrogen), and subsequently transferred into pLenti4/TO/V5-DEST lentiviral vector (Invitrogen), using the LR Clonase reaction (Invitrogen) to create pLenti4/TO/GFP-miR-Let-7 and pLenti4/TO/GFP-miR-Neg vectors, respectively.
Cell transfection
Cells were transfected with pre-microRNAs (Ambion/Applied Biosystems, Courtaboeuf, France) at 10 nM final concentration, or with 1 μg of microRNA-encoding plasmid [psiSTRIKE(Let-7) and psiSTRIKE(Luc)], using siPort NeoFx reagent (Ambion/Applied Biosystems) and Nucleofector II (Amaxa, Cologne, Germany), respectively. Cy3-control premicroRNA was purchased from Ambion/Applied Biosystems.
Cell proliferation and cell viability assays
Cell proliferation assays were performed in 35-mm dishes for 48 hr. Cells were counted with a Coulter counter model ZM (Beckman Coulter, Roissy, France) as described by Buscail and colleagues (1992).
LV-miR-Neg and LV-miR-Let-7 Capan-1 cells (see below) were seeded at 6 × 103 cells per well of a 96-well dish. After 96 hr, cell viability was measured using a CellTiter 96 AQueous One Solution cell proliferation assay (MTS) (Promega).
RNA extraction
Total RNA was extracted from cell lines and tumors with TRIzol reagent (Invitrogen). Quality and concentration of the RNA were determined with an Experion electrophoresis system (Bio-Rad, Marnes La Coquette, France) and an ND-100 nanospectrophotometer (Thermo Scientific, Courtaboeuf, France), respectively. RNA samples displayed a 28S/18S ratio > 1.8. Total RNA from FNA was purified with an RNeasy micro kit (Qiagen).
MicroRNA detection in FNA
Five nanograms of total RNA isolated from FNA samples was amplified as complementary DNA (cDNA), using a Full Spectrum global amplification kit (System Biosciences, Mountain View, CA) as per the manufacturer-recommended conditions. As control, we used 5 ng of total RNA from normal human pancreas and tissue samples adjacent to PDAC. cDNAs were diluted 1:50 (pre-let-7a-1) or 1:50,000 (18S rRNA) before real-time PCR was performed in duplicate with an ABI PRISM 7900 sequence detection system (Applied Biosystems), SYBR green PCR master mix (Applied Biosystems), and specific primers for let-7a precursor as described (Takamizawa et al., 2004). Expression values were calculated according to the comparative C T method as 2−ΔC T [ΔC T = C T(let-7a) − C T(18S)] and arbitrarily multiplied by 102.
MicroRNA detection in cell lines and tumors
Quantification of mature microRNA was conducted on 100–500 ng of total RNA extracted from cell lines and tumors, using the miScript PCR system (Qiagen) as per the manufacturer's instructions. U6, U18, and 5S were used as controls. cDNA samples were diluted (1:10) before duplicate real-time PCR assays for microRNA (Qiagen) were performed with an ABI PRISM 7900 sequence detection system (Applied Biosystems), with SYBR green PCR master mix (Qiagen). Expression values were calculated according to the comparative C T method as 2−ΔC T [ΔC T = C T(let-7a) − C T(geometric mean U6, 5S, U18)] and arbitrarily multiplied by 102.
Reverse transcription in situ PCR
The tissue microarray (TMA; Euromedex, Souffelweyersheim, France) comprises 70 PDAC sections (female, n = 25; male, n = 45; mean age, 56.6 ± 11 years) with 49 cases of stage I cancer, 13 cases of stage II, 5 cases of stage III, and 3 cases of stage IV according to the International Union against Cancer (UICC, Geneva, Switzerland) classification. All were proven PDAC (grade I/well differentiated, n = 6; grade II/moderately differentiated, n = 38; and grade III/poorly differentiated, n = 20; not determined in 6 cases). The TMA also contains nine sections of normal pancreas (female, n = 5; male, n = 4; mean age, 52.8 ± 12 years). The RT in situ PCR was conducted as previously described, with minor modifications (Cordelier et al., 2003). Briefly, paraffin-embedded TMAs were deparaffinized in xylene and rehydrated in descending grades of ethanol. Tissues were digested with pepsin (Dako, Trappes, France), and incubated in 0.2 N HCl for 30 min at room temperature. Deacetylation was conducted in 100 mM triethanolamine (pH 8.0) for 10 min with gentle agitation, before treatment with 1 U of DNase I (Invitrogen). Slides were postfixed in 4% paraformaldehyde, rinsed with phosphate-buffered saline (PBS), and dehydrated in ascending grades of ethanol. RNA was converted to cDNA on slides, using a single-step RT-PCR kit (Invitrogen), biotin-conjugated dUTP (Applied Biosystems), Self-Seal reagent (Bio-Rad), and 1.2 μM concentrations of primers targeting pre-let-7a-1 RNA (Takamizawa et al., 2004) in a TC-512 gradient thermal cycler with an in situ block (Thermo Scientific). In control slides, primers, RT/Platinum Taq polymerase mix (Invitrogen), and/or conjugated dUTPs were omitted. After incubation at 55°C for 30 min, slides were heated at 94°C for 2 min before performing 20 cycles consisting of 94°C for 30 sec, 55°C for 1 min, and 72°C for 2 min. A final extension was performed at 72°C for 10 min. Slides were incubated in 100% ethanol for 1 min and washed in 2 × standard sodium chloride–sodium citrate (SSC: 3 M NaCl, 0.3 M trisodium citrate), 2× SSC − 0.5% sodium dodecyl sulfate (SDS), 1× SSC − 0.5% SDS, and 0.2× SSC − 0.5% SDS at room temperature for 5 min each. Sections were treated with StreptABComplex/HRP (horseradish peroxidase) reagent and AEC + high-sensitivity substrate (Dako), in accordance with the manufacturer's recommendations. After staining with Mayer's hematoxylin, slides were observed with an optical microscope, coupled to an image analyzer (VisioLab2000; Biocom, Les Ulis, France).
Western blotting analysis
Total proteins were extracted from Capan-1 cells and separated by SDS–10% polyacrylamide gel electrophoresis. Proteins were transferred onto a nitrocellulose membrane (Biotrace NT; Pall, Saint-Germain-en-Laye, France) as previously described (Sirach et al., 2007). After saturation at room temperature for 1 hr, membranes were incubated with antibodies directed against either K-Ras (diluted 1:250; Santa Cruz Biotechnology, Le Perray en Yvelines, France) or phospho-specific p42/p44 mitogen-activated protein kinase (MAPK) (Cell Signaling Technology, Saint Quentin Yvelines, France). Secondary anti-mouse or anti-rabbit HRP-conjugated antibodies (diluted 1:10,000; Perbio Science, Brebières, France) were added. Membranes were incubated for 1 hr at room temperature before chemoluminescence detection (Immobilon; Millipore, Vandoeuvre, France). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) protein was used as loading control (diluted 1:1000; Santa Cruz Biotechnology).
Generation of pancreatic cancer Capan-1 stable cell line overexpressing let-7 microRNA
Lentiviral particles were produced with pLenti4/TO/GFP-miR-Let-7 and pLenti4/TO/GFP-miR-Neg vectors and ViraPower lentiviral packaging mix (Invitrogen). Capan-1 cells were incubated with the lentiviral particles (multiplicity of infection, 1) for 24 hr and subsequently selected with Zeocin (Invivogen, Toulouse, France) at 25 μg/ml for 2 weeks.
Efficiency of viral infection and selection was estimated by measuring the percentage of GFP-expressing cells by flow cytometric analysis (see below). Overexpression of mature miR-Let-7 was quantified by RT real-time PCR (see above).
Flow cytometric analysis
Control and LV-miR-Let-7-transduced cells were harvested after trypsinization and collected by centrifugation at 1500 rpm for 5 min. Cells were washed once with PBS. The number of EGFP-positive cells was determined by flow cytometric analysis with a FACSCalibur system (Becton Dickinson France, Le Pont-De-Claix, France).
In vivo model of pancreatic cancer
An in vivo model of xenografted PDAC-derived cells into athymic mice has been extensively described (Carrere et al., 2005). Briefly, 5 × 106 exponentially growing Capan-1 cells in 100 μl of PBS were subcutaneously inoculated into 8-week-old athymic female nude mice (Swiss nude/nude; Iffa-Credo, l'Abresle, France) bred and maintained under pathogen-free conditions. Fourteen days after xenografting, intratumoral in vivo gene transfer was performed with linear polymers of ethylenimine (polyethylenimine [PEI], 22 kDa, ExGen 500; Euromedex). PEI:DNA complexes were prepared at a PEI nitrogen-to-DNA phosphate (N:P) ratio of 9 in 5% (w/v) glucose. Ten micrograms of psiSTRIKE(Let-7) plasmid was injected into the tumors with a sterile 29-gauge lymphography catheter set with a sterile, 29-gauge insulin needle. Control animals received 10 μg of psiSTRIKE(Luc) plasmid. Alternatively, in vivo transfection of either random or let-7 precursor microRNA (1 μM) was performed with siPort NeoFx reagent. Subcutaneous tumors were externally measured after gene transfer. Four to six mice were used per group.
LV-miR-Neg and LV-miR-Let-7 Capan-1 cells were subcutaneously injected into 8-week-old athymic mice (5 × 106 in 100 μl of PBS solution per animal). Three to five mice were used per group. Tumor volumes were externally measured from 7 days post-injection until day 27.
Statistical analysis
Results are expressed as means ± standard error (SE). An unpaired Student t test calculated with Graphpad Instat software (Graphpad Software, La Jolla, CA) was used to compare data (* p < 0.05, ** p < 0.01, *** p < 0.001). p < 0.05 was considered statistically significant.
Results
Reduced expression of let-7 microRNA in PDAC samples
Let-7 expression was analyzed in PDAC samples after surgery (Table 1). As control, we used corresponding tissues adjacent to the tumors. Total RNA was extracted and subjected to let-7 microRNA detection as described in Materials and Methods. As shown in Fig. 1A, let-7 expression is downregulated in PDAC samples as compared with their adjacent counterpart in 11 of 12 patients (91%). Altogether, let-7 expression was reduced by 59.9 ± 8.5% (p < 0.001) in PDAC samples (Fig. 1B).
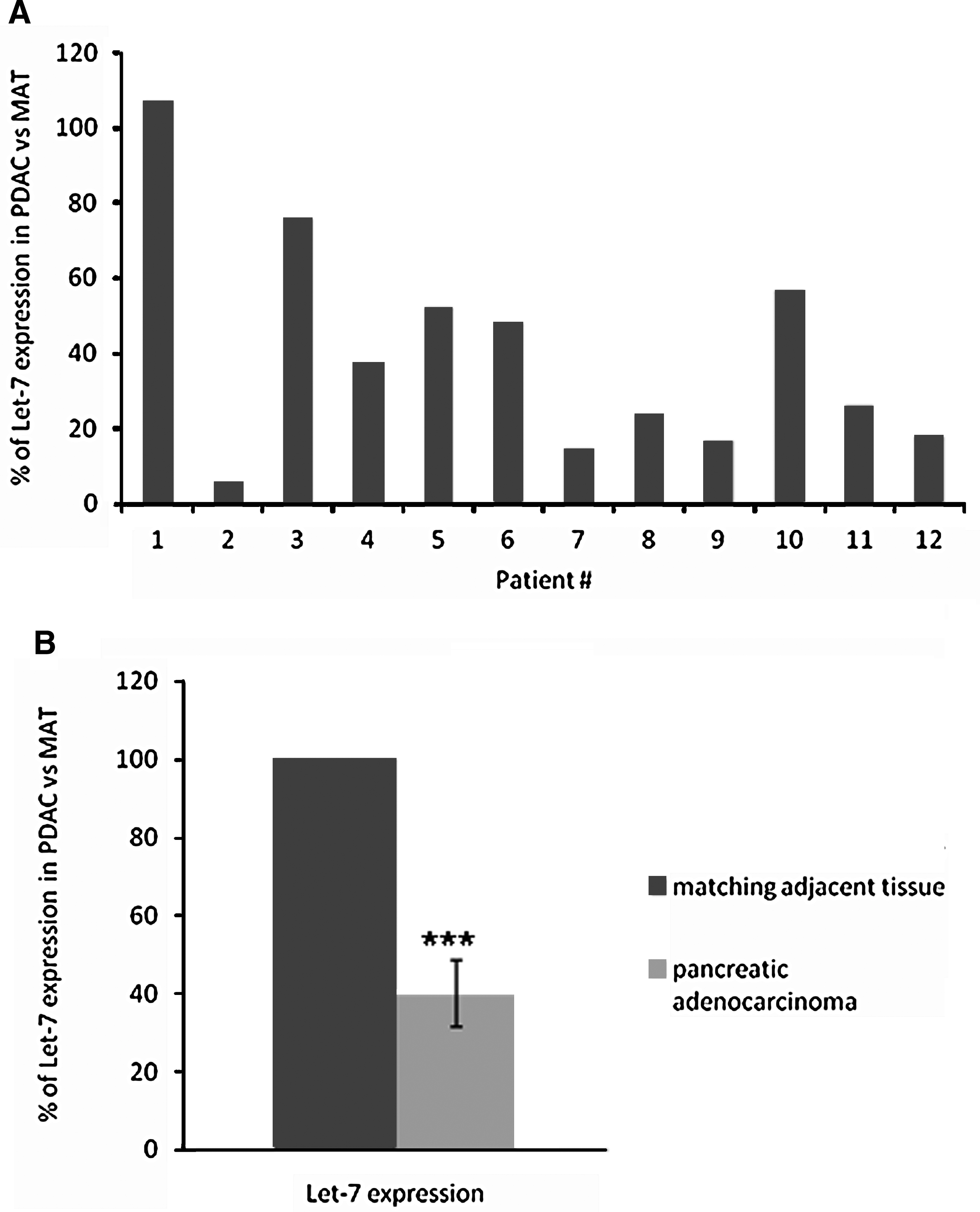
let-7 microRNA expression in resected pancreatic cancer. (
To identify the cell population expressing let-7 in normal and diseased pancreas, we performed reverse transcription in situ PCR (Fig. 2A). Whereas no specific signal is present in control slides, let-7 was strongly expressed in pancreatic acini adjacent to PDAC but not in stromal or cancerous cells. Extending let-7 detection to the PDAC tissue macroarray (TMA), we found that let-7 was present in 88% of normal pancreatic samples (eight of nine). In contrast, let-7 expression is inversely correlated with histological grade, that is, expression was weak in poorly differentiated samples (Fig. 2B). Taken together, these results demonstrated that let-7 expression is detected in normal pancreas, and in pancreatic acini adjacent to the tumor, whereas its expression is decreased in poorly differentiated PDAC samples.

In situ detection of let-7 microRNA in pancreatic cancer. (
Endoscopic ultrasound (EUS) has become an essential modality in diagnosis and management of pancreatic malignancies (Buscail et al., 2006). We identified molecular markers for PDAC by gene expression analysis of EUS-guided fine needle aspiration (FNA) material (Laurell et al., 2006). Because of the paucity of the collected material, a constant amount of total RNA (5 ng) was amplified to maintain representation of gene expression in the original sample (Steg et al., 2006). However, because of their small size, mature microRNAs are excluded by this technique. Therefore, we quantified let-7 precursors as previously described (Takamizawa et al., 2004). 18S rRNA was selected as internal control because its expression is conserved in all the samples analyzed (Fig. 3A). We observed that let-7 precursors are expressed in tissue adjacent to the tumor and normal pancreas (Fig. 3B). Again, let-7 precursors are strongly reduced in PDAC samples. let-7 levels were extremely low (≤1) in 10 of 12 (83.3%), moderate (<100) in 1 of 12 (8.3%), and high (>100) in 1 of 12 (8.3%) EUS-FNA samples from patients diagnosed with PDAC. In conclusion, we demonstrated that let-7 levels are strongly reduced in both resected PDAC tumors, and FNA material.

let-7 detection in endoscopic ultrasound-guided fine needle aspiration (EUS-FNA) material from patients with pancreatic cancer. Total RNA was purified from normal pancreas, normal adjacent tissue, and EUS-FNA material before amplification as described in Materials and Methods. (
Overexpression of let-7 in PDAC-derived cell line strongly inhibits cell proliferation, K-ras expression, and MAPK activation
The data presented here describing the reduced expression of let-7 in PDAC samples, together with its negative role in K-Ras protein expression, prompted us to explore the effect of let-7 overexpression on PDAC-derived cell proliferation. Let-7 expression was quantified in cell lines derived from primary tumors or metastasis. Human primary HPNE cells, derived from pancreatic ducts, were used as control (Lee et al., 2003). Whereas HPNE cells express high levels of let-7, PDAC-derived cell lines displayed low levels of let-7, similarly to PDAC samples (Fig. 4A). In summary, let-7 expression is reduced by 82.5 ± 4.5% (p < 0.001) in PDAC-derived cells as compared with normal pancreatic cells (Fig. 4B).

let-7 expression in pancreatic cancer-derived cells. (
Next, Capan-1 cells were used as a model of K-ras-mutated (verified by restriction fragment length polymorphism [RFLP] and sequencing; Costentin et al., 2002), aggressive PDAC-derived cells (Carrere et al., 2005). We first optimized the transfection conditions for subsequent expression of both precursor and mature let-7 microRNAs in these cells. Using nucleofection, Capan-1 cells were transfected to obtain detectable levels (39 ± 8%), with minimal toxicity (Fig. 5A, left). As expected, the use of synthetic microRNA precursors resulted in high levels of transfected cells (93 ± 5%), (Fig. 5A, right). We also transfected Capan-1 cells with plasmids encoding mature let-7 microRNA [psiSTRIKE(Let-7)], or synthetic let-7 microRNA precursors. As control, cells were transfected with an anti-luciferase siRNA-encoding plasmid or with a negative control microRNA precursor. By qRT-PCR, we measured 5.3 ± 1.6-fold and 8.2 ± 3.1-fold increases in let-7 expression after transfection with psiSTRIKE(Let-7) and let-7 precursors, respectively (Fig. 5B). Interestingly, let-7 levels in transfected Capan-1 cells were similar to those in normal pancreatic cells.
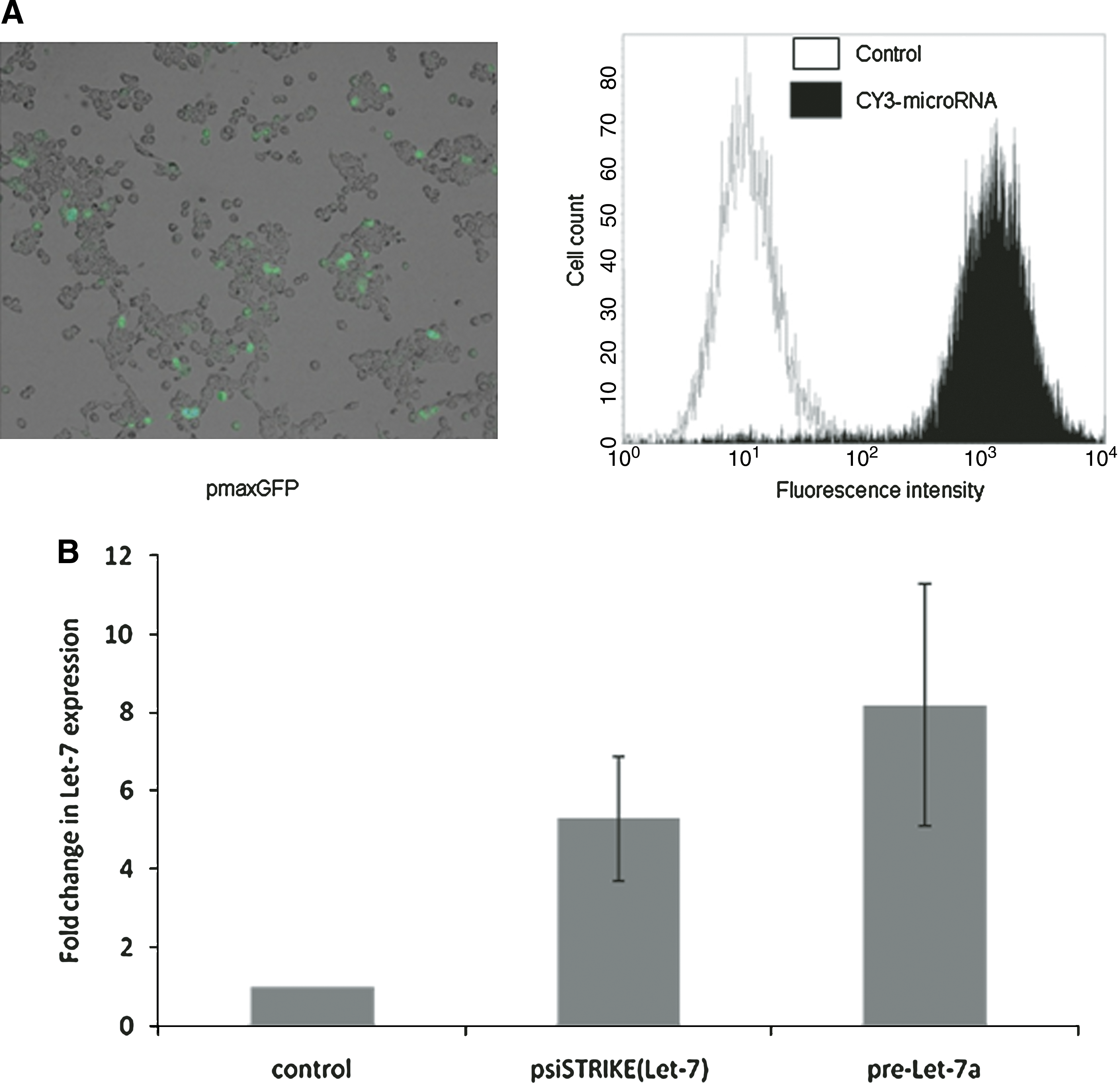
let-7 transfection in pancreatic cancer-derived cells. (
We examined the transfected cells for alteration in cell proliferation. As shown in Fig. 6A, let-7 expression in Capan-1 cells resulted in reduced in vitro cell confluence. We quantified the extent of the let-7 antiproliferative effect. As shown in Fig. 6B, both plasmid-encoding and synthetic let-7 microRNAs strongly inhibit Capan-1 cell proliferation, as compared with control-transfected cells (−52.5 ± 12.8% and −48 ± 3%, respectively; p < 0.001). We further studied let-7 function in PDAC-derived cells, by analyzing K-Ras protein expression, a canonical let-7 target (Johnson et al., 2005) (Fig. 6C). We found that K-Ras expression and MAPK phosphorylation, a downstream effector of K-Ras, are decreased in Capan-1 cells after let-7 transfection (−59 ± 17% and −45 ± 18%, respectively; p < 0.05). Thus, these data demonstrate that restoring let-7 expression in PDAC-derived cell lines strongly inhibits cell proliferation, K-Ras expression, and MAPK activation.
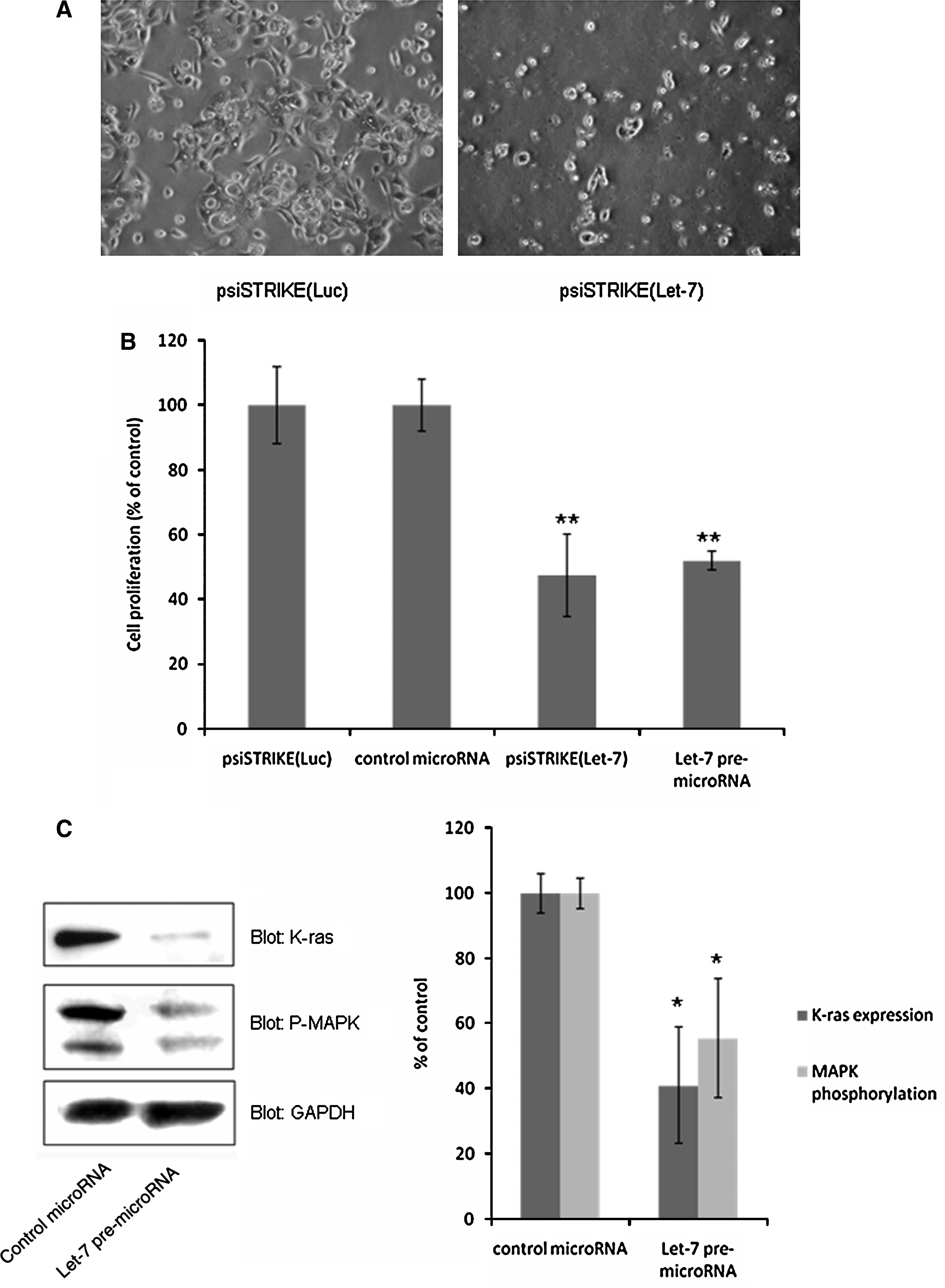
let-7 overexpression in pancreatic cancer-derived Capan-1 cells. (
Restoring let-7 expression in PDAC tumors
We tested whether let-7 transfection could inhibit in vivo PDAC tumor progression. Subcutaneous Capan-1 xenografts were established in athymic mice as an ectopic model of PDAC (Carrere et al., 2005; Cordelier et al., 2007). Fourteen days after tumor engraftment, PEI–DNA complexes or synthetic microRNA precursors were in vivo transferred into subcutaneous Capan-1 pancreatic tumors. Tumor growth was measured every 2 to 5 days, up to 14 days after gene transfer. Tumor analyses demonstrated detectable in vivo plasmid-driven marker gene expression and Cy3-labeled synthetic control microRNA detection up to 7 days after transfer (Fig. 7A and B). Again, transfection efficiency was higher with synthetic microRNAs. However, neither psiSTRIKE(Let-7) (Fig. 7C) nor let-7 precursor microRNA transfection (Fig. 7D) delayed tumor progression. In addition, no statistical difference in tumor weight or in tumor cell proliferation (asserted by Ki-67 labeling; data not shown) was found in response to let-7 overexpression.
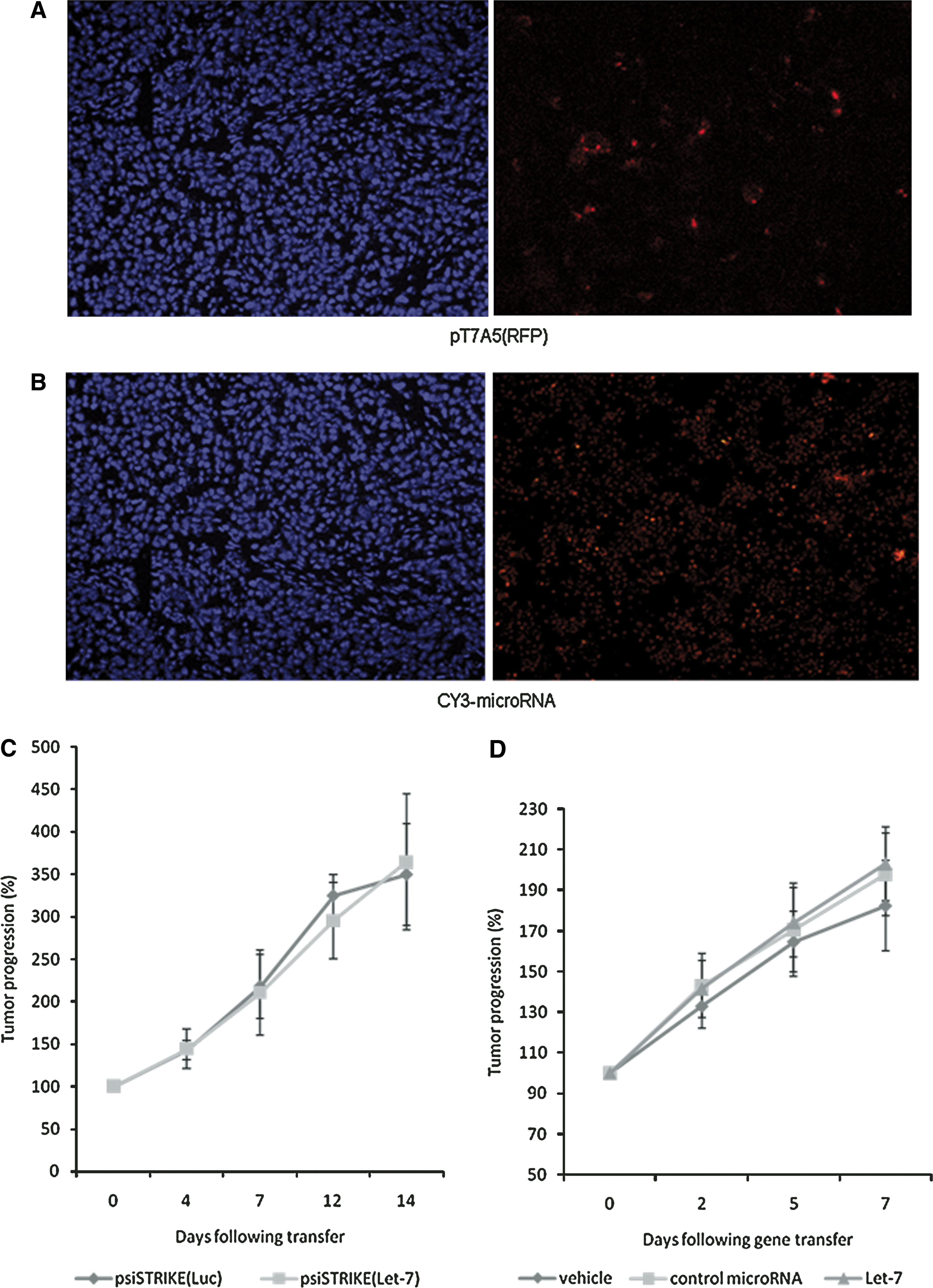
In vivo let-7 microRNA transfer into exponentially growing pancreatic tumors. Capan-1 tumors were established in athymic mice as described in Materials and Methods. Fourteen to 17 days later, pT7A5(RFP) plasmid encoding (
To ensure that the absence of effect of let-7 overexpression was not due to low in vivo transfection efficiency, we generated a stable Capan-1 cell line overexpressing let-7 microRNA (LV-miR-Let-7), using lentiviral transduction. As control, we also established a stable Capan-1 cell line overexpressing negative control microRNA (LV-miR-Neg). We determined by FACS analysis that more than 90% of the cell population was transduced (Fig. 8A). We also verified that the LV-miR-Let-7 cell line overexpressed let-7 microRNA (Fig. 8B). As expected, we observed that overexpression of let-7 microRNA reduces the viability of the stable Capan-1 cell line (Fig. 8C).
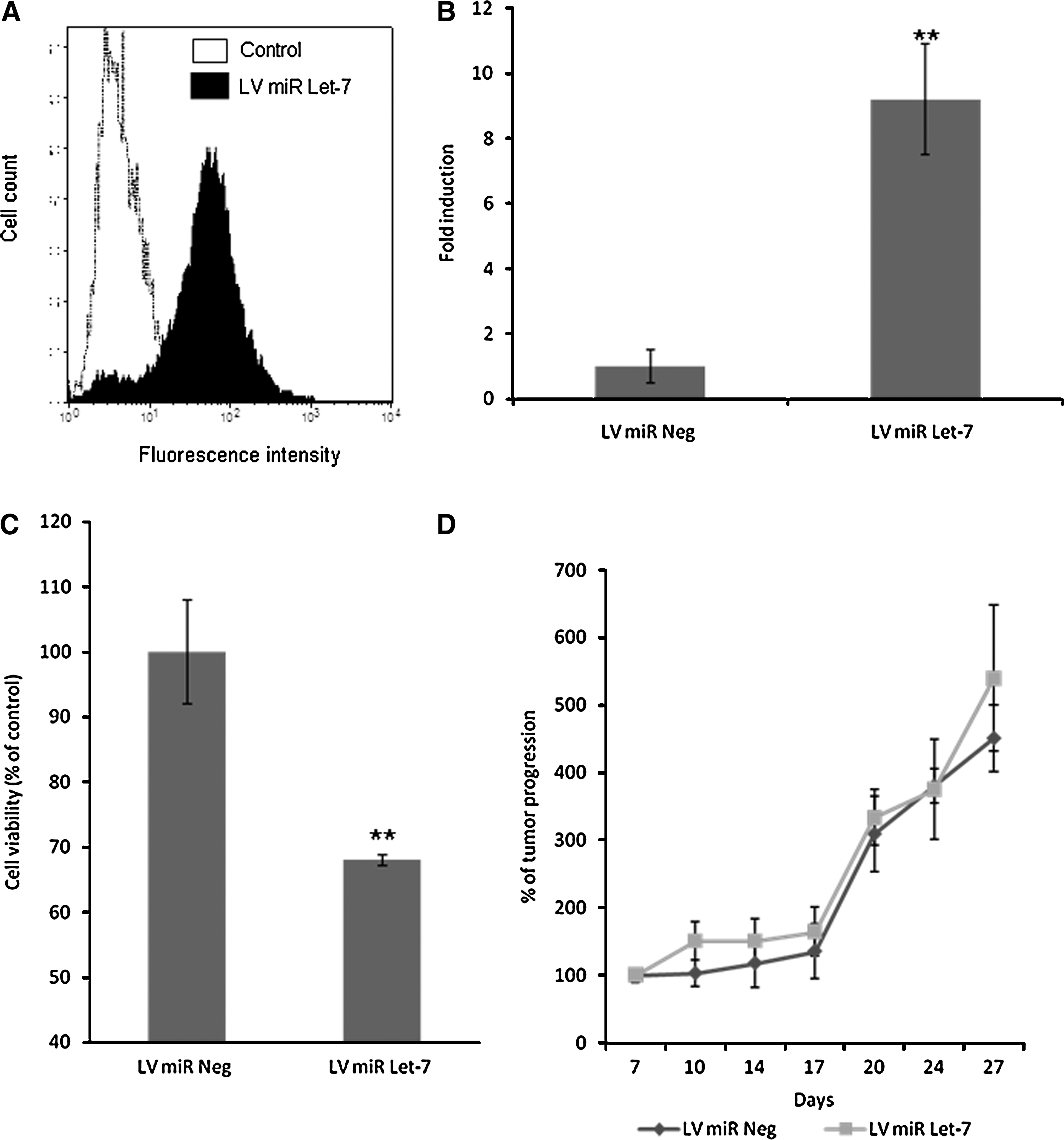
In vitro and in vivo effect of stable Let-7 expression in Capan-1 cells. (

Further, to determine the effect of stable overexpression of let-7 microRNA on tumor progression, LV-miR-Let-7 and LV-miR-Neg Capan-1 cells were injected subcutaneously into athymic mice. Seven days after inoculation, tumor growth was measured every 3–4 days until day 27 (Fig. 8D). These measurements revealed that let-7 overexpression did not delay in vivo tumor progression, although overexpression of let-7 microRNA is still present in tumors 27 days after inoculation (Fig. 8E).
Altogether, these results demonstrate that whereas let-7 microRNA potentially inhibits pancreatic cancer cell proliferation in vitro, let-7 microRNA gene transfer or stable overexpression fails to impede in vivo tumor growth.
Discussion
MicroRNA alterations have been linked to the initiation and progression of cancer (Bandres et al., 2007). MicroRNA expression was further associated with diagnosis, staging, prognosis, and response to treatment (Cummins and Velculescu, 2006). Pioneering studies described let-7 as a negative regulator of the oncogenic family of Ras guanosine triphosphatases in both Caenorhabditis elegans and human tumor cell lines (Johnson et al., 2005). Later, deregulation of let-7 expression was reported in several cancers, suggesting that let-7 may act as a tumor suppressor (Torrisani et al., 2007). The broad changes in microRNA profiles at late stages of PDAC have been reported by several independent groups (Bloomston et al., 2007; Lee et al., 2007; Szafranska et al., 2007). Hahn and colleagues found that the ratio of miR-196 and miR-217 expression provides a reliable index to identify diseased pancreatic tissues (Szafranska et al., 2007, 2008). Moreover, Bloomston and colleagues demonstrated that few microRNAs can differentiate PDAC from chronic pancreatitis or predict poor survival (Bloomston et al., 2007).
We focused on let-7 because this microRNA targets K-ras (Johnson et al., 2005), a key oncogene in PDAC carcinogenesis (Costentin et al., 2002). Using quantitative RT-PCR (qRT-PCR), we found that let-7 expression is strongly inhibited in PDAC samples as compared with adjacent tissue to the tumor. These findings are comforted by two independent studies, which reported a significant downregulation of let-7 microRNA in PDAC as determined by microarray analyses (Bloomston et al., 2007; Szafranska et al., 2007). However, in these studies, let-7 microRNA was not one of the most differentially expressed microRNAs between PDAC, chronic pancreatitis, and normal cancer. Therefore, its expression was not further studied by qRT-PCR.
However, we provide here for the first time a comprehensive examination of the molecular anatomy of cells in tissue sections expressing let-7. Because conventional in situ hybridization was not sensitive enough to detect let-7 in our samples (data not shown), we devised a new reverse transcription in situ PCR-based approach for let-7 detection in tissue sections. We confirmed that let-7 is less abundant in cancer and stromal cells, when pancreatic acini adjacent to the tumor expressed high levels of this microRNA. We measured let-7 expression in 70 sections of proven, resected PDAC from stage I to IV. We found that let-7 expression negatively correlates with pathology, differentiation, and stage disease. We also quantified let-7 expression in EUS-FNA material. We confirmed let-7 loss of expression in PDAC, and demonstrated the feasibility of using microRNAs as potential molecular markers in routine clinical practice for the diagnosis of patients ineligible for surgery. During the redaction of this manuscript, Szafranska and colleagues demonstrated that FNA material permitted microRNA detection (Szafranska et al., 2008). Because of its invasive nature, FNA is not likely to be used for PDAC detection. However, screening high-risk individuals, establishing a prognosis, and predicting the response to treatment in the numerous cases in which the tumor is not operable may be achieved by such a procedure. Because we found that let-7 expression is lost in poorly differentiated late-stage disease, detectable let-7 microRNA expression in EUS-FNA material may be of potential interest to select patients amenable for further management.
Reasons for the loss of let-7 expression in PDAC samples remain unknown. MicroRNAs are frequently located at fragile sites and genomic regions involved in cancers (Calin et al., 2004). The let-7a gene is hosted by chromosome 9q12, which is not frequently lost in PDAC (Kern, 2000) or in the PDAC-derived cell lines used in this study (data not shown). MicroRNA expression can be controlled by epigenetic alterations such as DNA hypermethylation and histone modification in human cancer cells (Weber et al., 2007). let-7a-3, a member of the let-7 microRNA family, is slightly methylated in normal human tissues but hypomethylated in some lung adenocarcinomas (Brueckner et al., 2007). Studies are underway to elucidate whether epigenetic modifications are responsible for let-7 repression in PDAC samples.
Previous work demonstrated that let-7 may directly control lung cancer and liver cancer-derived cell proliferation by negatively regulating the human K-ras gene (Johnson et al., 2005, 2007). Because activated K-ras is a key oncogene in PDAC, these previous findings strongly suggest that let-7 could be a candidate to inhibit PDAC proliferation. In this work, we demonstrate that restoring let-7 expression strongly affects the proliferation of a PDAC-derived cell line. This effect is independent of apoptosis induction, as previously demonstrated (our personal observation; and see Johnson et al., 2007). In addition, let-7 expression does not alter cell cycle progression of PDAC-derived cells (data not shown). We demonstrated here for the first time that let-7 targets K-ras in PDAC-derived cell lines. Consequently, MAPK activation was strongly impaired in PDAC-derived cells expressing let-7. These experiments strongly suggest that let-7 could be a valuable tool to impair PDAC progression. Efforts have been made to target K-ras as a therapeutic option to treat cancer, using either ribozymes, RNA interference (RNAi), or farnesyltransferase inhibitors such as R115777 (Fleming et al., 2005; Cengel et al., 2006). However, R115777 was demonstrated to be ineffective as monotherapy in advanced PDAC in a phase 2 trial (Macdonald et al., 2005).
Considering the data presented herein, we believe that restoring endogenous molecules known to regulate multiple oncogenic pathways, such as let-7 microRNA, will be a valuable tool in interventions aimed at treating PDAC. We demonstrated that delivering either plasmid-driven genes or synthetic oligonucleotides into tumors successfully inhibited PDAC progression in vivo, despite a low transfection efficiency (Carrere et al., 2005). Unfortunately, we failed to antagonize in vivo pancreatic tumor growth by the same approach used to deliver let-7. Such a discrepancy may be due, at least in part, to the molecule itself, and/or to the delivery method. In vitro, we found that let-7 slows cancer cell proliferation without affecting cell cycle progression or inducing cell death by apoptosis. The lack of therapeutic efficacy in the intratumoral administration of let-7 may reflect the absence of apoptosis induction and, consequently, of a strong antitumoral bystander effect essential for the inhibition of pancreatic tumor progression. In contrast, the use of microRNAs as potentially novel therapeutic targets in vivo remains trivial. Numerous groups have demonstrated the inhibition of microRNA function through antisense and microRNAs sponges (“antagomiRs”; Marquez and McCaffrey, 2008). However, efficient and specific delivery of these molecules either systemically or locally still limits the application of antagomiRs. Manipulation of microRNA levels still has therapeutic benefits to fight cancer. Si and colleagues demonstrated that the proliferation of breast cancer-derived cells treated ex vivo with oligonucleotide complementary to miR-21 was inhibited after injection into mammary pads of female nude mice (Si et al., 2007). In the same ex vivo settings, miR-29 was used to normalize the epigenetic status of non-small-cell lung cancer (NSLC), and to alter tumor progression in vivo (Fabbri et al., 2007). Let-7g, introduced in vitro into cancer cells by lentiviral vectors, was demonstrated to inhibit NSLC tumor initiation and development (Kumar et al., 2008). Although these models are fairly artificial, these reports demonstrate that modulation of microRNA can inhibit tumor growth.
Progress has been made for the delivery of microRNA into tumors in preclinical settings. MiR-34a transiently inhibits human colon cancer tumor progression when administered with atelocollagen for drug delivery (Tazawa et al., 2007). Using an established orthotopic mouse lung cancer model, Esquela-Kerscher and colleagues showed that intranasal let-7 administration reduces tumor formation in vivo in the lungs of animals expressing a G12D activating mutation for the K-ras oncogene (Esquela-Kerscher et al., 2008). We demonstrated that viral vectors, such as simian virus 40 (SV40) (Cordelier et al., 2007), are efficient for in vivo gene delivery into PDAC tumors. Clearly, these well-characterized vectors, together with lentiviral vectors, could be used in future to introduce mature microRNAs into PDAC-derived cells, to inhibit exponential in vitro and in vivo cell proliferation.
Footnotes
Acknowledgments
The authors thank Jean-José Maoret (molecular biology facility, IFR105, Toulouse, France) for expertise in RNA detection and quantification, and the BSL-3 facility for vector production (BiVIC platform, IFR150, Toulouse, France). Tumor samples were obtained through the Department of Pathology and the Tumor Banks of CHU de Bordeaux, CHU de Toulouse and Cancéropôle Grand Sud-Ouest (France). This work was supported by grants from INSERM, Région Midi-Pyrénées, the Association pour la Recherche contre le Cancer, and Cancéropôle Grand Sud-Ouest. J.T. was funded by the Institut National du Cancer and the Ligue Nationale contre le Cancer.
Author Disclosure Statement
The authors declare no competing interests.