
Abstract
The highly aggressive cancer syndrome of female mice carrying a p53 knockout allele and a rat HER-2/neu (Neu) transgene (BALB-p53Neu) can be prevented by a cell vaccine presenting three components: Neu, interleukin (IL)-12 production, and allogeneic major histocompatibility complex (MHC) alleles (Triplex cell vaccine). Here we tested a second-generation Triplex DNA-based vaccine (Tri-DNA), consisting of the combination of three gene components (a transmembrane–extracellular domain fragment of the Neu gene, IL-12 genes, and the H-2Dq allogeneic MHC gene), carried by separate plasmids. The Tri-DNA vaccine was at least as effective as the Triplex cell vaccine for cancer immunoprevention, giving a similar delay in the onset of mammary cancer and complete protection from salivary cancer. Both vaccines induced anti-Neu antibodies of the murine IgG2a isotype at similar levels. The Tri-DNA vaccine gave more restricted immunostimulation, consisting of a fully helper T cell type 1 (Th1)-polarized response, with effective production of interferon (IFN)-γ in response to the vaccine but no spontaneous production, and no induction of anti-Neu IgG3 antibodies. On the other hand, the Triplex cell vaccine induced both Th1 and Th2 cytokines, a strong increase in spontaneous IFN-γ production, and high levels of IgG3 antibodies recognizing Neu-positive syngeneic cells. In conclusion, the Tri-DNA vaccine is as effective as Triplex cell vaccine, exploiting a more restricted immune stimulation.
Introduction
DNA vaccines could offer advantages in terms of feasibility and safety: In fact, several anticancer therapeutic DNA vaccines have already entered clinical testing, and are well tolerated (Rice et al., 2008). Electroporation greatly increases the efficacy of DNA vaccines administered by the intramuscular route (Fattori et al., 2002; Rice et al., 2008). The central role in DNA vaccination played by antigen-presenting cells through direct antigen presentation and cross-priming has been clearly demonstrated, but electroporation seems also to elicit direct presentation by transfected muscle cells (Shirota et al., 2007). It has been proposed that plasmid DNA-based vaccines have intrinsic helper T cell type 1 (Th1)-inducing adjuvant properties due to the presence of unmethylated CpG sequences (Krieg et al., 1998), which mediate production of Th1-inducing cytokines via Toll-like receptor (TLR)-9 (Hemmi et al., 2000). In addition, it was reported that double-stranded DNA, delivered by electroporation, has an adjuvant effect in itself through TANK-binding kinase-1 and interferon (IFN) regulatory factor-3-dependent innate activation of both immune and nonimmune cells to produce type I interferon and their inducible genes (Ishii et al., 2008). A cytosolic DNA sensor was found to activate these pathways (Wang et al., 2008).
Electroporated Neu DNA vaccines gave interesting results in transgenic murine models (Quaglino et al., 2004; Pupa et al., 2005; Jacob et al., 2006; Cho et al., 2008) and are being studied in clinical trials in therapeutic approaches (Wei et al., 2008). We therefore designed a second-generation Triplex DNA vaccine (Tri-DNA) based on the combination of three plasmids, each carrying one of the required components (Neu, IL-12, and an allogeneic histocompatibility gene), to be administered by intramuscular injection and electroporation. To test both the immune response elicited by Tri-DNA vaccine and its cancer-preventive efficacy in comparison with Triplex cell vaccine, we choose a model system able to outline either better or worse effects. Mice knocked out for a p53 allele and transgenic for Neu (BALB-p53Neu mice) quickly develop a cancer syndrome, consisting of salivary and mammary tumors (in females) and of rhabdomyosarcoma and salivary tumors (in males). Early administration of the Triplex cell vaccine gave high prevention of both salivary and mammary tumors (Croci et al., 2004). We therefore compared Tri-DNA and Triplex cell vaccination in BALB-p53Neu females, using short vaccination schedules (5 to 14 weeks of age), and found that the Tri-DNA vaccine is as effective as Triplex cell vaccine, and can avoid for unnecessary immune stimulation.
Materials and Methods
Mice and cell lines
Mice knocked out for the p53 oncosuppressor gene and transgenic for a mutant rat HER-2/neu oncogene driven by a mouse mammary tumor virus (MMTV) long terminal repeat (LTR) (hereafter referred to as BALB-p53Neu) were obtained as reported (Croci et al., 2004). Parental BALB/c p53 +/– mice (BALB/cJ-Trp53tm1Tyj ) were purchased from Jackson Laboratory (Bar Harbor, ME). BALB/c Neu-transgenic mice (BALBneuT) were bred in our animal facilities (Nanni et al., 2001). All of the experiments with animals fulfilled institutional guidelines and were approved by the institutional committee for animal use and care. Neu-positive TT12.E2 cells (here referred to as Neu/H-2q cells) and Neu-negative N202.1E cells (here referred to as Neuneg/H-2q cells) derived from mammary carcinomas arisen in FVB-NeuN#202 mice, transgenic for the rat neu proto-oncogene (Nanni et al., 2000). Neu-positive TUBO cells (here referred to as Neu/H-2d cells) derived from a mammary carcinoma arisen in BALBneuT mice (Rovero et al., 2000). IL-12-engineered TT12.E2 cells (here referred to as Neu/H-2q/IL12) were obtained and described previously (De Giovanni et al., 2004).
Plasmids
Plasmid pcDNA3 encoding a transmembrane–extracellular domain fragment of rat HER-2/neu (here referred to as pNeu) has been described previously (Quaglino et al., 2004). Plasmid encoding both murine IL-12 genes (pIL12) was bicistronic pIL12-IRES1neo (De Giovanni et al., 2004; Faggioli et al., 2008); it induced a high IL-12 expression level when used to obtain stable IL-12-engineered cells (De Giovanni et al., 2004). To check its ability to express IL-12 in vivo, we performed intramuscular injection of 50 μg of pIL12 followed by electroporation (see below for conditions) and collected sera at various time points thereafter. Empty vector pIRES1neo was used under the same experimental conditions. The presence of IL-12 and IFN-γ in sera was assessed through enzyme-linked immunosorbent assays (ELISAs) (R&D Systems, Minneapolis, MN).
To obtain a plasmid encoding an allogeneic MHC gene, we chose a gene of the same haplotype carried by the Triplex cell vaccine (H-2q, FVB/N strain) and cloned H-2Dq cDNA from various sources (lymphocytes and tumor cells). Briefly, cells were lysed in TRIzol (Invitrogen, Milan, Italy) and total RNA was isolated with the RNeasy system (Qiagen, Chatsworth, CA). The open reading frame of H-2Dq was amplified by RT-PCR from total RNA, using the following primers: forward, 5′-CGATGGCTCCGCGCACGCTGCTC-3′ and reverse, 5′-CAGTCCAGGCAGCTGTCTTCACGC-3′. H-2Dq cDNA was cloned in the PCR2.1 vector, using the TOPO-TA cloning system (Invitrogen), sequenced, and then subcloned into the pcDNA3 expression plasmid (Invitrogen): this plasmid is hereafter referred to as pDq. Sequencing of the various H-2Dq cDNA clones obtained showed almost complete homology with a reported Dq sequence (Pullen et al., 1992), with only two mismatches leading to amino acid changes (CAG/AGG at codon 69 leading to a Q-to-R amino acid change, and CGC/GCC at codon 104 leading to an A-to-R change). Because of the consistency of our sequences in different clones, mismatches can be interpreted either as mistakes in the previously published sequence (Pullen et al., 1992) or as substrain variations.
Transfection
To verify the ability of pDq to transfer H-2Dq expression, a variant of B16 melanoma (B78H1) was chosen, because it is a good transfection recipient and does not express MHC molecules (Graf et al., 1984; De Giovanni et al., 1991). B78H1 cells were seeded at the concentration of 0.25 × 106 cells per well in 6-well plates (Falcon; BD Biosciences, Bedford, MA) and left to attach for 24 hr. pDq (2 μg/well) was then transfected with Lipofectamine 2000 (Invitrogen) at a 1:2.5 ratio (DNA [μg]:Lipofectamine [μl]) according to the manufacturer's protocol. Two days after transfection H-2Dq expression was evaluated on harvested cells, through indirect immunofluorescence and flow cytometry, using the monoclonal antibody 28-14-8S (Ozato et al., 1982) (Cedarlane Laboratories, Hornby, ON, Canada).
Complement-dependent cytotoxicity
BALB/c mice (H-2d ) at 5 and 7 weeks of age were vaccinated by intramuscular injection of pDq plasmid into the tibial muscles (25 μg in 20 μl, each muscle) followed by electroporation (according to the same protocol reported below for DNA vaccination). One week after the second injection, pDq-vaccinated mice were bled and serum was stored at −80°C. Aliquots of 0.5 × 106 lymphocytes, collected from BALB/c (H-2d) and FVB-NeuN#202 (H-2q) mice, were resuspended in RPMI + 5% fetal bovine serum (FBS) containing serum from pDq-vaccinated and untreated BALB/c mice (serum diluted 1:10 in a final volume of 0.5 ml) and incubated for 30 min in ice. A baseline control in RPMI + 5% FBS medium and a positive control, consisting of 28-14-8s monoclonal antibody (recognizing both H-2q and H-2d epitopes) at 1:10 dilution, were run in parallel. Lymphocytes were then washed once with phosphate-buffered saline (PBS) and incubated with a 1:10 dilution of rabbit complement (Cedarlane Laboratories) for 30 min in ice. Lymphocytes were then washed again and resuspended in RPMI + 5% FBS, and cell number and viability were determined by erythrosin dye exclusion test. The percentage of cytotoxicity was calculated as follows: 100 − (cell yield in complement-containing RPMI + 5% FBS/cell yield with serum) × 100.
DNA and cell vaccinations
Plasmid large-scale preparations were set up and purified with Endofree Qiagen Plasmid-Giga kits (Qiagen). DNA was precipitated, dissolved in water at a concentration of 3–5 mg/ml, and stored in monodose aliquots at −20°C for use in immunization protocols. For the Tri-DNA vaccine, 50-μg amounts of each plasmid were mixed together and diluted to a final volume of 40 μl per mouse in 0.9% NaCl and polyglutamate (6 mg/ml) (final concentrations). Groups treated with a combination of two plasmids or with pNeu alone received 50 μg of each plasmid. Anesthetized mice received the injection of DNA vaccine into the tibial muscles (20 μl in each muscle) through a 28-gauge needle syringe and immediately thereafter were subjected to electroporation, consisting of two square-wave 25-msec, 375-V/cm pulses generated with a T830 electroporator (BTX, San Diego, CA). Mice received four vaccinations (at 5, 7, 12, and 14 weeks of age). The control group received mock vaccinations as described previously, but lacking plasmids. Mice were monitored weekly for mammary or salivary tumor onset.
Triplex cell vaccine consisted of Neu-positive IL-12-engineered allogeneic cells (Neu/H-2q/IL12). Cells and vaccine schedule were previously reported (Croci et al., 2004; De Giovanni et al., 2004), with the exception that only the first three vaccination cycles were performed (from 5 to 14 weeks of age). Mice were then monitored as described previously.
Cytokine production and cell-mediated cytotoxicity
Mice were killed 1 week after the first cycle of vaccination and spleens were collected. Total spleen mononuclear cells, or subpopulations (CD4+ or CD8+) purified by magnetic sorting (Miltenyi Biotec, Bergisch Gladbach, Germany), were cultured for 6 days alone or in the presence of proliferation-blocked restimulator cells (at a 10:1 lymphocyte:tumor cell ratio) in RPMI 1640 supplemented with 10% fetal bovine serum and with recombinant IL-2 (20 units/ml), as described (Nanni et al., 2001). Culture supernatants were collected and concentrations of IL-4, IL-5, IL-10, IL-12(p70), granulocyte-macrophage colony-stimulating factor (GM-CSF), IFN-γ, and tumor necrosis factor (TNF)-α were simultaneously determined with the Bio-Plex 200 suspension array system (Bio-Rad, Milan, Italy). A commercially available premixed multiplex cytokine set was used (mouse cytokine Th1/Th2 panel) in accordance with the manufacturer's instructions and at a high-sensitivity setting. Briefly, 50-μl volumes of supernatants and serial dilutions of the cytokine standards were incubated at room temperature for 30 min with a mix of different fluorescently dyed beads conjugated with monoclonal antibodies specific for the various cytokines. Afterward samples were incubated for 30 min with 25 μl of detection antibody and then with 50 μl of streptavidin–phycoerythrin (PE) for 10 min. Each step was followed by three washings. Finally, samples were resuspended in 125 μl of assay buffer and analyzed with the Bio-Plex reader. A 50-μl volume was sampled from each well and the PE-fluorescent signal of a minimum of 100 beads per cytokine type was evaluated. Bead doublets were excluded from analysis. Data were elaborated with Bio-Plex Manager Software 5.0. Extrapolating PE-fluorescent signals to a standard curve allowed quantitation of each cytokine in the samples. The experimental ranges of detection of the various cytokines were as follows: IL-4, 2100–0.2 pg/ml; IL–5, 500–0.1 pg/ml; IL-10, 1100–1 pg/ml; IL-12, 1100–1 pg/ml; GM-CSF, 500–2 pg/ml; IFN-γ, 1400–0.3 pg/ml; TNF-α, 3700–3 pg/ml. Production of murine IL-4 and IFN-γ was also determined in parallel by ELISAs (Endogen, Woburn, MA) with similar results. For the cytotoxicity assay spleen mononuclear cells of mice that had completed the vaccination cycles (15 weeks old) were restimulated by coculture at a 50:1 ratio with proliferation-blocked Neu/H-2d cells for 6 days in RPMI 1640 supplemented with 10% fetal bovine serum and recombinant IL-2 (20 units/ml). The ability of restimulated lymphoblasts to lyse Neu/H-2d or Neu/H-2q tumor cells was evaluated in a standard 51Cr release assay as described (De Giovanni et al., 2004).
Antibody response
Mice were routinely bled via the tail vein and serum samples were stored frozen at −80°C. Production of antibodies was then studied both by ELISA and by indirect immunofluorescence of syngeneic and allogeneic Neu-positive cells. An ELISA to detect antibodies recognizing rat HER-2 was performed in Maxisorp NUNC 96-well microplates as reported (Cipriani et al., 2008). Plates coated with rat Neu were incubated overnight with serum at 1:300 dilution, washed, incubated with alkaline phosphatase-conjugated goat anti-mouse IgG antibody (Sigma-Aldrich, St. Louis, MO) for 30 min at room temperature, washed again, and developed with p-nitrophenyl phosphate (Sigma-Aldrich). A standard curve with anti-c-ErbB2/Neu mouse monoclonal antibody (clone 7.16.4; EMD Biosciences, San Diego, CA) was run in parallel (0.10–250 ng/ml). For immunofluorescence studies, Neu/H-2d and Neu/H-2q cells were incubated with sera at a 1:65 dilution for 30 min in ice, and then washed and incubated with an Alexa Fluor 488 F(ab′)2 fragment of goat anti-mouse IgG (H-L) chains antibody (Invitrogen Molecular Probes, Eugene, OR) to evaluate total immunoglobulin binding. For isotype subclass analysis, the following secondary fluorescein isothiocyanate (FITC)-conjugated monoclonal antibodies were used (BD Biosciences): anti-mouse IgG1 clone A85-1, anti-mouse IgG2a clone R19-15, anti-mouse IgG2b clone R12-3, and anti-mouse IgG3 clone R40-82. Cytofluorometric analysis was performed with a FACScan flow cytometer (BD Biosciences). Rat HER2/neu expression of target cells was evaluated in each set of experiments with anti-c-ErbB2/Neu mouse monoclonal antibody. Data from each sample were normalized over target cell expression.
Histology and immunohistochemistry
For histologic evaluation, tissue samples were fixed in 10% neutral-buffered formalin, embedded in paraffin, sectioned at 4 μm, and stained with hematoxylin and eosin (H&E). For immunohistochemistry, paraffin-embedded sections were immunostained with anti-c-erbB-2 (Dako, Milan, Italy). After washing, sections were overlaid with biotinylated anti-rabbit immunoglobulin (Jackson ImmunoResearch Europe, Suffolk, UK ) for 30 min. Unbound immunoglobulin was removed by washing, and slides were incubated with ready-to-use peroxidase streptavidin (Lab Vision, Bio-Optica Milan, Milan, Italy) and diaminobenzidine (Dako).
Statistical analysis
The Mantel–Haenzel test was used to compare tumor latency Kaplan–Meier curves; the Student t test or Wilcoxon nonparametric test was used for other comparisons.
Results
Plasmid validation
We checked the ability of the plasmid encoding murine IL-12 (pIL12) to give rise to cytokine production in vivo. Mice received an intramuscular injection of 50 μg of pIL12 and electroporation under the same conditions chosen for vaccination, and then were bled at several time points. A rise in serum IL-12 (and secondary induction of serum IFN-γ) followed pIL12 injection and remained at detectable levels for some days (Fig. 1A). Similar data have been reported in the literature (Faggioli et al., 2008). Serum IL-12 or IFN-γ levels were undetectable in untreated mice as well as in mice treated with empty pIRESneo, pNeu, and pDq vectors at up to 150 μg/mouse administered via intramuscular injection plus electroporation (data not shown). Transfection of a MHC-negative cell line with plasmid encoding H-2Dq (pDq) gave rise to cells with surface expression of H-2Dq (Fig. 1B). Intramuscular injection of pDq followed by electroporation in H-2d mice determined complement-dependent cytotoxic activity toward H-2q, but not H-2d, lymphocytes (Fig. 1C). Therefore pDq plasmid is able to transfer H-2Dq expression and to elicit a specific antibody response. Plasmid encoding a transmembrane–extracellular domain fragment of rat HER-2/neu (pNeu), already used as a single DNA vaccine in our work and in other studies reported in the literature (Rovero et al., 2000; Quaglino et al., 2004; Pannellini et al., 2006; Cho et al., 2008), induced effective anti-Neu antibody responses in Neu-transgenic mice.

Validation of pIL12 and pDq vectors. (
Immunoprevention of cancer syndrome through DNA vaccines
Tri-DNA vaccine consisted of the combination of the three plasmid vectors pNeu, pIL12, and pDq (also referred to as pNeu+pIL12+pDq). BALB-p53Neu female mice received vaccinations with Tri-DNA at 5, 7, 12, and 14 weeks of age, in comparison with Triplex cell vaccine administered in the same time window (Fig. 2). Groups of mice were also vaccinated with DNA vaccines lacking allogeneic MHC gene or both adjuvants (referred to as pNeu+pIL12 and pNeu, respectively). Vaccinated mice were observed weekly, with recording of the occurrence of the two types of cancer typical of such a model, that is, salivary and mammary cancer. BALB-p53Neu females, either untreated (Croci et al., 2004) or subjected to mock vaccination and electroporation (Fig. 2, dashed line), showed a fast and almost simultaneous occurrence of salivary and mammary cancers (with latencies of 13–15 weeks for the former and slightly longer for the latter). All the vaccines studied, both DNA based and cellular, induced highly significant delays in the onset of both types of tumors. Vaccinated mice reached a median tumor-free survival time of 45–55 weeks for mammary cancer. A 90–100% prevention of salivary tumors was obtained with Triplex cell and Tri-DNA vaccines. Overall survival for the different groups and the relative frequency of the two types of tumors at sacrifice are shown in Fig. 3. Almost all untreated mice developed salivary tumors, rapidly arising and leading to death (see also Fig. 4A–D), and in about half of the cases combined with mammary tumors (Fig. 3, bottom). In vaccinated mice the situation was almost reversed: Salivary tumors rarely occurred alone even in mice treated with the simplest DNA vaccines. In particular, with the Tri-DNA vaccine salivary cancer was fully prevented and mouse survival was determined by mammary carcinoma onset. Similar results were obtained with the Triplex cell vaccine. Tri-DNA-vaccinated mice in fact showed morphologically normal salivary glands with well-defined ducts and acini that were enclosed in delicate stromal tissue (Fig. 4E). On-going inflammatory aspects or inflammation effects (e.g., scars) were not detected. Neu expression was not found in salivary glands from Tri-DNA-vaccinated mice (Fig. 4F), although it was observed in hyperplastic ducts before vaccination (Fig. 4B). On the other hand, Tri-DNA-vaccinated mice showed several Neu-positive mammary cancer nodules with inflammatory cells in their stroma (Fig. 4G and H).
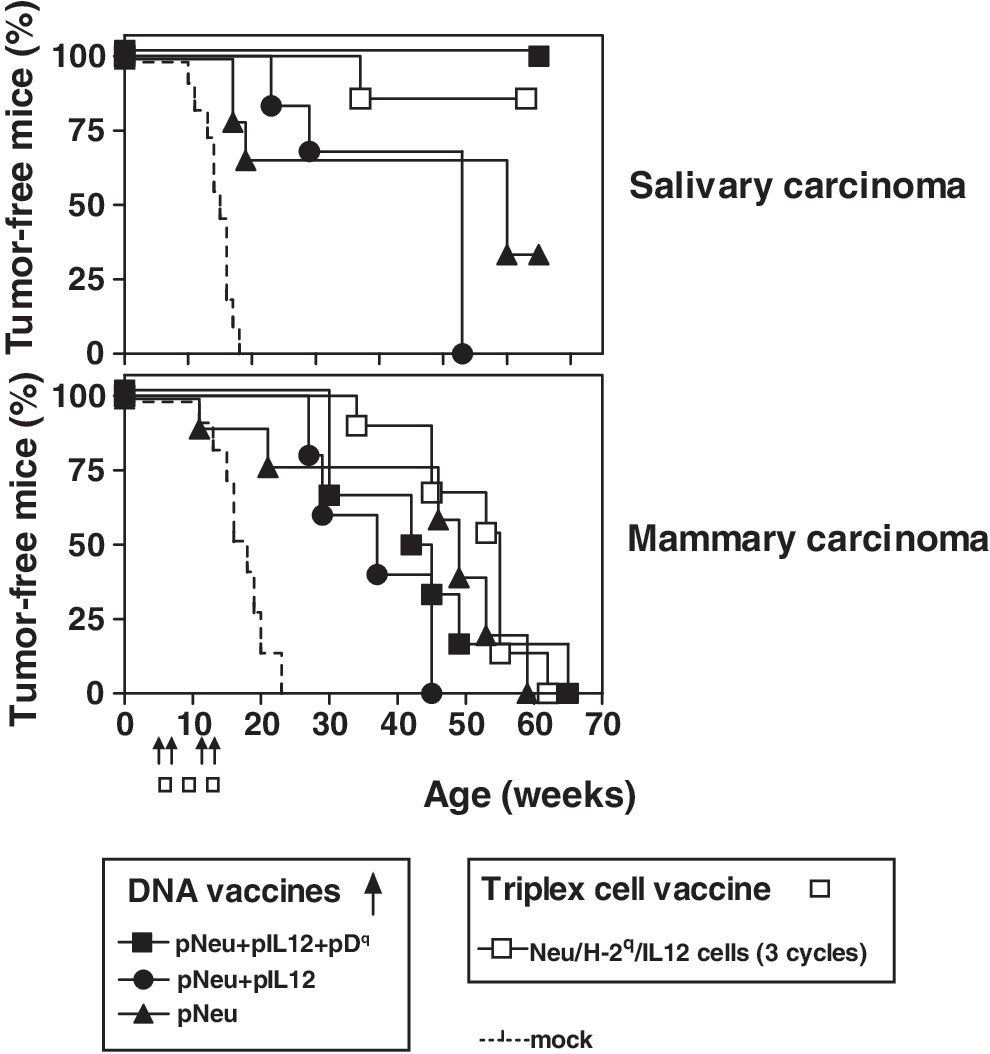
Cancer-immunopreventive activity of DNA vaccines and Triplex cell vaccine in BALB-p53Neu female mice (6–11 mice per group). Vaccination cycles are shown with arrows (DNA vaccines) or boxes (Triplex cell vaccine). Dashed line: Mock-electroporated control mice. Significance of differences with Mantel–Haenzel test: any vaccine versus mock, p < 0.05 at least (both salivary and mammary carcinoma); pNeu+pIL12+pDq versus pNeu+pIL12 or pNeu, p = 0.074 or p = 0.096, respectively (salivary carcinoma only).

Effect of DNA vaccines and Triplex cell vaccine on overall survival (top) and tumor frequency at death (bottom) of BALB-p53Neu female mice (6–11 mice per group).
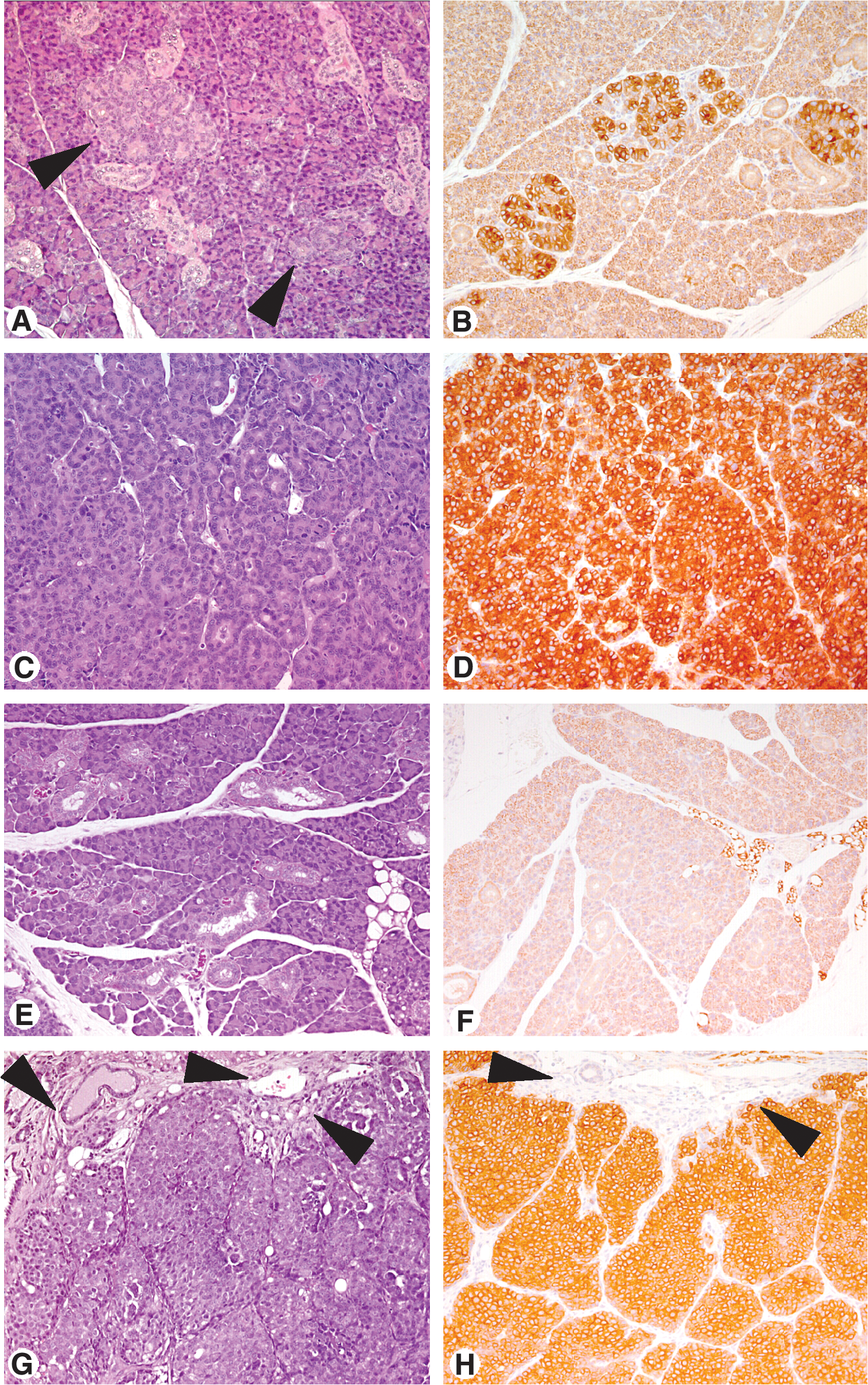
Histology (
Because key mechanisms of the immunopreventive effect in Neu transgenic mice rely on IFN-γ production and antibodies (Nanni et al., 2004), we focused on these mechanisms to compare multi-DNA and cell vaccines.
Cytokine production and cell-mediated cytotoxicity
To study how the various vaccines drive the cellular immune responses, we studied the cytokine production profile of vaccinated mouse spleen cells cultured alone (spontaneous production) or on in vitro restimulation (Fig. 5A). The Triplex cell vaccine induced high production of IFN-γ, already evident without in vitro restimulation, but at the same time induction of IL-4 together with other Th2 cytokines such as IL-5 and IL-10 was observed. Spleen cells from the Tri-DNA vaccine group showed significant IFN-γ production only on restimulation with cells presenting allogeneic MHC alone or combined with Neu expression. Spleen cells from the other DNA vaccine groups (pNeu+pIL12 and pNeu alone) as well as those from mock-electroporated control mice (data not shown) showed barely detectable spontaneous cytokine production levels; in vitro stimulation did not increase IFN-γ or IL-4.
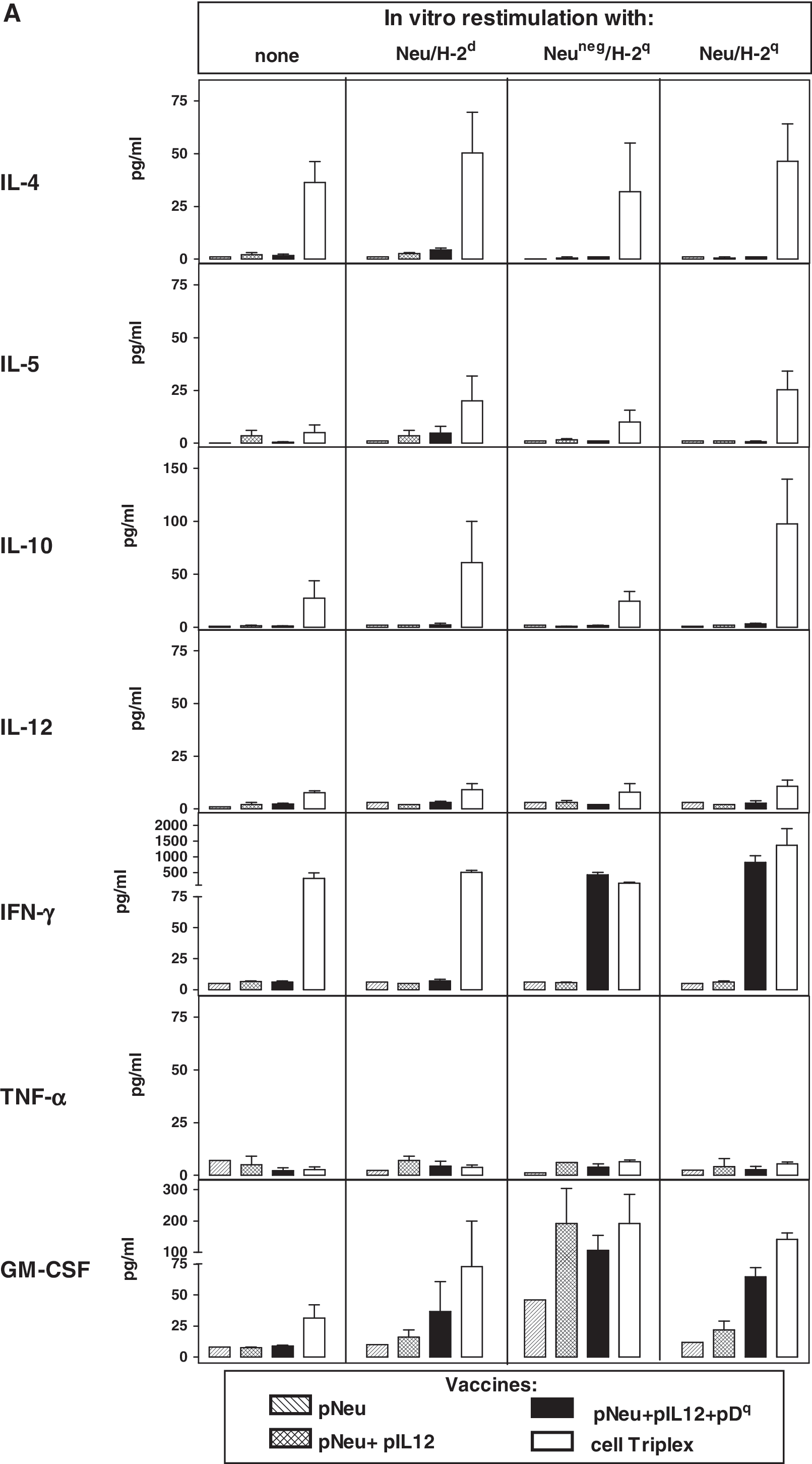
Cytokine production by spleen cells collected after the first vaccination cycle and cultured for 6 days alone or in the presence of the indicated restimulator cells. (

The production of IFN-γ and IL-4 by CD4+ and CD8+ populations was also studied (Fig. 5B). Triplex cell vaccine elicited both IFN-γ- and IL-4-producing CD4+ cells; in vitro restimulation with Neu-negative allogeneic cells, but not with Neu-positive allogeneic cells, led to a significant decrease in IL-4 production by CD4+ cells. CD8+ splenocytes from the Triplex cell group showed low spontaneous IFN-γ production, which increased on restimulation, reaching high levels only in the presence of allogeneic Neu-positive stimulator cells. In the DNA vaccine groups CD4+ cells showed low IL-4 production even after in vitro restimulation; IFN-γ production was found to be significant only in cultures restimulated with allogeneic cells, with the highest values in CD8+ cultures.
Neither the Triplex cell nor Tri-DNA vaccine gave rise to cytotoxic effectors against Neu/H-2d cell targets (data not shown).
On the whole, the Triplex cell vaccine gave rise to powerful Th1 plus Th2 immune stimulation (also comprising cytokines with immunosuppressive activity, such as IL-10), both of which were increased by culture with Neu-positive syngeneic cells. On the other hand, the Tri-DNA vaccine induced a highly restricted Th1 response, stimulated mainly by allorecognition.
Humoral response
Antibodies recognizing the Neu antigen were specifically detected by ELISA (Fig. 6A). All the mice in the Tri-DNA vaccine group (solid squares) already showed detectable levels of anti-Neu antibodies after the first vaccination cycle, whereas in the groups receiving DNA vaccines lacking one or both adjuvants as well as in the Triplex cell vaccine group some late-responder mice were also found. After the second vaccination cycle all the DNA vaccines studied produced similar ranges of anti-Neu antibody levels. At the end of the vaccination cycles the highest antibody levels were obtained with the Triplex cell vaccine (Fig. 6A, open squares).
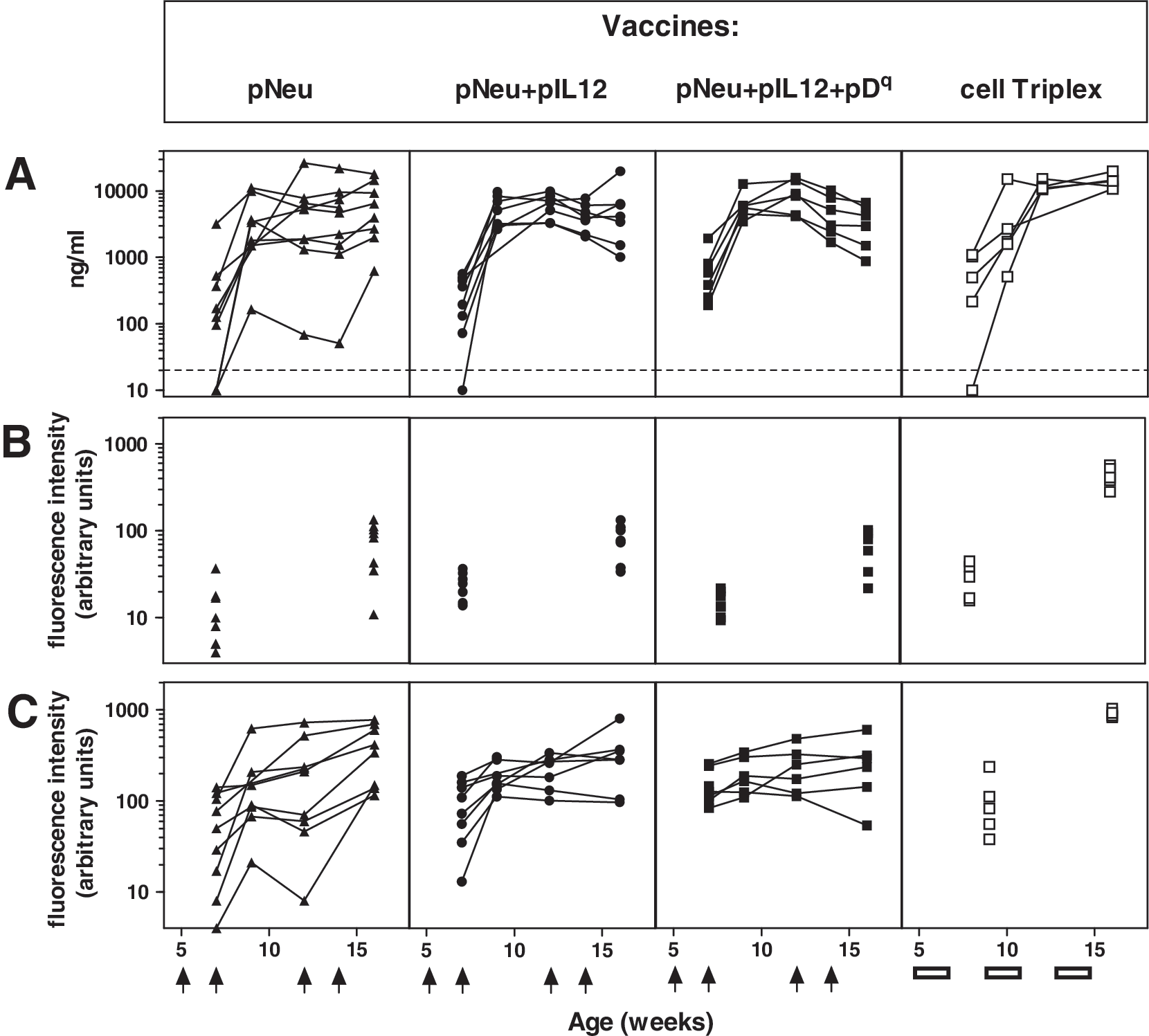
Humoral response of BALB-p53Neu female mice vaccinated with the indicated DNA vaccines compared with Triplex cell vaccine. Vaccination cycles are shown with arrows (DNA vaccines) or boxes (Triplex cell vaccine). Data from five to nine individual mice are shown in each panel. (
The use of a cell vaccine such as the allogeneic Triplex vaccine could induce antibodies recognizing other cell surface moieties, some of which could be downstream to Neu. Transduction of muscle cells could elicit antibodies recognizing molecules induced by the Neu pathway: in fact, skeletal muscle cells naturally show some level of HER-2 expression and can respond to its pathway (LeBrasseur et al., 2005). To study the antibody response directed against Neu-expressing cells, we then tested the ability of sera from vaccinated mice to bind syngeneic or allogeneic neu-positive cells (Neu/H-2d and Neu/H-2q cells, respectively; Fig. 6B and C). Kinetics of induction of humoral reactivity were similar to those found with ELISAs. Mock-electroporated control mice did not develop any humoral response, both when tested in ELISA and as binding to Neu-positive cells (data not shown). The reactivity against Neu/H-2q cells observed with sera from groups including allogeneic MHC in vaccine formulation obviously comprises anti-histocompatibility antibodies; however, a minor contribution to binding is attributable to histocompatibility because such sera tested against a Neu-negative H-2Dq-expressing target cell had low binding activity (data not shown). In fact, DNA vaccines lacking an allogeneic MHC gene (pNeu+pIL12 and pNeu alone) could induce antibodies almost as effectively as the DNA vaccine including the H-2Dq gene.
On the whole, the various DNA vaccines did not reach remarkably different antibody levels, but the presence of adjuvants in the Tri-DNA vaccine seems to confer a faster and more homogeneous response. Sera from the Triplex cell vaccine group reached significantly higher levels of binding both to syngeneic and to allogeneic Neu-positive cells.
To further investigate whether the various vaccines could also lead to qualitatively different humoral responses, we studied the isotype profile of the antibodies binding syngeneic Neu-positive cells (Fig. 7). All the vaccines studied induced comparable levels of IgG2a antibodies, but only the Triplex cell vaccine caused a high level of IgG3 antibodies, together with a slight increase in IgG1 antibodies.

Analysis of antibody isotypes induced by DNA and Triplex cell vaccination. Normalized fluorescence intensity against Neu/H-2d cells by sera collected from 16-week-old mice is shown. Shown are the means and standard error of five to nine mice per group.
Discussion
The Tri-DNA vaccine was designed as a second-generation Triplex vaccine, because it combines three plasmids each carrying one of the three components conferring the high immunopreventive effectiveness of the Triplex cell vaccine: the Neu target antigen and, as adjuvants, IL-12 and allogeneic MHC molecules. The Tri-DNA vaccine was as effective as the Triplex cell vaccine, but induced more restricted immunostimulation. In particular, the Tri-DNA vaccine gave a fully Th1-polarized response, with effective production of IFN-γ in response to the vaccine but no spontaneous production, and no induction of IgG3 antibodies binding syngeneic Neu-positive cells. On the other hand, the Triplex cell vaccine induced both Th1 and Th2 cytokines, high spontaneous IFN-γ production, and high levels of IgG3 antibodies against syngeneic cells.
The Tri-DNA vaccine has controlled antigenicity, in comparison with the Triplex cell vaccine. In fact, a cellular vaccine in principle could carry a portfolio of immune stimuli, other than desired, whereas our Tri-DNA vaccine carries only the three genes chosen. In particular, an allogeneic cell vaccine carries multiple different MHC genes. In the Tri-DNA vaccine a single MHC gene was used as adjuvant instead of a complete haplotype. The MHC gene chosen, H-2Dq , has about 88% homology with the corresponding MHC gene of the host d haplotype; nevertheless, it gave effective stimulation as well, as determined on the basis of the high IFN-γ response by allo-restimulated spleen cells. Such IFN-γ induction could take place even in vivo after each vaccination cycle, contributing to the effectiveness of the vaccine.
DNA vaccines are generally well tolerated (Rice et al., 2008). However, potential side effects must be taken into consideration, such as the risk for a stable integration potentially leading to insertional mutagenesis on the one hand (Rice et al., 2008) and prolonged antigen expression leading to immune anergy on the other (Radcliffe et al., 2007). Our choice to combine a HER-2 DNA vaccine with an allogeneic MHC gene vaccine could overcome both concerns, because allorecognition, besides its Th1-driving effect (Wallgren et al., 2005), should lead to rapid elimination of transfected cells (Gronevik et al., 2005). Plasmids carrying the IL-12 genes are currently being tested to correct Th2-mediated pathologies such as allergic asthma (Malheiro et al., 2008).
Monoantigenic anti-Neu-electroporated DNA vaccines have been reported to confer protection against carcinogenesis driven by Neu alone (Quaglino et al., 2004; Cipriani et al., 2008). Carcinogenesis induced by the combination of Neu and p53 knockout is much more aggressive and difficult to prevent (Croci et al., 2004; Pannellini et al., 2006). In this model Tri-DNA vaccine was more effective than the monoantigenic Neu vaccine in keeping all mice free of salivary tumors. The presence of the two adjuvants (IL-12 and allogeneic MHC gene) did not modify the long-term level of antibodies induced, but gave a faster and homogeneous rise of antibodies after only one vaccination cycle. In fact, the response to DNA vaccines shows heterogeneity among individuals (Jacob et al., 2006; Cipriani et al., 2008) that is generally circumvented by repeated boosting. Because immunoprevention in Neu-transgenic models was reported to be related both to the individual antibody level (Cipriani et al., 2008) and to the precociousness of the response induced (Croci et al., 2004; Pannellini et al., 2006), it is likely that the more homogeneous response induced by the Tri-DNA vaccine combined with the high IFN-γ boost determined by allorecognition could make the difference.
In contrast to salivary carcinogenesis, however, no superior effect of the Tri-DNA vaccine was observed for mammary carcinogenesis. We hypothesize that a more defined temporal window for salivary carcinogenesis could take place, so our temporally defined schedule of vaccination (ending at 14 weeks of age) could be sufficient to completely prevent salivary carcinogenesis, whereas a longer or even life-long schedule may be required to halt carcinogenesis in the continuously evolving mammary gland.
The Tri-DNA vaccine induced a Th1-biased response and led to clarification of the antibody isotypes involved in immunoprevention. We previously demonstrated that murine IgG2a was required and that IgG1 is dispensable (Nanni et al., 2001; De Giovanni et al., 2004), but no data on the role of murine IgG3 have been reported so far in the literature. Here we found that IgG3 is not necessary for immunoprevention, because DNA vaccines, which did not induce IgG3 at all, were at least as effective as the Triplex cell vaccine, which induced high levels of IgG3. Murine IgG3 is the predominant murine IgG subclass elicited by bacterial polysaccharide antigens, and is likely to correspond to the human IgG2 isotype (Yoo et al., 2003). Murine IgG3 has mechanisms of action different from those of the other isotypes: it does not bind the Fc receptor, but can activate the complement system through Fc–Fc interactions (Getahun and Heyman, 2006; Nimmerjahn and Ravetch, 2008). Some data on the involvement of murine IgG3 in autoimmunity have also been reported (Baudino et al., 2006). Autoimmunity could be a serious side effect of cancer immunoprevention (Jacob et al., 2006; Wei et al., 2008), because cancer antigens, HER-2 included, can be expressed in normal tissues. We did not observe any sign of autoimmunity in mice of our vaccinated series (both with cell and DNA vaccines), suggesting that the high level of IgG3 antibodies elicited by the Triplex cell vaccine was not required for tumor prevention, but at the same time was not detrimental to mouse health.
In conclusion, DNA vaccines were as effective as the Triplex cell vaccine, exploiting a more restricted mode of immune stimulation. The combination of multiple plasmids and the high efficacy obtained through electroporation makes the system handy and flexible. Our results suggest that this novel strategy may be effective in the prevention of HER-2-expressing tumors in humans at high risk, but also suggest its potential efficacy in the therapeutic setting. In fact, DNA-based combined approaches are being studied for therapeutic approaches (Martel-Renoir et al., 2003; Feng et al., 2005). Multi-DNA vaccines could in principle be more suitable to tailor-cut treatment, allowing combinations of the right panel of tumor antigen, adjuvant cytokine, and adjuvant allogeneic MHC gene for each tumor type or even each individual.
Footnotes
Acknowledgments
This work was supported by grants from the following: the Italian Association for Cancer Research (Milan, Italy); the Italian Ministry for Education, University, and Research PRIN projects; the University of Bologna (Bologna, Italy); the AlmaMedicina Foundation (Bologna, Italy); and the Pallotti Funds, Department of Experimental Pathology, University of Bologna. A. Palladini is the recipient of a fellowship from the Italian Foundation for Cancer Research (Milan, Italy). S. Croci is the recipient of a research fellowship (Assegno di Ricerca) from the University of Bologna. A. Antognoli is the recipient of a Ph.D. fellowship from the University of Bologna. A. Murgo is the recipient of a research fellowship from the W. Vanini–S. Cavagnino Foundation, Interdepartmental Centre for Cancer Research, University of Bologna.
Author Disclosure Statement
For all authors, no competing financial interests exist.