
Abstract
RNA interference (RNAi) is rapidly becoming an important tool that is advancing research with therapeutic aims. It is necessary to develop efficient ways of guiding small interfering RNA (siRNA) to targeted tissues to induce an RNAi effect. Herein, we report on an active method for delivering macromolecular siRNA and its plasmid vector into the skin, using erbium:YAG (Er:YAG) laser pretreatment. The amount of siRNA transported through nude mouse skin was determined with an in vitro Franz diffusion assembly. Confocal laser scanning microscopy (CLSM) was used to examine the in vivo uptake of siRNA and the vector by the skin. The stratum corneum was partially ablated with the low-fluence laser. The results of in vitro experiments indicated a significant improvement in siRNA permeation with laser exposure, which showed a 2.4- to 10.2-fold increase compared with the nontreated group depending on the fluence used (1.2–1.7 J/cm2). A photomechanical wave generated by filtering the laser irradiation was sufficient to enhance siRNA permeation by 5-fold. CLSM revealed intense green fluorescence from naked siRNA within the epidermis and upper dermis after laser pretreatment, producing a 3.5-fold enhancement compared with the control. The green signal intensity in 1.7 J/cm2-treated skin was 4.2-fold higher than that in intact skin after the in vivo topical application of the siRNA expression vector. The increased signal was mainly in the dermis. This noninvasive, precisely controlled technique for siRNA therapy provides an efficient way to deliver siRNA and its vector into the skin.
Introduction
Although topical delivery has many potential benefits, the permeability of skin to macromolecules is extremely low because of the formidable barrier function of the stratum corneum (SC). Intradermal injections are a commonly used method to target genes or related products to the skin (Sawamura et al., 2002; Hickerson et al., 2008). A safe and noninvasive method is thus urgently needed to efficiently deliver them to the skin. Removal of the SC by mechanical abrasion, tape-stripping, or chemical treatment has been shown to significantly enhance the permeation of genes (Lin et al., 2001). However, these approaches may be limited because of the lack of control and reproducibility, as well as their potential to cause skin irritation (Hadgraft, 1999; Lee et al., 2006; Gómez et al., 2008). The erbium:yttrium–aluminum–garnet (Er:YAG) laser can precisely ablate the superficial SC with minimal residual thermal damage to the skin; it is currently used for skin rejuvenation, and resurfacing of rhytides, scars, and photodamage (Caniglia, 2004). We suggested that the Er:YAG laser at a low fluence can effectively enhance and precisely control the delivery of macromolecules via the skin (Fang et al., 2004b; Lee et al., 2006, 2008). The duration of the skin's exposure to the laser is in the range of nano- to microseconds, indicating a rapid enhancing effect.
The aim of the present work was to assess the feasibility of topical siRNA delivery using a pulse of an Er:YAG laser. This study used an in vitro Franz cell to evaluate the topical delivery of fluorescein-labeled siRNA by laser pretreatment. In the in vivo study, the distribution of siRNA and the expression of green fluorescent protein (GFP) by siRNA expression vector in nude mouse skin were monitored by confocal laser scanning microscopy (CLSM). To obtain pure photomechanical waves (PWs) generated by the laser and to exclude the SC ablation effect, a polystyrene lens was used to filter the laser light for comparison. Hence the mechanisms of siRNA permeation enhancement by the laser could be explored.
Materials and Methods
Materials
Fluorescein isothiocyanate (FITC)-labeled siRNA (5′-UGCUGACUCCAAAG-3′ and 3′-ACGACUGAGGUUUC-5′) was synthesized by Bio Basic (Markham, ON, Canada). The molecular mass of this siRNA was 9266 Da. FITC was purchased from Sigma-Aldrich (St. Louis, MO). The plasmid siRNA vector pRNAT-U6.2/Lenti encoding enhanced green fluorescent protein (GFP) was supplied by GenScript (Piscataway, NJ). Lipofectamine 2000 was obtained from Invitrogen (Carlsbad, CA). RNase-free water was purchased from PhytoTechnology Laboratories (Lenexa, KS). All other chemicals and reagents were of analytical grade and were used as received.
Er:YAG laser assembly
The Er:YAG laser (Continuum Biomedical, Santa Clara, CA) used here has a wavelength of 2940 nm and a pulse duration of 250 μsec. An articulated arm was used to deliver the laser beam onto the skin. Output energies of 0.45–0.65 J with a beam spot diameter of 7 mm achieved fluences of 1.2–1.7 J/cm2. The energy of the laser pulse was monitored with an energy meter (Nova display; Ophir-Spiricon, Jerusalem, Israel) before and after treatment. The protocol for laser application followed Guidelines for Laser Safety and Hazard Assessment by the Occupational Safety and Health Administration of the Department of Labor (Washington, DC; available at
In vitro topical delivery of siRNA and FITC
The diffusion cell used in this study was a Franz vertical diffusion assembly. A piece of excised nude mouse skin (ICR-Foxn1nu strain, 8 weeks old) was mounted on the receptor compartment with the SC side facing upward into the donor compartment. The laser hand-piece was located about 3.7 cm from the skin surface. After laser irradiation, the skin surface was wiped with a cotton wool swab several times. A black polystyrene target material was positioned on the skin for laser irradiation when evaluating the effect of the PWs. The receptor compartment (5.5 ml) was filled with a pH 7.4 citrate–phosphate buffer composed of RNase-free water. The donor compartment (0.3 ml) contained siRNA or FITC (2.08 μM) in pH 5.5 buffer and was occluded by paraffin film. The available diffusion area of the Franz cell was 0.785 cm2. The receptor was maintained at 37°C and stirred with a magnetic bar at 600 rpm. At appropriate intervals, 0.3-ml aliquots of receptor medium were withdrawn and immediately replaced with an equal volume of fresh medium. Samples were analyzed with a fluorescence spectrometer (F-2500; Hitachi, Tokyo, Japan) at λexcitation of 494 nm and λemission of 519 nm.
In vivo topical administration of siRNA
A glass cylinder similar to the donor compartment of the Franz cell with an available area of 0.785 cm2 was attached with glue to the back skin of a nude mouse. A laser pulse was applied to the skin area inside the cylinder. siRNA in pH 5.5 buffer (0.3 ml) at a concentration of 2.08 μM was then added to the cylinder for a 2-hr application. After 2 hr, the skin on which the vehicle had been applied was excised and wiped 10 times with a cotton cloth. All procedures were carried out in the dark to prevent the influence of ambient light. Skin samples were directly examined by CLSM. The full thickness was optically scanned at approximately 10-μm increments through the z axis of a Leica TCS SP2 confocal microscope (Leica Microsystems, Wetzlar, Germany). Optical excitation was carried out with a 488-nm argon laser beam, and the fluorescence emission was detected at 496–559 nm. The fluorescence signal was quantified with Leica QWin software.
Plasmid amplification
The pRNAT-U6.2/Lenti vector was transformed into Escherichia coli (strain DH5α)-competent cells by standard procedures and then amplified and purified by column chromatography. The purity of the plasmid was measured by determining the ratio of optical density at 260 nm to that at 280 nm (OD260/OD280, 1.85–1.90) as well as by electrophoresis in a 0.8% agarose gel. The purified plasmid was resuspended in RNase-free water and frozen in aliquots.
In vivo topical administration of siRNA expression vector
The procedure for in vivo topical administration of the siRNA expression vector was the same as that for naked siRNA administration. The 1-mg/ml plasmid in pH 5.5 buffer (0.3 ml) was used as the vehicle. The duration of administration was 6 hr. Punch biopsies were performed 24 hr after removing the pRNAT-U6.2/Lenti vector. For analysis of GFP expression, CLSM was used to scan the fluorescence signals of GFP at various skin depths. Optical excitation and emission were detected at the same wavelengths as for naked siRNA. In another experiment, Lipofectamine 2000 was complexed with plasmids (1 mg/ml) at a ratio of 1:1 as the donor vehicle.
Statistical analysis
The statistical analysis of differences between various treatments was performed by unpaired Student t test. A 0.05 level of probability was taken as the level of significance.
Results
In vitro topical delivery of siRNA and FITC
The cumulative percentage of siRNA in the receptor compartment as a function of time after laser pretreatment along with various energies is shown in Fig. 1. The cumulative percentage at the end of the experiment (12 hr) and enhancement ratio (ER) by the laser are given in Table 1. In the control group, nude mouse skin did not undergo any treatment to breach the barrier function of the SC before applying siRNA. As a result, siRNA showed low permeability without laser irradiation. Exposure to one pulse of the laser produced a significant increase (p < 0.05) in siRNA permeation by comparing the cumulative percentage at 12 hr. The Er:YAG laser irradiated a limited area of the skin surface, about 49% of the available diffusion area. Extrapolating the original cumulative percentage data of the laser-irradiated area to an area of 100% exposure (normalized cumulative percentage) resulted in an ER of 2.4–10.2 depending on differences in the fluence. The increase in laser fluence led to greater enhancement of siRNA permeation. However, the use of the highest fluence (1.7 J/cm2) did not further increase the siRNA percentage at 12 hr compared with a fluence of 1.4 J/cm2 (p > 0.05).
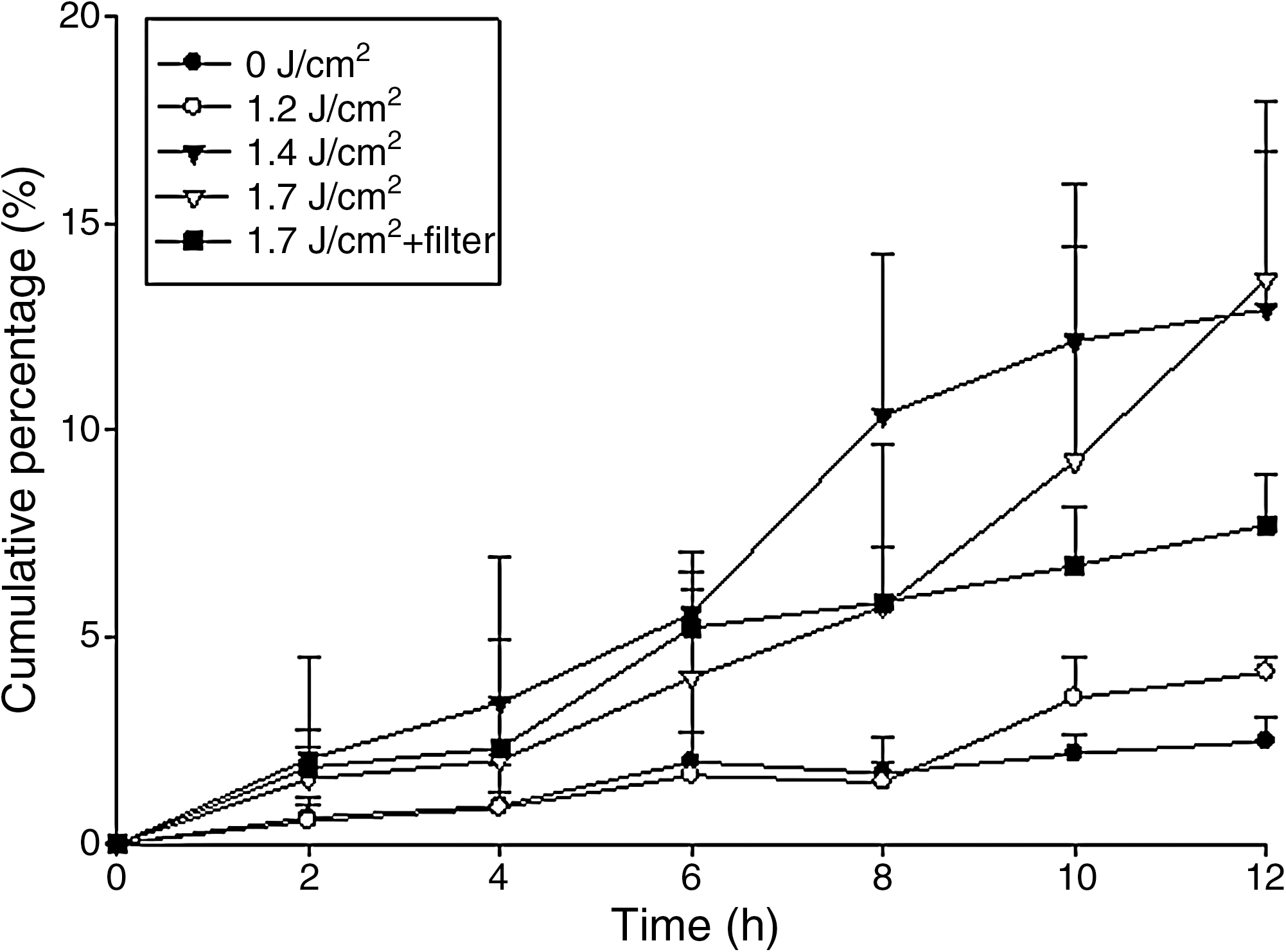
In vitro cumulative percentage (%)–time profiles of the topical delivery of small interfering RNA (siRNA) by erbium:yttrium–aluminum–garnet (Er:YAG) laser pretreatment of the skin at fluences of 1.2–1.7 J/cm2. Each value represents the mean ± SD (n = 4).
Each value represents the mean ± SD (n = 6).
Original cumulative percentage was calculated directly from the permeated percentage across partly laser-pretreated skin at 12 hr.
Normalized cumulative percentage was calculated from the cumulative percentage across fully laser-treated skin (100%), which was calibrated by cumulative percentage of control group (without laser pretreatment) at 12 hr.
Enhancement ratio (ER) was normalized cumulative percentage of laser-pretreated group/cumulative percentage of control group at 12 hr.
The small, hydrophilic FITC molecule was used as a model permeant in the in vitro permeation study for comparison with FITC-labeled siRNA as shown in Fig. 2. Table 2 demonstrates that the extent of passive transport without laser treatment was greater for the smaller species. FITC showed meaningful enhancement (p < 0.05) of its permeation until the laser fluence had increased to 1.4 J/cm2. Extrapolating the original data of the pretreated area to an area of 100% exposure resulted in 5.5- to 9.1-fold increases in ER in the case of FITC. The enhancement of FITC permeation was lower than that of siRNA.

In vitro cumulative percentage (%)–time profiles of the topical delivery of fluorescein isothiocyanate (FITC) by Er:YAG laser pretreatment of the skin at fluences of 1.2–1.7 J/cm2. Each value represents the mean ± SD (n = 4).
Each value represents the mean ± SD (n = 6).
Original cumulative percentage was calculated directly from the cumulative percentage across partly laser-pretreated skin at 12 hr.
Normalized cumulative percentage was calculated from the cumulative percentage across fully laser-treated skin (100%), which was calibrated by cumulative percentage of control group (without laser pretreatment) at 12 hr.
Enhancement ratio (ER) was normalized cumulative percentage of laser-pretreated group/cumulative percentage of control group at 12 hr.
—, No significant difference as compared with the original cumulative percentage of the control (0 J/cm2).
Effect of PWs on in vitro topical delivery of siRNA and FITC
To elucidate the mechanisms of the Er:YAG laser on topical siRNA delivery, PWs were generated by laser ablation of a polystyrene target and then launched into the skin. In this arrangement, laser irradiation was totally absorbed by the target so that only the PWs reached the skin (Lee and Doukas, 1999). This arrangement eliminated any ancillary effects associated with laser radiation. As shown in Fig. 1 and Table 1, it is clear that pure PWs at a fluence of 1.7 J/cm2 were sufficient to induce enhancement of siRNA permeation (p < 0.05) by 5.3-fold. However, this effect was lower than that generated by the laser without the polystyrene lens filter. On the other hand, PWs at 1.7 J/cm2 did not increase the permeation of FITC compared with the control group with no laser treatment, as depicted in Fig. 2 and Table 2.
In vivo topical administration of siRNA
The fluorescence and corresponding distribution of naked siRNA from a skin site either with or without exposure to laser pretreatment are shown in Fig. 3. The nude mouse skin thickness was optically scanned at approximately 10-μm increments for 16 fragments from the surface of the skin (Fig. 3, left to right, top to bottom). Because the thicknesses of nude mouse SC and epidermis are about 11 and 18 μm, respectively (Lee et al., 2002), the first three fragments roughly characterized the SC and epidermal parts. The other parts below the first three fragments were dermal layers. No or negligible fluorescence signal was observed in the profiles of skin treated with blank buffer without siRNA (data not shown). Topical application of naked siRNA without laser irradiation produced a weak fluorescence signal in the lower dermis. No evidence of a green signal was observed in the SC, epidermis, or upper dermis. Compared with the control, increased green fluorescence in skin treated with the laser could clearly be seen with an intense signal as shown in Fig. 3B and C. There was a broad band of fluorescence extending from the SC into the upper dermis. The signal faded out in the lower dermal section.

Confocal laser scanning microscopic (CLSM) micrographs of nude mouse skin after the in vivo topical administration of siRNA via the skin by laser pretreatment at (
The fluorescence intensity depended on the concentration of FITC-labeled siRNA in the tissue. Quantification of the signal of the sum from 16 fragments is summarized in Table 3. The increase in fluorescence intensity relative to the control was statistically significant (p < 0.05) for the laser-treated group. The Er:YAG laser at 1.4 J/cm2 caused an approximately 3.5-fold enhancement of siRNA permeation. There was no significant difference (p > 0.05) between the intensity of the 1.4- and 1.7-J/cm2 treatments.
Each value represents the mean ± SD (n = 3).
Enhancement ratio (ER) was intensity of laser-pretreated group/intensity of control group.
In vivo topical administration of siRNA expression vector
Besides naked siRNA, we also attempted to optimize the present technique for delivering an siRNA expression plasmid vector into the skin tissue. This vector carried a GFP marker under the control of a cytomegalovirus (CMV) promoter to allow tracking of transfection efficiency. Expression of the reporter gene encoding GFP in skin was assessed by CLSM. As shown in Fig. 4A there was almost no fluorescence signal in nontreated skin except for weak fluorescence detected at the bottom of the dermis. Fluences of 1.4 and 1.7 J/cm2 both increased the green signal in the skin as depicted in Fig. 4B and C. A fluence of 1.4 J/cm2 produced a paler signal compared with that of 1.7-J/cm2 fluence. The energy at 1.7 J/cm2 enhanced GFP expression by 4.2-fold. The expression with 1.7 J/cm2 became evident in the dermis, but not in the SC or epidermis (Fig. 4C). The effects of PWs on GFP expression were also tested as shown in Fig. 4D. The PWs did not increase the intensity compared with the control.

CLSM micrographs of nude mouse skin after the in vivo topical administration of an siRNA expression vector via the skin by laser pretreatment at (
The siRNA expression vector complexed with Lipofectamine was also evaluated on in vivo nude mouse skin. A lower fluence of 1.4 J/cm2 was used here because Lipofectamine may be sufficient to enhance the transfection ability. Figure 5A and B shows fluorescence images obtained from skin biopsies of a control site and a laser-exposed site, respectively. For the untreated site (Fig. 5A), the fluorescence was mainly from the dermis. The intensity from the complex was higher compared with that of the vector alone (28.45 vs. 17.30), although the difference was not significant (p > 0.05) (Table 3). GFP expression of the complex permeated into laser-treated skin was extensively distributed in the epidermis and dermis. Exposure to a pulse of the laser enhanced the expression level by a factor of up to 1.6 over that of the untreated group.
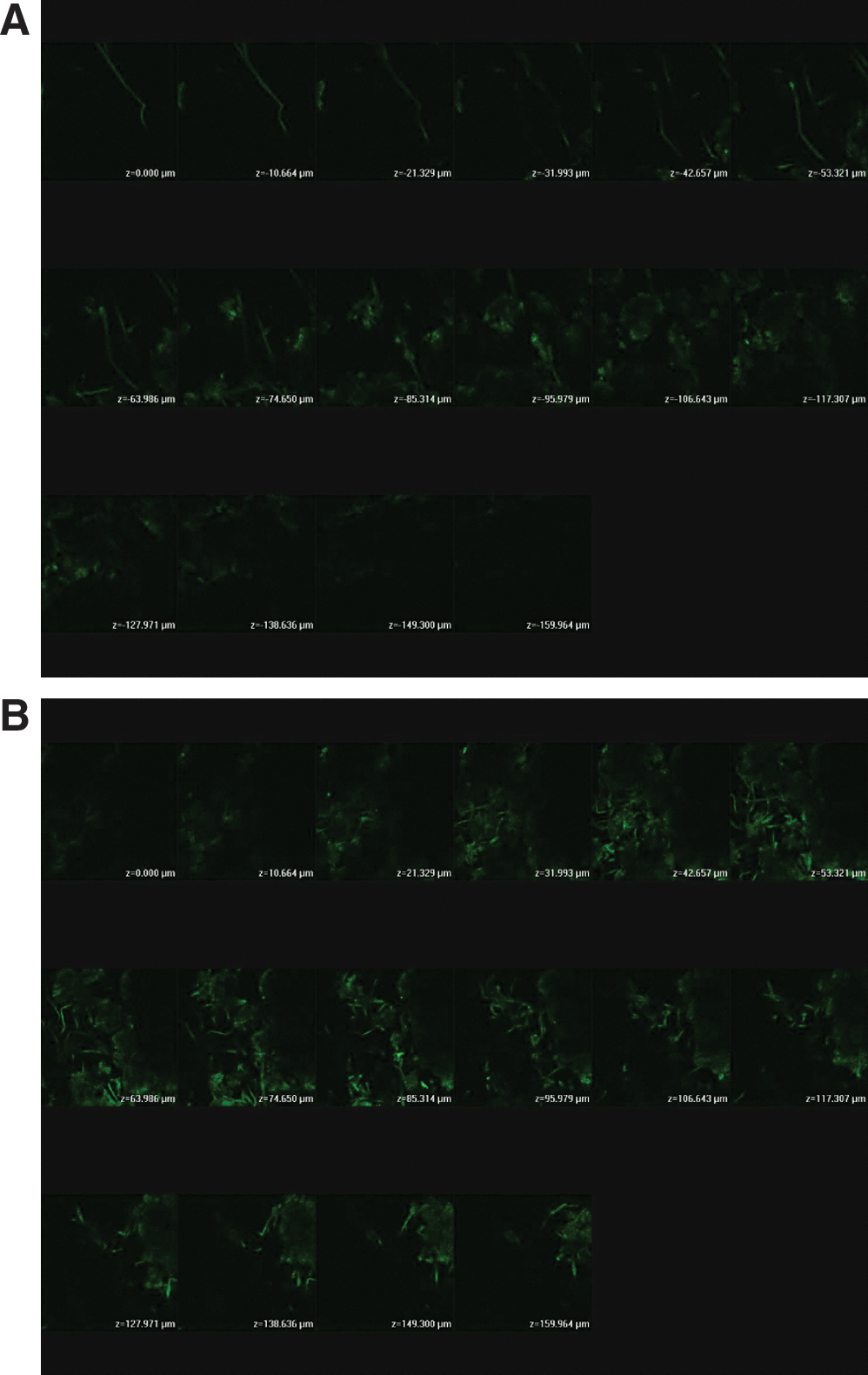
CLSM micrographs of nude mouse skin after the in vivo topical administration of an siRNA expression vector and Lipofectamine complex via the skin by laser pretreatment at (
Discussion
The discovery of siRNA has opened up wide perspectives in therapeutics for treating many diseases (Fattal and Bochot, 2008). The efficiency of RNAi and its limited side effects have made this technique an attractive alternative to the use of antisense oligonucleotides and ribozymes for therapies based on inhibitors of target genes (Golzio et al., 2007). The skin represents an attractive target for nucleic acid-based therapies owing to its ease of accessibility and the ability to locally deliver therapeutic agents. The local injection of siRNA may cause skin irritation and infection. A noninvasive method for topical application via the skin is strongly needed. In the present report, we show for the first time successful siRNA permeation and expression in skin, using Er:YAG laser pretreatment of the SC. The results presented indicate the feasibility and usefulness of the laser to enhance skin permeability.
To be of clinical utility, the removal of the SC must be relatively safe and without adverse effects. The Er:YAG laser tested in this study used lower energies than those used in clinical situations for resurfacing aims. Only a limited part of the SC was removed by these low laser fluences. As the SC rapidly regenerates, the skin should recover after laser treatment. In our previous studies, the skin recovered to normal status within 3 days, as evaluated by SC thickness and transepidermal water loss (Lee et al., 2001, 2004). The microsecond range of the pulse duration used is shorter than the thermal relaxation time of the skin (1 msec), which thus avoids thermal injury to the skin (Gómez et al., 2008). A previous investigation (Singer et al., 2005) also indicated that the Er:YAG laser at a fluence of 3.5 J/cm2 showed no evidence of damage or infection to pig skin.
The permeability of siRNA into or across intact skin is generally low, making it difficult to achieve the desired therapeutic benefits. The Er:YAG laser is an ablative tool for the SC with the capability of precise control. The low-fluence laser was effective in promoting the skin permeability of siRNA at all energies studied (1.2–1.7 J/cm2). Histological and structural alterations of the skin may be responsible for this enhancement. With ablation, laser radiation causes decomposition of the target material into smaller fragments, which move away from the surface of the target at supersonic speed (Doukas and Kollias, 2004). The laser fluence was found to play an important role in controlling siRNA and FITC permeation. As demonstrated in our previous study (Fang et al., 2004a), the etched thickness of the SC layer after laser ablation appeared to be proportional to the treatment fluence. siRNA permeating into the receptor compartment at a fluence of 1.7 J/cm2 did not exceed that at 1.4 J/cm2. This indicates that mechanisms other than SC ablation may have affected the increasing permeation by the laser. The permeant is first partitioned into the SC, after which it progresses through the skin to deeper layers. Although removal of a part of the SC can reduce the inherent barrier function of the skin, the partitioning of the permeant into the SC should decrease after its partial removal, which results in an offset effect (Lee et al., 2002). This phenomenon was not observed for FITC, a small molecule. This suggests that SC ablation, rather than the offset mechanism, predominated FITC permeation by the laser.
It is clear that the threshold of laser fluence for inducing meaningful enhancement of FITC permeation is higher than that of siRNA. A fluence of 1.2 J/cm2 was insufficient to induce significant enhancement of FITC delivery. Moreover, the ER value of FITC permeation by the laser was lower compared with siRNA at the same fluence. This may have been due to the easier penetration of the smaller molecule across the skin in a passive manner. Thus the effect of the partial ablation of the SC by the laser is limited to promoting FITC permeation. This suggests that the more important barrier property of the SC acts against the passing of larger molecules.
Three mechanisms, including direct ablation, optical breakdown by PWs, and a photothermal effect, are involved in laser–tissue interactions (Doukas and Kollias, 2004). The photothermal effect can be ruled out as a mechanism of the Er:YAG laser because this laser emits light with a minimal residual thermal effect. PWs generated by lasers are broadband (with a fast rise time), unipolar, compressive waves (Doukas and Flotte, 1996). It is reasonable to assume that the Er:YAG laser produced PWs that affected topical siRNA delivery. In the experiment to induce PWs, laser radiation was totally absorbed by the target and did not reach the SC. Hence the permeability of the skin was strictly the result of PWs impinging on the SC. Pure PWs produced a 5.3-fold increase in siRNA permeation compared with the control group. We previously indicated, on the basis of electron microscopic evidence, that PWs widened the intercellular junctions of the SC (Fang et al., 2004b; Lee et al., 2008), thus reducing the barrier function. Previous studies reported that PWs cause disruption of lipids by inducing the transient formation of a continuous permeable lacunar system within the SC (Lee et al., 2001). This effect can facilitate drug transport into viable skin (Doukas and Kollias, 2004).
Although the PWs were sufficient to enhance siRNA delivery via the skin, this effect was still less than that achieved by direct laser exposure. This suggests that the SC ablation effect is still the main force responsible for siRNA permeation by the laser. As indicated from our results, there was no enhancement of FITC permeation by PWs compared with the control. This may have been because preexisting lacunae in the lipid bilayers of the SC were large enough for smaller molecules to penetrate in the absence of laser exposure. Further enlargement of the lacunae by PWs did not affect the transport of FITC.
An approximately 3.5-fold increase in siRNA distributed in the skin, as determined by CLSM examination, was detected with Er:YAG laser pretreatment of the skin. Enhancement of the siRNA distribution by fluences of 1.4 and 1.7 J/cm2 was approximate, confirming the offset hypothesis elucidated in the in vitro experiment. The laser greatly increased the distribution of naked siRNA in the SC, epidermis, and upper dermis but not in the lower dermis. It appears that the SC is not the sole significant contributor to resistance against siRNA. Viable skin appeared to be responsible for some part of the resistance. Of course, the SC contribution as an important barrier for siRNA permeation cannot be neglected.
There are two basic options for introducing siRNA into mammalian cells. The first employs synthetic siRNA in a naked form. The second option deploys a DNA-dependent expression vector for siRNA. The expression system may encode short hairpin RNA (shRNA), which is subsequently processed by cells to generate siRNA (Jiang et al., 2004). The advantage of the siRNA expression system is that the RNAi effects last much longer when plasmid-based siRNA is used (Inoue et al., 2006). However, plasmid diffusion to the target tissue is expected to be extremely low because of its relatively larger size compared with naked siRNA. It is important to enhance the transfection of the plasmid in the skin by external forces. As shown in the results of the present work, the Er:YAG laser indeed strongly stimulated skin permeation and cell uptake of the plasmid vector. The vector should be included in the cytoplasm to emit the GFP signal. Contrary to naked siRNA, limited success was achieved in enhancing vector permeation and transfection at a fluence of 1.4 J/cm2. This suggests that a higher energy is required for the vector to break through the permeation barrier. A fluence of 1.7 J/cm2 significantly enhanced siRNA expression in the entire dermis. This observation somewhat differs from the results for naked siRNA, which showed a significant enhancement in the epidermis and upper dermis. In this regard, there may be different choices for targeting specific sites by laser application with RNAi-based therapeutics.
The fact that most classes of macromolecules have great difficulty in reaching intracellular targets is a severe limitation (Høgset et al., 2004). Our results of in vivo vector permeation suggest that the low-fluence laser not only overcomes the skin barrier but also actively drives the applied permeant into cells by means of a short pulse. There are two possible ways in which the laser promotes gene transfer: acceleration of exogenous genes and/or disruption of the plasma membrane by PWs (Ogura et al., 2004). PWs with a short rise time are effective in permeabilizing cell membranes. There are indications that membrane permeabilization is mediated by membrane proteins (aquaporins) that form a water-filled channel and are present in most mammalian cells (Doukas and Kollias, 2004). Our CLSM results suggest that PWs alone did not affect the fluorescence expression in the skin (Fig. 4D). The Er:YAG laser should first ablate some SC layers to drive the vector into viable skin, and then the PWs drive the vector into cells. That is, the effect of pure PWs on the SC structure was insufficient to force the vector into viable skin.
The main obstacle to using siRNA for in vivo therapy remains the efficiency of intracellular delivery (Xie et al., 2006; Golzio et al., 2007). Intracellular targeting methods for skin include tape-stripping, gene gun, intradermal injection, and depilatory technology (Peachman et al., 2003). However, skin stripping with tape is painful and poorly reproducible (Mikszta et al., 2002). The force of air pressure with the gene gun causes mild damage to the skin (Peachman et al., 2003; Barfoed et al., 2004). Intradermal injections are an invasive method. The recovery of skin to normal status after application of depilatory cream requires 5–7 days (Kanikkannan et al., 1999). Hence the Er:YAG laser may be an attractive tool for targeted transfer into cells because of its noninvasive ability and high level of control. Further study is needed to elucidate the detailed mechanisms.
When lipids form complexes with siRNA to produce amorphous particles, they are known as lipoplexes. Lipoplexes are formed by mixing siRNA with most commercial transfection agents, such as Lipofectamine. We showed that Lipofectamine did little to increase GFP expression of the siRNA vector. A further increase in the fluorescence signal was achieved after laser irradiation. The ER value of the vector with or without Lipofectamine with the laser was the same, indicating that the effect of the laser on vector permeation still remained after incorporating a transfection agent.
Conclusions
Disease-specific siRNA can provide future therapeutics for treating cutaneous disorders. One potential problem for siRNA delivery is the dose required to elicit the desired effect, which would be prohibitively expensive. In the present study, we demonstrated the potent efficacy of a low-fluence laser at enhancing the skin delivery of siRNA and its expression vector, thus possibly reducing the siRNA dose and concomitant cost required. The emerging field of RNAi therapy also relies on siRNA that must be internalized into cells. A single pulse of the laser with PWs was sufficient to permeabilize cells and allowed increased expression of siRNA in the cells. The results shown in the present work describe the first use of a laser to breach the skin barrier and deliver siRNA topically, suggesting significant clinical and practical advantages. This basic study encourages further investigations of siRNA permeation and expression by a low-fluence laser.
Footnotes
Acknowledgments
The authors thank the National Science Council, Taiwan for financial support (NSC-96-2628-B-182-002-MY3).
Author Disclosure Statement
No competing financial interests exist for any of the authors.