
Abstract
The CH-296 recombinant fragment of human fibronectin is essential for murine leukemia virus (MLV)-derived retroviral transduction of CD34+ cells for the purpose of stem cell gene therapy. Although the major effect of CH-296 is colocalization of the MLV-derived retrovirus and target cells at specific adhesion domains of CH-296 mediated by integrins expressed on CD34+ cells, the precise roles of the integrins are unclear. We examined the kinetics of integrin expression on CD34+ cells during the course of MLV-derived retrovirus-mediated gene transduction with CH-296. Flow cytometry revealed that the levels of both very late activation protein (VLA)-4 and VLA-5 on CD34+ cells freshly isolated from cord blood were insufficient for effective MLV-derived retroviral transduction. However, increases were achieved during culture for preinduction and MLV-derived retrovirus-mediated gene transduction in the presence of a cocktail of cytokines. In addition, we confirmed by using specific antibodies that inhibition of the cell adhesion mediated by the integrins significantly reduced transduction efficiency, indicating that integrin expression is indeed important for CH-296-based MLV-derived retroviral transduction. Only a few cytokines are capable of inducing integrin expression, and stem cell factor plus thrombopoietin was found to be the minimal combination that was sufficient for effective transduction of an MLV-derived retrovirus based on CH-296. Our findings should be useful for improving the culture conditions for CH-296-based MLV-derived retroviral transduction in stem cell gene therapy.
Introduction
Stem cell gene therapy in which a defective gene in some hematopoietic stem and progenitor cells (HSPCs) is replaced with a functional copy can reconstitute the immune system and is therefore expected to be an effective treatment for inherited immunodeficiencies. Indeed, clinical trials in children with severe combined immunodeficiency (SCID), adenosine deaminase (ADA) deficiency, and chronic granulomatous disease (CGD) have been successful (Onodera et al., 1998; Cavazzana-Calvo et al., 2000; Ariga et al., 2001; Ott et al., 2006).
Murine leukemia virus (MLV)-derived retroviral vectors are used to place a functional copy of a gene in HSPCs. Although MLV-derived retroviruses have the ability to introduce and express genes stably in the host cell genome, applicable protocols in humans have been limited because of the low gene transfer efficiency achieved (Bodine et al., 1998). However, the development and application of a truncated form of recombinant human fibronectin (CH-296 [RetroNectin]; Takara Bio, Otsu, Shiga, Japan) have significantly improved the efficiency of transduction of MLV-derived retroviral vectors into reconstituting hematopoietic stem cells (Kimizuka et al., 1991).
CH-296 is a recombinant fragment of human fibronectin composed of the cell-binding domain (C domain), heparin-binding domain II (H domain), and CS1 site. The H domain was found to bind MLV-derived retroviral particles, and the RGD sequence in the C domain and the LDV sequences in the CS1 site are recognized by a variety of cell types through integrins α5β1 (VLA-5) and α4β1 (VLA-4), respectively. Consequently, CH-296 is capable of enhancing MLV-derived retrovirus-mediated gene transduction by binding to both MLV-derived retroviral particles and target cells that express VLA-5 and/or VLA-4 (Hanenberg et al., 1996, 1997).
The practical effects of CH-296 on MLV-derived retrovirus-mediated gene transduction into HSPCs have been well established in both basic biological experiments and clinical trials of stem cell gene therapy using MLV-derived retroviral vectors (Ariga et al., 2001). However, the actual effects of integrins expressed on HSPCs on MLV-derived retrovirus-mediated gene transduction with CH-296 have not been well characterized. Although differences in integrin expression on HSPCs should significantly affect the efficiency of MLV-derived retroviral gene transduction and thus greatly influence the success of clinical trials of stem cell gene therapy, the details remain unclear, and investigating the kinetics of integrin expression on HSPCs during their manipulation for gene transduction should help to improve stem cell gene therapy.
In this paper we present evidence that CD34+ cells freshly isolated from cord blood express only small amounts of integrins, but that they accumulate integrins during culture for preinduction and MLV-derived retrovirus-mediated gene transduction in the presence of a cocktail of cytokines. We also confirmed the effect of integrin levels on CH-296-based MLV-derived retrovirus-mediated gene transduction in human CD34+ cells and attempted to identify the minimal combination of cytokines that would allow effective integrin expression sufficient for MLV-derived retrovirus-mediated gene transduction.
Materials and Methods
Reagents, cells, and cell culture
The cord blood was distributed by the Tokyo Cord Blood Bank (Tokyo, Japan) and had originally been collected and stored for stem cell transplantation. Stocks inappropriate for transplantation because they contained too few cells were distributed for research use with the informed consent of the donor and the permission of the ethics committee of the Tokyo Cord Blood Bank. All the experiments in which cord blood was used in this study were performed with the approval of the local ethics committee. After isolating mononuclear cells from the cord blood by density gradient centrifugation with Ficoll-Hypaque, immunomagnetic enrichment of CD34+ cells was performed with a magnetic-activated cell-sorting (MACS) system according to the instructions of the manufacturer (Miltenyi Biotec, Bergisch Gladbach, Germany).
X-VIVO 10 (Lonza Walkersville, Walkersville, MD) containing 1% human albumin and a cytokine cocktail consisting of stem cell factor (SCF, 50 ng/ml), thrombopoietin (TPO, 50 ng/ml), Flt3 ligand (FL, 300 ng/ml), interleukin-6 (IL-6, 100 ng/ml), and soluble IL-6 receptor (sIL-6R, 500 ng/ml) was used for pretransduction and maintenance of the CD34+ cells ex vivo. In some experiments the cytokines were used alone or in combinations of fewer cytokines than in the complete cocktail, as indicated in the figures. All the cytokines used in this study were purchased from PeproTech EC (London, UK).
Viral supernatants were obtained from P13/DNEGFP cells developed from PG13 packaging cells transfected with pGCDNsap (DNEGFP), as described previously (Suzuki et al., 2002), and used as the MLV-derived retroviral vector for enhanced green fluorescent protein (EGFP) expression. PG13 is a retrovirus-packaging cell line derived from TK-NIH/3T3 cells and stably expresses the gibbon ape leukemia virus (GALV) envelope glycoprotein of this virus for pseudotyping of the MLV vectors that are transfected in these cells. The viral titer was determined with HT-1080 cells (American Type Culture Collection [ATCC], Manassas, VA).
The monoclonal antibodies (mAbs) used for immunofluorescence analysis were as follows: phycoerythrin (PE)-conjugated anti-CD49d and anti-CD49e from BD Biosciences (San Jose, CA), and PE-conjugated anti-CD29 and PE–cyanine 5 (PC-5)-conjugated anti-CD34 from Beckman Coulter (Fullerton, CA).
Transduction of CD34+ cells
CD34+ cells isolated from cord blood were suspended at a concentration of 1 × 106 cells/ml in X-VIVO 10 and cultured for 48 hr, as preinduction, in the presence of the cytokines described previously. To transduce CD34+ cells with the EGFP expression vector, 1 × 105 cells were resuspended in 1 ml of diluted P13/DNEGFP supernatant supplemented with the cytokines used for preinduction and plated in retronectin-precoated 35-mm dishes. After incubation for 12 hr the cells were collected by centrifugation (200 × g for 5 min) and washed once with X-VIVO 10 medium. The cells were then cultured for 48 hr in X-VIVO 10 supplemented with the same cytokines as used for induction. In this experiment, only a single induction was performed to identify the effect of the reagents. All the transduction assays in this study were performed at a multiplicity of infection (MOI) of 1.
In some experiments, purified mAbs against CD49d and/or CD49e (1 μg/ml, Beckman Coulter; both capable of inhibiting the cell adhesion mediated by their target molecule) were added to the culture 10 min before MLV-derived retroviral induction to assess the effect of inhibition of integrin-mediated cell adhesion.
The colony assay was performed with Methocult GF+H4435 (StemCell Technologies, Northampton, UK). After MLV-derived retroviral transduction, 3 × 102 CD34+ cells were suspended on 1 ml of Methocult and plated in 35-mm dishes. The number of colonies comprising more than 50 cells, including erythroid colonies, was scored after incubation for 21 days. The experiments were performed in triplicate, and colony numbers (means ± SD) are indicated in Table 1.
Results of Colony Assays
Abbreviations: IL-6, interleukin 6; SCF, stem cell factor; sIL-6R, soluble IL-6 receptor; TPO, thrombopoietin.
Immunofluorescence study
After cultivation periods the cells were stained with fluorescence-labeled mAbs and analyzed by flow cytometry as described previously (Kiyokawa et al., 1990). A three-color immunofluorescence study was performed with a combination of PE, PC-5, and GFP introduced by MLV-derived retroviral vectors. Experiments were performed in triplicate, and means and standard deviations of the cell counts are indicated in the figures.
Results
Expression of integrins on CD34+ cells on the course of MLV-derived retroviral transduction
As shown in Fig. 1, we first confirmed that the recombinant fibronectin fragment CH-296 is effective for transduction of CD34+ cells with an MLV-derived retroviral vector for GFP expression. After transduction with a single exposure to fresh viral supernatant, almost no GFP expression was observed in CD34+ cells in the absence of CH-296, whereas when the dish precoated with CH-296 was used, about 20% of the cells (15.6 to 29.0%; mean, 21.0 ± 3.1%) expressed GFP in repeated experiments (Fig. 1), indicating that CH-296 was indeed effective for MLV-derived retroviral transduction of CD34+ cells.
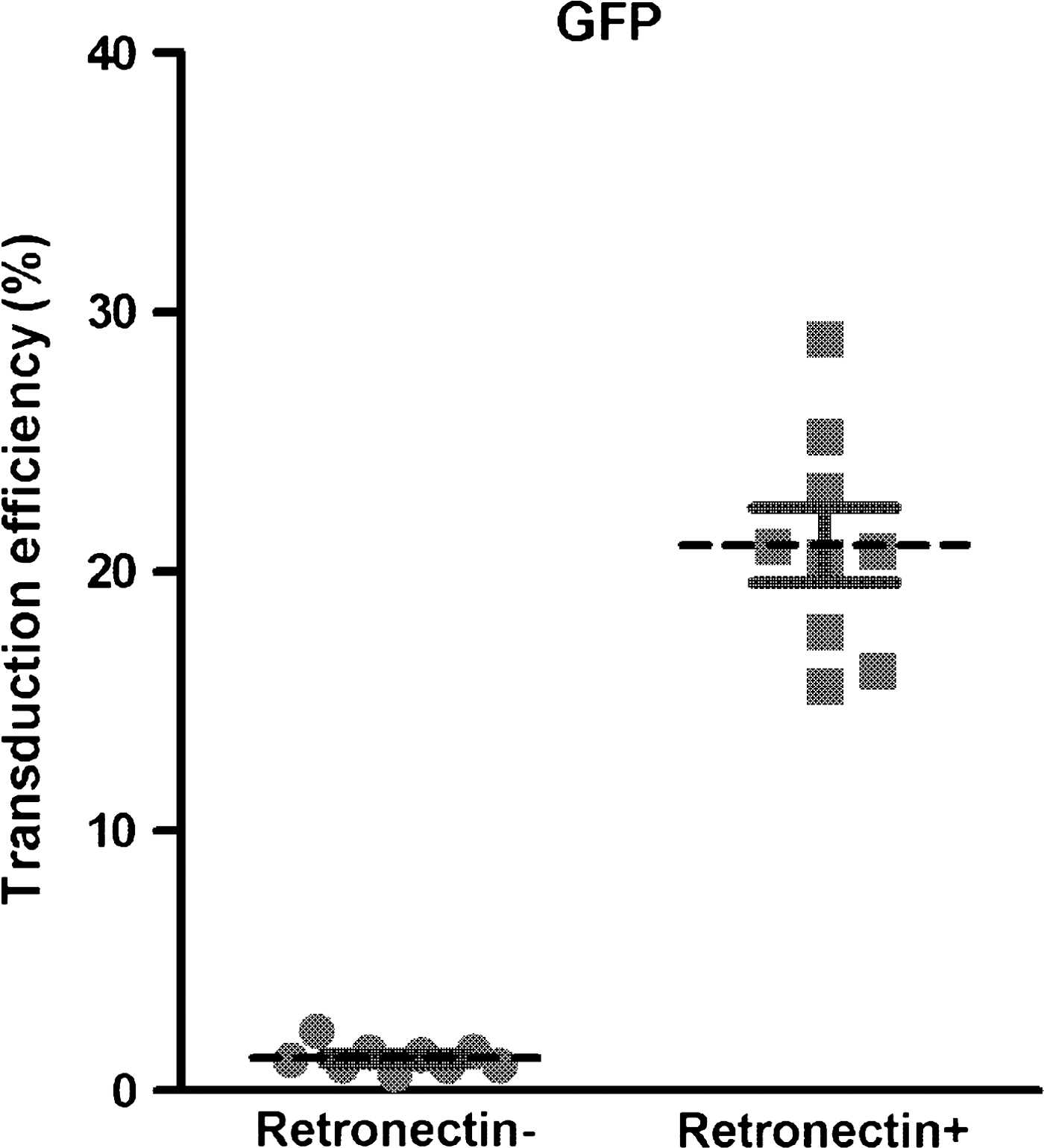
Effect of CH-296 (RetroNectin) on murine leukemia virus (MLV)-derived retrovirus-mediated green fluorescent protein (GFP) gene introduction. CD34+ cells were transduced with an MLV-derived retroviral vector for GFP expression in the absence (left) or presence (right) of the recombinant fibronectin fragment CH-296, as described in Materials and Methods, by means of a single exposure. Subsequent GFP expression was tested by flow cytometry. Results are expressed as means ± SD.
We therefore tested CD34+ cells for integrin expression during MLV-derived retroviral transduction. As shown in Fig. 2A, when freshly isolated CD34+ cells were tested for expression of CD49d, a component of VLA-4, the percentage of positive cells was already about 80% (67.6 to 96.0%; mean, 79.1 ± 6.0%), but it rose to approximately 100% during culture with the combination of cytokines for MLV-derived retroviral transduction. Moreover, comparison of the mean fluorescence intensity of CD49d in the positive cells showed a significant increase after the culture periods (Fig. 2B). Testing CD34+ cells for expression of CD49e, a component of VLA-5, revealed a low positive rate and mean fluorescence intensity when first isolated, but they both increased significantly after cultivation for transduction. A significant level of expression of CD29, another component of both VLA-4 and VLA-5, was found in freshly isolated CD34+ cells, but the mean fluorescence intensity still increased in the course of cultivation for MLV-derived retroviral transduction. The data indicate that the levels of VLA-4 and VLA-5 in freshly isolated CD34+ cells were not always high, but that the intensity of expression of both molecules significantly increased in the course of MLV-derived retroviral transduction.
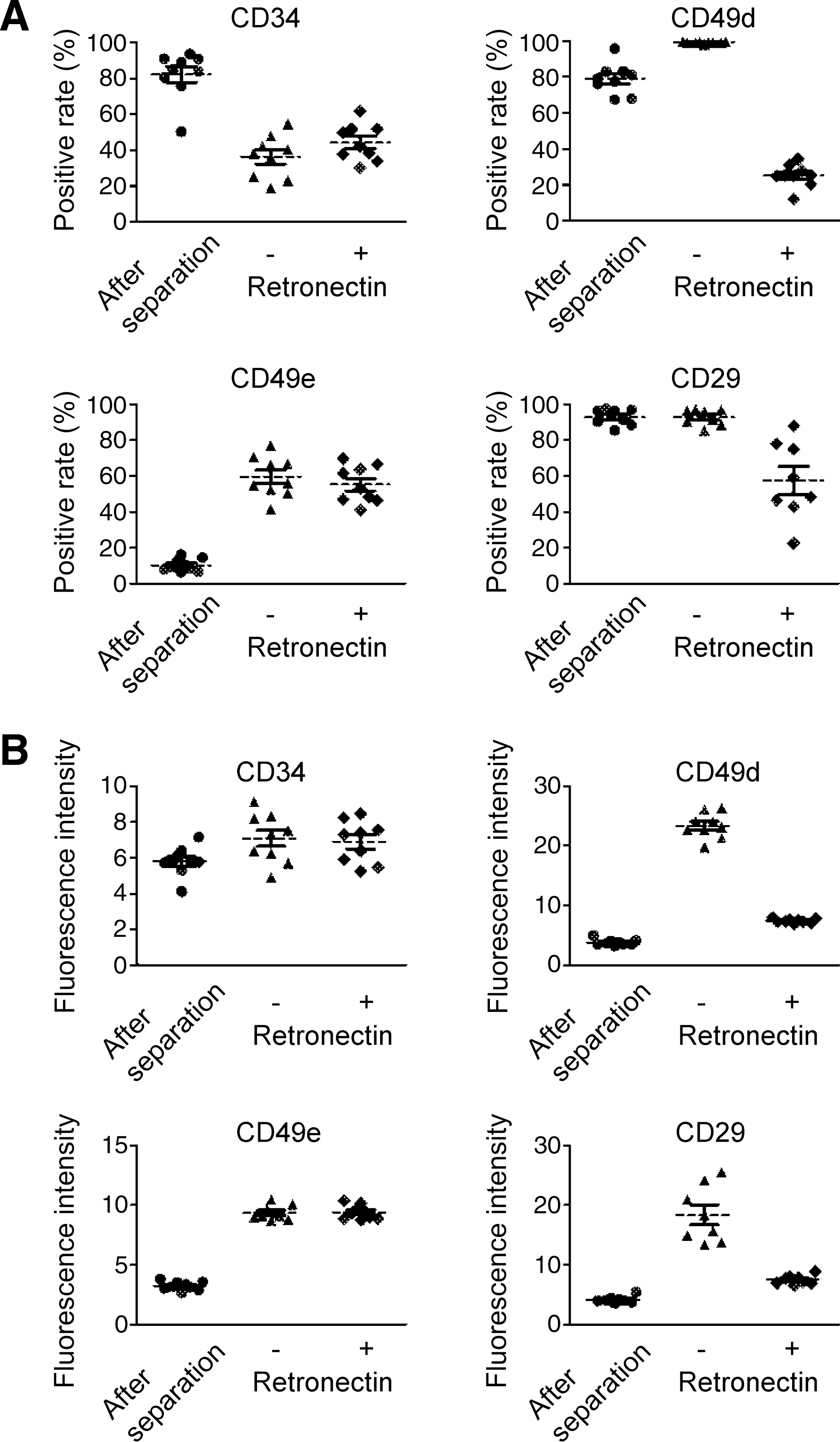
Expression of VLA-4 and VLA-5 in the course of MLV-derived retroviral transduction of CD34+ cells. Expression of VLA-4 and VLA-5 on CD34+ cells before (left) and after ex vivo culture for MLV-derived retroviral transduction in the absence (middle) or presence (right) of CH-296 was examined by flow cytometry, using specific antibodies as indicated. (
Interestingly, although integrin expression was significantly increased by cultivation with the cytokine cocktail, it was significantly reduced in the presence of CH-296 (Fig. 2). The degree of reduction was more significant for CD49d expression, but the decrease in CD49e expression was less marked (Fig. 2).
Role of integrins in CH-296-based MLV-derived retroviral transduction of CD34+ cells
We next investigated whether the integrins are indeed involved in the CH-296-based MLV-derived retroviral transduction of CD34+ cells. When anti-CD49d antibody, which inhibits cell adhesion via CD49d, was added to the culture, transduction efficiency was significantly reduced (Fig. 3). The anti-CD49e antibody similarly reduced the transduction efficiency (Fig. 3), and when both antibodies were added at the same time, a synergistic effect in reducing transduction efficiency was observed. The data indicate that the integrins expressed on CD34+ cells are indeed important to CH-296-based MLV-derived retroviral transduction.
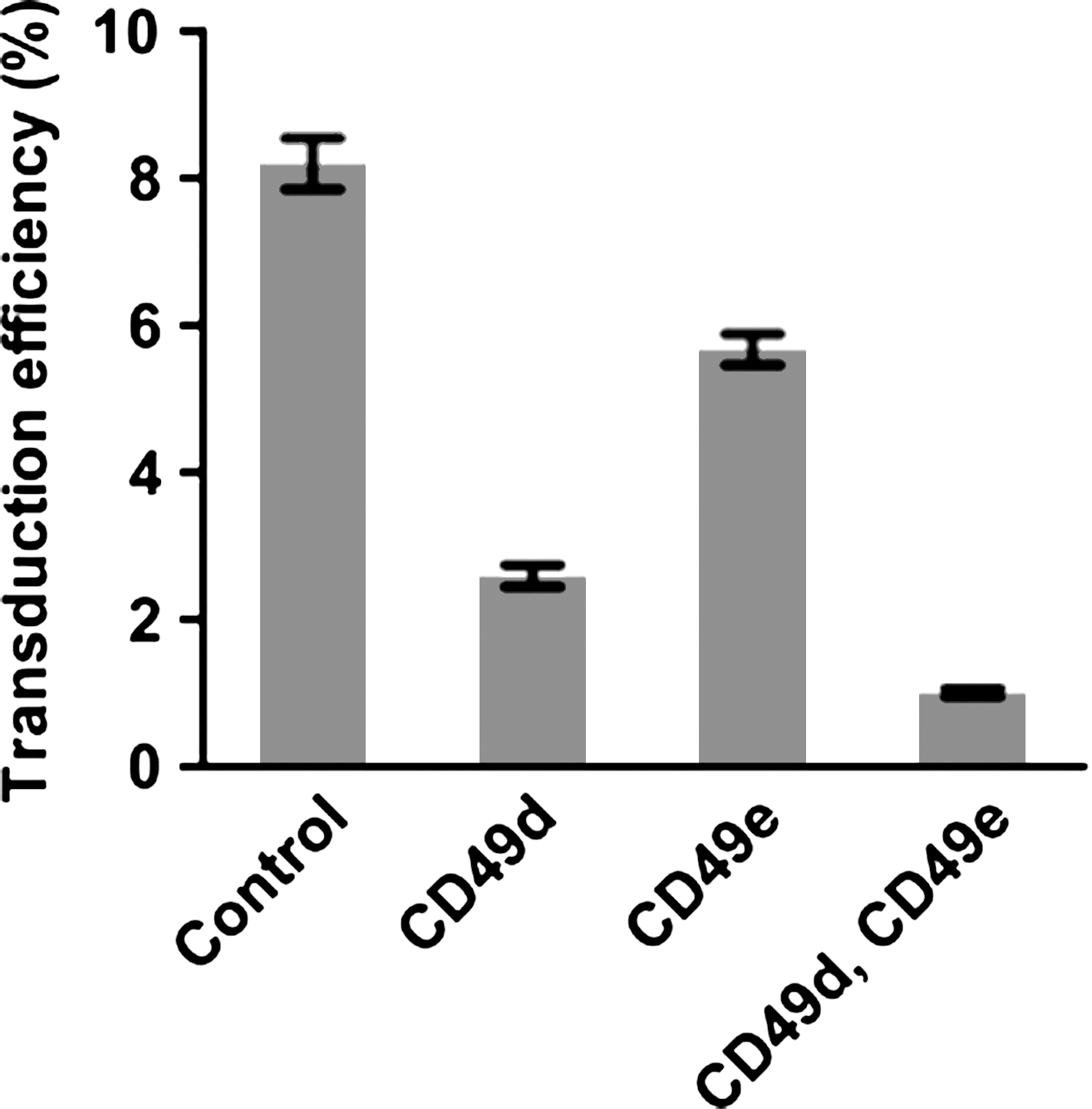
Effect of inhibition of integrin-mediated attachment to CH-296 on the efficiency of transduction of CD34+ cells with MLV-derived retroviral vector. CD34+ cells were transduced with MLV-derived retroviral vector for GFP expression in the presence of CH-296 as described in Fig. 1. Antibody against CD49d, CD49e, or both was added to the cells before exposure to the virus, and their effects were examined as described in Materials and Methods.
Effect of cytokines on integrin levels on CD34+ cells
We next attempted to identify the cytokines in the cocktail that were responsible for inducing expression of the integrins during the course of MLV-derived retroviral transduction. Both the positive rates and the fluorescence intensity measurements showed that SCF, TPO, and FL each increased expression of CD49d, but their effects were limited (Fig. 4A and B). SCF was the most effective of the three cytokines in enhancing CD49d expression. Exposure to a combination of IL-6 and sIL-6R had a weaker effect on CD49d expression than SCF, TPO, and FL alone (Fig. 4A and B). When SCF and TPO were combined, however, the level of CD49d expression increased significantly, and as shown in Fig. 4A and B, the positive rate and fluorescence intensity of CD49d on CD34+ cells cultured with the combination of SCF and TPO were almost the same as when cultured with the complete cocktail of five factors. In addition, the cytokines enhanced CD49e expression to the same extent as the cytokine-induced expression of CD49d (Fig. 4C and D).

Effect of cytokines on CD49d and CD49e expression on CD34+ cells. CD34+ cells were cultured in the presence of the cytokines indicated, and the expression of CD49d and CD49e was examined as described in Fig. 2. (
We then examined the effect of the cytokines on transduction efficiency with the GFP-expressing MLV-derived retroviral vector. Flow cytometry showed that the degree of GFP expression after transduction with SCF and TPO was comparable to that achieved with the full cocktail, whereas no significant GFP expression was observed after transduction with SCF, TPO, or FL alone or with the IL-6 and sIL-6R combination (Fig. 4E). When transduction efficiency was tested by colony assay, the effect of SCF and TPO was more prominent (Table 1), and the colony assay also showed that the SCF and TPO combination could maintain progenitor cells with colony-forming ability at a level comparable to that achieved with the complete cocktail.
The data indicated that the SCF and TPO combination is sufficient to induce integrin expression that is adequate for effective MLV-derived retroviral transduction in the presence of CH-296, and that FL, IL-6, and sIL-6R are not essential for transduction of MLV-derived retroviruses with CH-296.
Discussion
Introduction of the recombinant human fibronectin fragment CH-296 into the MLV-derived retroviral gene transduction of HSPCs has contributed significantly to progress in stem cell gene therapy and has enabled success in clinical trials of gene therapy for children with inherited immunodeficiencies (Ariga et al., 2001). In theory the effect of CH-296 is achieved by colocalizing the MLV-derived retrovirus and target cells of specific adhesion domains of CH-296, including integrin-binding sites (Hanenberg et al., 1996). Although the efficacy of this method for MLV-derived retroviral gene transduction of human HSPCs has been confirmed in a number of experiments in vitro (Murray et al., 1999), the actual role of the integrins expressed on HSPCs had not been analyzed. In the present study we investigated the kinetics of VLA-4 and VLA-5 expression on CD34+ cells during ex vivo culture for MLV-derived retroviral gene transduction and demonstrated the importance of integrins for efficient transduction.
As demonstrated in this study, use of either the anti-CD49d antibody or anti-CD49e antibody, both of which are capable of inhibiting integrin-mediated cell adhesion, led to a reduction in the efficiency of transduction with the MLV-derived retroviral vector (Fig. 3). Because culture conditions insufficient to enhance CD49d expression on CD34+ cells were found to result in extremely low gene transduction (Fig. 4E), abundant levels of integrins VLA-4 and VLA-5 are required for the effective transduction of MLV-derived retroviral vectors. Interestingly, CD49d expression on the surface of CD34+ cells has been shown to be significantly downregulated during treatment with MLV-derived retroviral gene transduction in the presence of CH-296, and the magnitude of the downregulation was far greater than that of CD49e expression. Because downregulation of integrins is thought to indicate adhesion-mediated activation of integrin signaling (Pellinen and Ivaska, 2006), our data indicate that VLA-4 is more effectively activated in the presence of CH-296. We also observed that the anti-CD49d antibody inhibited MLV-derived retroviral gene transduction more effectively than the anti-CD49e antibody did. Thus, our findings indicate that VLA-4 plays a more important role in CH-296-mediated MLV-derived retroviral gene transduction than VLA-5 does, although more detailed experiments are needed to be able to draw a definite conclusion.
The levels of VLA-4 and VLA-5 on CD34+ cells are quite low immediately after purifying the cells from a source such as cord blood, and they are insufficient for effective MLV-derived retroviral gene transduction. As we demonstrated in this study, the cells must be cultured with certain cytokines to yield adequate integrin levels for effective MLV-derived retroviral transduction. Therefore, pretransduction culture is needed not only to introduce cells into the cell cycle but also to induce sufficient expression of VLA-4 and VLA-5 on CD34+ cells. It is also important to be able to induce the expression of VLA-4 and VLA-5 with only a limited number of cytokines, and we determined that a combination of SCF and TPO is the minimum required for sufficient expression of integrins to achieve effective MLV-derived retroviral gene transduction.
A combination of SCF, TPO, FL, and IL-3 is commonly used for MLV-derived retroviral gene transduction, and a combination of SCF, TPO, FL, IL-6, and soluble IL-6 receptor has proven useful for stem cell gene therapy to treat ADA deficiency. Although both cocktails are able to maintain stem cells or progenitor cells and supply a sufficient number of target cells in which the defective gene has been replaced (Ariga et al., 2001; Gaspar et al., 2006), they induce significant expression of CD33, a myeloid marker antigen, in most CD34+ cells during the course of MLV-derived retroviral gene transduction (data not shown). Because our data indicate that the combination of SCF and TPO is sufficient to induce adequate integrin expression to achieve effective MLV-derived retroviral gene transduction, it is worth investigating more appropriate combinations of cytokines based on these two, to expand the population of progenitor cells for more successful replacement with functionally complemented target cells according to the lineage of the defective cells in each disease. Interestingly, Dao and Nolta have reported that engagement of the integrins VLA-4 and VLA-5 to CH-296 in combination with cytokines induces up- and downregulation of specific transcription factors and sustains the capacity of cultured CD34+ cells to undergo hematopoiesis in immunodeficient mice, suggesting that CH-296-induced stimulation via integrins affects the maintenance of their ability as HSPCs (Dao and Nolta, 2007). The use of CH-296 in MLV-derived retroviral gene transduction should therefore not only be beneficial in terms of achieving higher transduction efficiency but in maintaining HSPCs.
In conclusion, we confirmed that expression of VLA-4 and VLA-5 on HSPCs is necessary for effective CH-296-based MLV-derived retroviral gene transduction and that a combination of cytokines is important to inducing expression of integrins on HSPCs. Our results should aid in the development of a more successful protocol of MLV-derived retroviral gene transduction for stem cell gene therapy with CH-296.
Footnotes
Acknowledgments
The authors thank the Tokyo Cord Blood Bank for the distribution of cord blood for research use. This work was supported by Health and Labor Sciences Research grants (Research on Children and Families [H19-003 and H18-005], the third term comprehensive 10-year-strategy for cancer control [H19-010], Research on Human Genome Tailor Made and Research on Publicly Essential Drugs and Medical Devices [H18-005]) from the Ministry of Health, Labor, and Welfare of Japan. It was also supported by a grant from the Japan Health Sciences Foundation for Research on Publicly Essential Drugs and Medical Devices (KHA1004), and by a Child Health and Development grant from the Ministry of Health, Labor, and Welfare of Japan. The study was also supported by CREST, JST.
Author Disclosure Statement
No competing personal or financial interests exist for any of the authors in relation to this manuscript.