
Abstract
Preclinical studies showed that the allogeneic tumor cell line RCC-26 displayed natural immunogenic potential that was enhanced through expression of CD80 costimulatory molecules and secretion of interleukin-2. Here we report the study of RCC-26/CD80/IL-2 cells in a phase 1 vaccine trial of renal cell carcinoma patients with metastatic disease (mRCC). Fifteen patients of the HLA-A*0201 allotype, with at least one metastatic lesion, were included. Irradiated vaccine cells were applied in increasing doses of 2.5, 10, and 40 × 106 cells over 22 weeks. Primary study parameters included safety and toxicity. Sequential blood samples were analyzed by interferon-γ enzyme-linked immunospot assays to detect tumor antigen-associated (TAA) effector cells. The vaccine was well tolerated and the designated vaccination course was completed in 9 of 15 patients. Neither vaccine-induced autoimmunity nor systemic side effects were observed. Delayed-type hypersensitivity skin reactions were detected in 11 of 12 evaluated patients and were particularly strong in patients with prolonged survival. In parallel, vaccine-induced immune responses against vaccine or overexpressed TAA were detected in 9 of 12 evaluated patients. No tumor regressions occurred according to RECIST (Response Evaluation Criteria in Solid Tumors) criteria; however, median time to progression was 5.3 months and median survival was 15.6 months, indicating substantial disease stabilization. We conclude that vaccine use was safe and feasible in mRCC. Clinical benefits were limited in these patients with advanced disease; however, immune monitoring revealed vaccine-induced responses against multiple TAAs in the majority of study participants. These results suggest that this vaccine could be useful in combination therapies and/or minimal residual disease.
Introduction
Among patients with cancer, those with RCC show some of the most favorable responses to immunotherapy (Kim et al., 2003; Motzer, 2003). Cancer vaccines represent one treatment strategy to exploit this immunological potential because there is evidence that immunization against tumors can reduce or even eliminate some tumors and induce long-lasting T cell memory responses with a capacity to control tumor relapse. Nevertheless, vaccine treatment alone has shown clinical benefit in only a small number of patients with mRCC (Simons et al., 1997; Frankenberger et al., 2005b; Schendel, 2007). Thus, future treatment strategies that combine targeted molecular therapies with low-toxicity vaccination might enhance the development of effective immune responses and improve the long-term perspectives of patients with advanced disease.
Various vaccine strategies have been developed to induce antitumor T cell responses (Pardoll, 1998), several of which have been applied in patients with mRCC (Frankenberger et al., 2005b; Schendel, 2007). One approach is to use autologous gene-modified tumor cells as vaccines, introducing immunologically relevant genes to enhance tumor cell immunogenicity (Mach and Dranoff, 2000). The first phase 1 trial in RCC implementing this strategy, through introduction of the gene encoding granulocyte-macrophage colony-stimulating factor (GM-CSF) into autologous tumor cells, demonstrated clinical benefit in several patients (Simons et al., 1997) and induction of specific T cell immunity (X. Zhou et al., 2005). More recently, an autologous gene-modified tumor cell vaccine that expressed the costimulatory molecule CD80 was tested in patients with mRCC in combination with systemic interleukin (IL)-2 (Antonia et al., 2002; Fishman et al., 2008). No severe toxicity was associated with these vaccines. However, strong limitations in feasibility and high costs are incurred with the production of individualized gene-modified tumor cell vaccines. Generic vaccines that could be applied to many patients would reduce development time and costs and enable treatment of more patients.
We developed an allogeneic gene-modified RCC vaccine based on a well-characterized tumor cell line that showed improved immunogenicity through expression of CD80 and IL-2 (Frankenberger et al., 2005a). The original cell line (RCC-26) (Schendel et al., 1993) was derived from an HLA-A*0201-positive patient with stage I disease (pT1pN0M0G2) in whom only a single brain metastasis appeared after nephrectomy, following a disease-free interval of 9 years. The RCC-26/CD80/IL-2 vaccine was tested in a clinical phase 1 trial to evaluate safety and feasibility with stage IV mRCC patients matched for the HLA-A*0201 allotype. Furthermore, the ability of vaccination to induce specific T cell reactivity was analyzed with enzyme-linked immunospot (ELISPOT) assays to detect T cell responses to vaccine cell lysates as well as to surrogate peptides that were derived from tumor-associated antigens (TAAs) that are overexpressed in mRCC lesions.
Materials and Methods
Patients
Between September 2003 and August 2006, 15 patients with histologically proven clear-cell RCC and at least one evaluable metastasis were enrolled in the clinical study. This study is registered with the German Somatic Gene Transfer Clinical Trial Database (DeReG [Deutsches Register für somatische Gentransferstudien], reference number 47). Potential study patients were screened for the HLA-A*0201 allotype first by serological typing using flow cytometry, followed by genotyping with an HLA-A*02-specific polymerase chain reaction (PCR), as described previously (Gatz et al., 2000). The PCR product encompassing exons 2 and 3 was subsequently sequenced. Only patients with an HLA-A*0201 subtype were finally included in the clinical study. Further inclusion criteria were good performance status (Eastern Cooperative Oncology Group [ECOG] score, 0 or 1), no surgery during the previous 4 weeks, no systemic antitumoral therapy during the previous 12 weeks, no evidence of autoimmune disease, and no use of systemic immune-modulatory therapies (e.g., corticosteroids). Initial staging on the basis of computer tomography (CT) and bone scan was performed 1 week before the first vaccination. All patients gave written informed consent before entering the study. The trial protocol was approved by the local ethics committee and the good manufacturing practice (GMP)-certified vaccine production was approved by the responsible national, state, and local authorities.
Vaccination and study schedule
The genetically modified RCC-26/CD80/IL-2 vaccine was generated as described previously (Frankenberger et al., 2005a). The cells were expanded under GMP conditions and stored in the gas phase of liquid nitrogen in ampoules with 2.5 × 106 or 10 × 106 cells. For safety reasons, all cryopreserved vaccine aliquots were irradiated (120 Gy) in one batch under controlled conditions. The vaccine cells were thawed immediately before patient application and given by intradermal injection into the inguinal region. Graded doses of cells were applied up to 10 times over a 22-week period. The vaccination schedule was as follows: 2.5 × 106 cells at weeks 1, 2, 4, and 6; 10 × 106 cells at weeks 8, 10, 12, and 14; and 40 × 106 cells at weeks 18 and 22. Clinical examination and routine blood checks were performed at every visit. Patients were assessed for signs of autoimmune disease by measurement of thyroid hormones and pancreatic enzymes, screening for anti-nuclear antibodies, as well as monitoring of rheumatoid factor and complement factors. Blood samples for immune monitoring were taken at weeks 1, 6, 14, 22, and 36. Restaging on the basis of CT was performed at weeks 8, 18, 24, and 36. Patients were withdrawn from the study on evidence of tumor progression according to the Response Evaluation Criteria in Solid Tumors (RECIST) (Therasse et al., 2000). In four patients, the lowest vaccine dose (2.5 × 106 cells) was omitted and vaccination was initiated with a dose of 10 × 106 cells.
Delayed-type hypersensitivity testing
We assessed the patients for delayed-type hypersensitivity (DTH) reactions to challenge with irradiated vaccine cells after 4, 8, and 10 vaccinations at weeks 6, 14, and 22, respectively. Skin tests were performed by intradermal injection of 2.5 × 106 cells at a site distant from the vaccination sites in the inguinal region. A DTH reaction evaluated 48 hr postchallenge was judged as positive if induration or erythema of at least 10 mm was observed.
Microarray analysis of TAA expression in tumors and RCC-26 cells
Tissue samples from 32 patients with clear-cell RCC metastases were collected, snap-frozen, and stored in liquid nitrogen after written informed consent had been obtained. Cryostat sections were made and laser microdissection was used to isolate tumor cells (PALM MicroBeam; Zeiss, Munich, Germany). Total RNA was extracted from these cells and also from RCC-26 cells (RNeasy micro kit; Qiagen, Hilden, Germany). RNA integrity was tested by capillary electrophoresis (2100 Bioanalyzer; Agilent, Santa Clara, CA). Thereafter, RNA amplification and hybridization on oligonucleotide microarrays (GeneChip Human Genome U133 Plus 2.0 array; Affymetrix, Santa Clara, CA) were performed with the manufacturer's reagents in accordance with the standard protocol. Normalized expression values were calculated for every transcript, using dChip software (Li and Wong, 2001) (A. Buchner, unpublished data). Expression levels of survivin, cyclin D1, adipophilin, and c-Met were quantified by quantitative real-time polymerase chain reaction (qRT-PCR) according to methods described elsewhere (Frankenberger et al., 2005a). Primer sequences are available on request.
Immune monitoring
Blood samples from the patients were drawn at study weeks 1, 6, 14, 22, and 36, and peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation using Leucosep (Greiner Bio-One, Frickenhausen, Germany) and cryopreserved. Additional samples from five patients were taken on day 15 after the last (tenth) vaccination and samples were available from two patients during longer follow-up. For interferon (IFN)-γ ELISPOT assays, the cells were thawed, washed with CTL Wash supplement medium (Cellular Technology, Cleveland, OH) plus Benzonase nuclease (Novagen Merck Biosciences, Darmstadt, Germany), rested, and seeded at 1.5 × 105 cells in quadruplicate on antibody-precoated polyvinylidene difluoride (PVDF) plates (Mabtech, Nacka, Sweden). The PBMCs were stimulated directly with selected peptides (each peptide, 5 μg/ml) in serum-free culture medium (CTL Test medium; Cellular Technology), supplemented with CD28 (1 μg/ml; BD Biosciences, San Jose, CA) and recombinant IL-2 (Proleukin, 2 U/ml; Chiron, Emeryville, CA). The assays were developed as previously described (Brill et al., 2007). Spots were counted with AID reader system ELR03 with software version 4.0 (Autoimmun Diagnostika [AID], Strassberg, Germany) and controlled by human audit. The performance of the assay was externally validated by participation in the immune assay proficiency panels of the international Cancer Vaccine Consortium at the Cancer Research Institute, New York, USA (Janetzki et al., 2008) and the Association for Immunotherapy of Cancer (CIMT), Mainz, Germany (Britten et al., 2008). Definition of positive responses was based on proposed international criteria (Britten et al., 2008; Janetzki et al., 2008).
Surrogate peptides for immune monitoring were selected from sequences of TAAs shown to be overexpressed in metastatic RCC lesions and/or the vaccine cells, using HLA-A*0201 motif-based epitope predictions available on the web (
Peptide synthesis and vaccine cell lysate preparation were performed as described previously (Brill et al., 2007). An irrelevant peptide for Bcr3/Abl2 [SSKALQRPV926–934 (Yotnda et al., 1998)] was used as a negative control and the HLA-A2 CEF (human cytomegalovirus, Epstein–Barr virus, and influenza virus) peptide pool (CMVpp65, NLVPMVATV495–503; EBV-BMLF1, GLCTLVAML280–288; EBV-LMP-2, CLGGLLTMV426–434; influenza M1 protein, GILGFVFTL58–66; and influenza RNA polymerase PA, FMYSDFHFI46–54), with 0.2 μg of each peptide per well, was used as a positive control (PANATecs, Tübingen, Germany).
Statistics
Time to progression and tumor-specific survival were analyzed by the Kaplan–Meier method.
Results
Characteristics of study patients
Table 1 summarizes the characteristics of the patients enrolled in this study. There were more male than female participants (11 males vs. 4 females) and the patients ranged in age from 39 to 78 years (mean, 59 years) at study entry. In nine patients, the time from first diagnosis to disease progression and study entry was 2 years or less. This interval was 3–12 years for six patients, with three patients showing disease progression only after 8, 11, and 12 years. The TNMG classification of the tumors was variable. At the time of enrollment, the majority of patients had multiple metastases at two or more sites, revealing their advanced states of disease. All but two patients had lung lesions and the second most prevalent site of metastasis was the lymph node. According to the study protocol, all patients had good performance status (ECOG scores of 0 or 1). Patients were also evaluated for prognosis according to the Palmer scoring criteria (Palmer et al., 1992). Seven patients had good scores of 0 or 1, whereas eight patients were classified with poor scores of 2 or 3, according to the designated criteria of performance status, number of metastatic sites, and time from first diagnosis to study entry.
Abbreviations: F, female; LN, lymph node; M, male; TNMG: tumor size, node involvement, metastasis, grade.
Score from 0 (very low risk) to 3 (high risk); based on performance status, number of metastatic sites, and time from first diagnosis to study entry (Palmer et al., 1992).
Cytokine: interferon-α, interleukin-2, 5-fluorouracil.
G250: monoclonal antibody directed against carbonic anhydrase-9.
Vaccine: multipeptide vaccine or dendritic cell vaccine.
The primary tumors of all patients were surgically removed at the time of primary tumor diagnosis. Ten patients were treated subsequently with some form of immunotherapy, with the majority receiving cytokine therapy in the form of subcutaneous IFN-α and IL-2, with or without 5-fluorouracil. Three patients received anti-G250 antibody therapy and one patient was treated with the multi-tyrosine kinase inhibitor sorafenib, which was stopped after only a few days because of severe skin reactions. Two patients were included in previous vaccine trials using either tumor lysate-pulsed dendritic cells (Holtl et al., 2005) or a pool of HLA-A2-restricted, tumor-associated peptides.
Vaccination schedule and clinical outcome
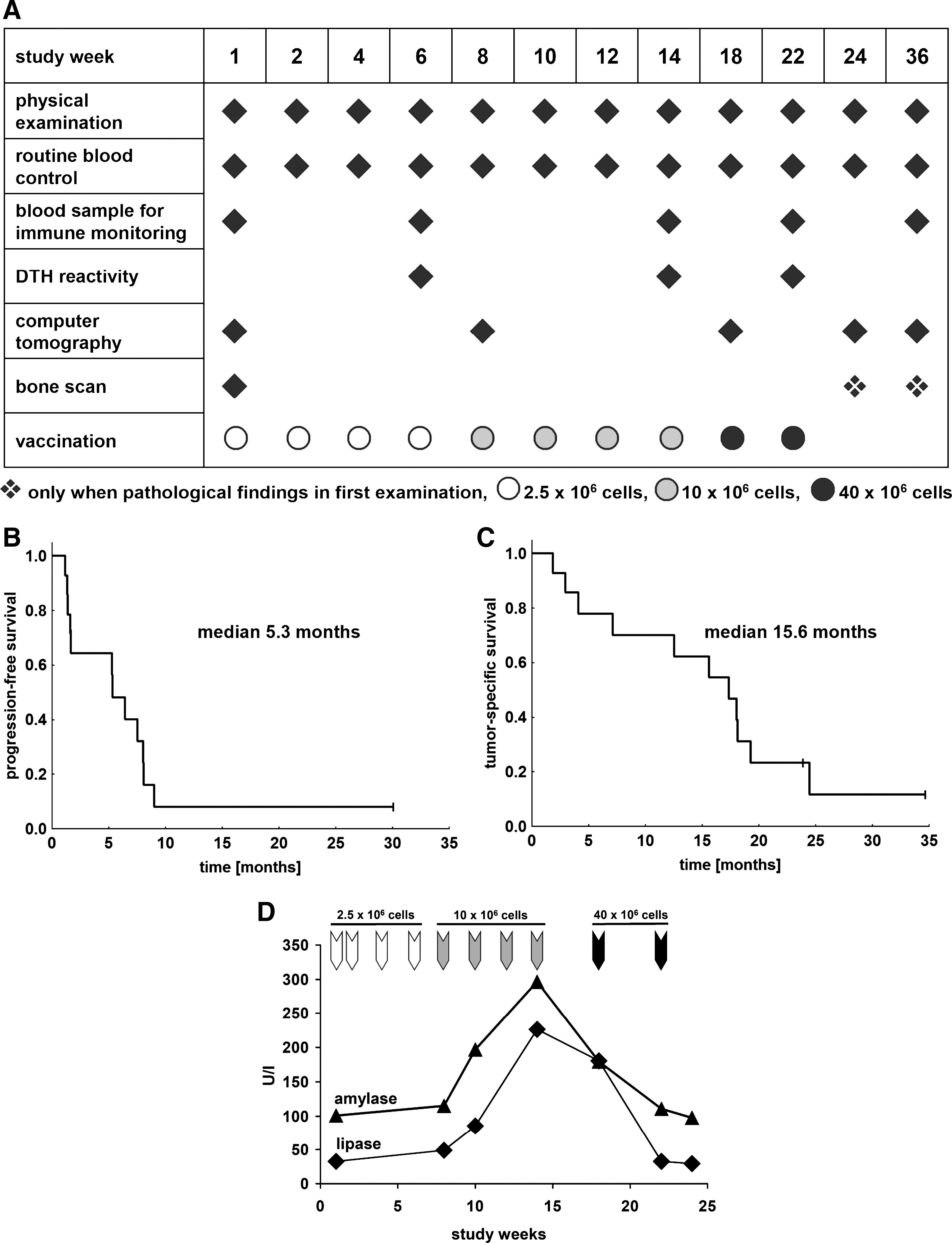
Figure 1A shows the vaccine dose and time schedule as well as points of clinical restaging, measurement of DTH reactions, and acquisition of samples for subsequent immune monitoring. The vaccination course for individual patients and various parameters of clinical outcome are summarized in Table 2. There was no difference in outcome between the four patients (MR-6, MR-7, MR-8, and MR-9) who received the short vaccination schedule that excluded the four low-dose vaccinations and the remaining patients. Patient MR-3 died 1 week after the second vaccination but no causal relationship was found between vaccination and death in this elderly patient. Patient MR-11 died in study week 17, 3 weeks after receipt of the eighth vaccine application. This patient had stable disease at the previous restaging according to RECIST criteria, and there was no evidence of a causal relationship with vaccination.
(
Abbreviations: DOD, dead of disease.
New metastases.
Stable disease at this time.
No remissions were observed in any patient according to RECIST criteria. In various patients tumor progression occurred at various sites, including the appearance of new lesions in the brain, bone, liver, adrenal glands, and lymph nodes in various individuals. Interestingly, progression of lung metastases was detected in only one instance and this patient showed a slow progression course (MR-4). At the end of the 36-week study period, 60% of the patients were alive. The median time to progression was 5.3 months (Fig. 1B). The median tumor-specific survival time was 15.6 months (Fig. 1C) and two patients are still alive to date, 24 and 35 months after study entry.
Analysis of safety and toxicity
The vaccine was well tolerated and no patient was withdrawn from the study because of side effects. The most common side effects noted were erythema, induration, and pruritus for a few days at the vaccine injection sites or DTH challenge sites. In most cases these responses occurred only after four or more vaccine applications. No local or systemic therapy was needed for treatment of adverse events in any study patient. In most cases, routine blood parameters (e.g., differential blood count, creatinine, electrolytes, and transaminases) showed no significant changes during the entire study period. Four patients (MR-9, MR-13, MR-14, and MR-15) showed transient increases in α-amylase and/or lipase, reaching up to 5-fold upper reference levels, with complete normalization after several weeks. The earliest detection of enzyme increase was after the fifth vaccination, when the vaccine dose was increased to 10 × 106 cells and vaccination applications were given at 2-week intervals (Fig. 1D). Further increases were not detected when the highest vaccine doses were applied at 4-week intervals. The enzyme increases were not accompanied by clinical symptoms and additional clinical examination revealed no evidence of pancreatitis. Moreover, no patient showed evidence of clinically relevant autoimmunity, as assessed by standard laboratory parameters.
Immunological assessment of vaccine responses in vivo
DTH reactions were detected in 11 of 12 evaluated patients after challenge with vaccine cells applied intradermally in the inguinal region at weeks 6, 14, and 22. Furthermore, most patients reported similar reactions at the sites of regular vaccine injections, first appearing after the receipt of at least three vaccine applications. The DTH challenge reactions were first assessed after patients had received four vaccine applications. Subsequent DTH challenge reactions increased in size and intensity with further vaccination. Reactions always disappeared within a few days and did not require medical intervention. Patient MR-6 failed to develop a DTH response even though he received four vaccine applications.
Immune monitoring of lymphocyte responses ex vivo
Sequential patient peripheral blood samples (see Fig. 1A) were obtained throughout the study period and IFN-γ ELISPOT analyses were performed to compare lymphocyte responses to a variety of stimuli before and after vaccination. These studies were performed with various numbers of samples from 12 patients, dependent on the course of vaccination completed by each individual. Data were not available for patients MR-3, MR-8, and MR-12, who did not reach the time point for obtaining the first immune monitoring sample after four vaccine applications at either the low or intermediate cell doses (see Table 2 and Fig. 1A).
To determine the general capacity of patient PBMCs to respond to antigenic stimulation, IFN-γ responses were assayed with a pool of HLA-A2-restricted viral peptides derived from proteins of cytomegalovirus, Epstein–Barr virus, and influenza virus, designated as the CEF peptide pool. Most HLA-A2 healthy donors have some memory T cells that respond to at least one of the peptides included in this pool (Currier et al., 2002). Responses to CEF peptides were detected in prevaccination samples of all 12 patients (data not shown). Positive responses to CEF peptides were also detected in 37 of 38 patient blood samples obtained postvaccination, indicating that overt immune suppression capable of blocking all capacity for memory immune responses did not occur despite disease progression in most patients. However, five of nine patients evaluated 22 or 36 weeks after study entry did show substantial decreases in the magnitude of their CEF responses (data not shown). This, however, was not indicative of a shorter period of time to progression.
Vaccine cell lysates were used to determine immune reactivity as a whole to vaccine-associated determinants (Fig. 2A). Interestingly, several patients showed substantial responses to vaccine lysates in PBMCs obtained before vaccination. Nevertheless, the median and mean ELISPOT responses in nine patients increased substantially in the postvaccination samples in comparison with prevaccination levels. The median and mean differences between the pre- and postvaccination responses of the nine immune responders were determined and depicted as delta (Δ) values. This revealed a mean change of 177 spots, with a median of 120 in the nine responding patients (Fig. 2A).

Monitoring of immune responses by ELISPOT to vaccine lysate and surrogate peptides and peptide pools. (
To determine whether TAA-specific responses were induced as a result of vaccination, ELISPOT responses were tested with a panel of surrogate peptides derived from TAAs that are prevalent in RCC or other tumors. The TAAs used for this analysis were tested for their prevalence and expression levels, using a microarray analysis of 32 mRCC lesions derived from a local tissue biorepository. RCC-26 cells were also included in this analysis (Fig. 3). All but one of the selected TAAs were expressed in the majority of tumor samples. Several TAAs were expressed in metastatic lesions but were not detected in RCC-26 cells. Nevertheless, they were included for future studies assessing epitope spreading. Several TAAs that were negative or poorly expressed in tumor samples and not expressed in RCC-26 cells were included as negative controls. It was not possible to perform similar array studies with patient tumors because the patients were recruited from throughout Germany and most entered the study several years after their primary diagnosis. Therefore, cryopreserved archival material was not available and for ethical reasons the protocol did not allow new biopsies to be made from the lung, bone, or liver metastases of these patients with advanced disease. Nevertheless, the analysis of metastatic RCC lesions obtained from the local tissue biorepository showed broad expression of most TAA candidates in a representative mRCC patient population.

Expression of tumor-associated antigens in metastatic RCC lesions and RCC-26 cells. Given are percentages of samples positive for selected candidate antigens in a panel of 32 metastatic RCC lesions and normalized levels of antigen expression in microdissected tumor cells from the metastatic lesions and in RCC-26 cells as determined by microarray analysis. The scale for the model-based expression index is as follows: –, no expression; +, <100; ++, 100–999; +++, ≥1000.
The immune-monitoring peptides derived from these 17 TAAs all carried anchor residues that allowed them to be presented by autologous HLA-A2 molecules (see Materials and Methods). Because of the high number of peptides to be evaluated, pools B and D, which contained multiple peptides, were created. Subsequent to vaccination, the responses of individual patients varied over time and various patients showed prominent responses to the two pools. Representative responses are summarized in Table 3, in which prevaccine values are juxtaposed to the strongest recorded responses detected at any time point postvaccination. An HLA-A2-restricted peptide derived from the Bcr/Abl fusion protein (Yotnda et al., 1998) was used as a background control and only low numbers of spots (median, 0 spots; range, 0 to 29 spots) were detected in pre- and postvaccination samples (data not shown). T cell responses were also measured against four single peptides, survivin, cyclin D1, adipophilin (adipose differentiation-related protein [ADFP]), and c-Met, which were shown previously by others to be associated with T cell reactivity in patients with RCC (Stevanovic, 2002). Seven of 12 patients showed responses to either the single peptides and/or the pools, with all seven immune responders showing responses to two or more peptides/pools that were greater than 2-fold compared with prevaccination values (Fig. 2B). It should be noted that most patients showed substantial prevaccination responses to one or both pools and several of the single peptides, indicating that their PBMCs contained effector memory cells. The TAAs from which the four single peptides were derived showed variations in mRNA expression levels in the vaccine cells, but this did not impact on the number of patients capable of responding to the individual peptides (Fig. 2C). The time courses of responses to the single peptides and pools B and D are illustrated for two representative patients (Fig. 2D and E). In general, responses to the individual stimuli (pools or single peptides) showed similar patterns of increase or decrease throughout the vaccine schedule.
Abbreviations: DTH, delayed-type hypersensitivity reaction; SD, stable disease.
Pool B contains the peptides for RGS-5, apolipoprotein, MUC-1, OFA, hTERT, and KIAA0367 (PRUNE2).
Pool D contains the peptides for VEGF, NY-ESO, Her2/neu, MAGE-1, −3, −6, and WT-1.
The values represent absolute numbers of spots for pre- and postvaccination (pre/post) responses. These values are given after subtraction of background responses (i.e., without peptide or with irrelevant Bcr/Abl peptide). The highest postvaccination response is given irrespective of the time point postvaccination (i.e., at 6, 14, 22, or 36 weeks).
No values mean no reactivity (≤10 Spots above background, which corresponds to a frequency of ≤1/1.5 × 104 IFN-γ-secreting peptide-specific T cells).
Values representing a greater than 2-fold increase in reactivity against the peptides post-versus prevaccination are indicated in bold. With the larger peptide pools a reactivity of ≥40 spots above background was considered positive.
Short protocol of vaccination starting with the middle dose of 107 vaccine cells.
SD (7) and SD (131) indicate stable disease at weeks 7 and 131, respectively.
Discussion
Individualized RCC cell vaccines offer the possibility to target immune responses to a unique array of epitopes that arise through genetic mutation in tumor cells; however, a parallel compensation must be made to improve the inferior immune-stimulatory capacity of tumor cells. Today, preparation of patient-individualized tumor vaccines entails obtaining tumor samples from individual patients, genetically engineering them under stringent GMP conditions, and performing quality assurance studies to demonstrate that sufficient numbers of cells show adequate levels of gene expression to achieve the desired immune effects. Combined, all of these steps require substantial amounts of time, result in high costs, and yield potential limitations in vaccine availability for individual patients, particularly if multiple vaccine applications with large numbers of cells are deemed to be beneficial.
Our vaccine strategy was designed to overcome these logistical hurdles by relying on the use of a generic vaccine that could be applied in multiple patients. For vaccine development, we used a well-characterized RCC-26 tumor cell line that showed strong immunogenic potential in extensive preclinical studies (Schendel and Gansbacher, 1993; Schendel et al., 1993, 2000; Frankenberger et al., 2005a,b). This inherent immune-stimulatory potential was improved through gene modification to allow the RCC-26 cells to express surface CD80 molecules in order to improve their capacity to induce T cell responses through provision of secondary activation signals to responding lymphocytes. This strategy was supported by our preclinical experimental mouse tumor models, which clearly demonstrated that CD80 expression on tumor cells induced cytotoxic T lymphocytes by direct priming (Cayeux et al., 1997). Furthermore, CD80 expression on RCC-26 cells supported reactivation of effector memory T cells that were present in patients with RCC (Schendel et al., 2000).
Gene modification of RCC-26 cells to secrete IL-2 was chosen because of the well-known clinical benefits of IL-2 achieved in some patients with RCC by systemic therapy (Rosenberg, 2007). Our vaccine cells secreted only low levels of IL-2 (8 ng of IL-2 per 106 cells per 24 hr) and thus systemic effects were not expected. In fact, it was shown previously that the amount of IL-2 produced by vaccine cells has a strong impact on the generation of tumor immunity. If vaccine cells released high amounts of IL-2, tumor immunity could be completely abrogated (Schmidt et al., 1995). Our preclinical studies of IL-2-modified RCC-26 cells showed clearly that low levels of IL-2 could indeed specifically support activation of effector memory cells. This was manifested in the remobilized effector lymphocytes by increased proliferation, enhanced cytotoxic function against tumor cells, and improved survival through upregulation of antiapoptosis genes in the remobilized effector lymphocytes (Frankenberger et al., 2005b). Furthermore, IL-2 expression by the vaccine cells supported the activation of natural killer cells (Schendel and Gansbacher, 1993).
Moreover, the vaccine cells could further impact the development of immune responses through their capacity to induce alloresponses due to expression of mismatched HLA allotypes with study patients, providing a rich cytokine milieu as well as nonspecific help through recruitment of CD4+ T cells that could support the development of antitumor immunity (Schendel et al., 2000). We showed previously that ongoing alloresponses stimulated by the vaccine cells did not suppress T cell responses to RCC-associated epitopes (Frankenberger et al., 2005a).
Because this was a generic vaccine designed for use in patients who expressed the HLA-A*0201 allotype, only one vaccine development phase was needed. This allowed extensive characterization of the vaccine cells postproduction according to GMP standards to certify that the genetically engineered cells presented excellent characteristics with respect to gene expression and immunological function (Frankenberger et al., 2005a). Thereafter, there were no time constraints for patient use and the amount of vaccine was not a limiting factor. The vaccine cells were available in an irradiated, cryopreserved formulation that allowed direct application to patients without manipulation after thawing. The clinical status of the patient was the primary factor that influenced the number of vaccine applications that could be delivered during the study period.
Because only one tumor cell line was employed, it was also possible to carry out extensive studies regarding expression of a variety of TAAs in the vaccine cells. This allowed the identification of a series of TAAs that were overexpressed in RCC-26 cells and made it possible to devise specific immune-monitoring tools to study the induction or reactivation of TAA-specific T cells. Surrogate peptides, representing epitopes derived from several overexpressed TAAs, were selected for their capacity to be presented by HLA-A2 molecules. They were tested either in pools or as single peptides to measure T cell responses in patients throughout the study course. This enabled several parameters of response to be assessed, providing a better picture of the development of specific immune responses generated through vaccination in the study group.
This form of tumor cell vaccination was found to be feasible and safe. The designated vaccine course was completed in 9 of 15 patients. Despite heavy tumor burdens at study entry, seven patients completed a vaccination schedule spanning 22 weeks. The majority of patients had multiple metastases at two or more sites at enrollment, and more than half of the patients had poor prognosis scores according to Palmer and colleagues (1992), revealing their advanced states of disease. Those patients who progressed too rapidly to complete the vaccine protocol had poor prognosis scores of 2 or 3, according to the criteria of Palmer and colleagues.
The vaccine was also determined to be safe, even when applied at doses of 40 × 106 cells. Although vaccine site reactions and DTH challenge reactions increased in magnitude with increasing vaccine applications, this did not prohibit 10 vaccine applications in the course of 22 weeks. These responses were transient, disappeared within a few days, and did not require medical intervention. The most serious vaccine-associated toxicity was the transient increase in amylase and lipase enzyme levels in four patients. These increases were first detected after the fifth vaccine injection. They increased during the subsequent vaccine applications given at 2-week intervals but then disappeared, despite further increases in cell dose. Thus, these transient increases seemed to be related more to the time interval between vaccine applications than to cell dose. These increases were not accompanied by other systemic symptoms and signs of pancreatitis were not detected by clinical examination, imaging, and measurement of additional blood parameters. Development of autoimmunity was assessed by the appearance of anti-nuclear antibodies and rheumatoid factor or changes in complement factors. No relevant changes were noted in any patient throughout the study course.
All patients had progressive disease before they entered the study, except one patient (MR-14) who was enrolled previously in a peptide vaccine trial (A. Buchner, unpublished observation). Of the 14 progressing patients, half showed stable disease for time periods ranging from 23 to 131 weeks. The median time to progression was 5.3 months and the median tumor-specific survival time was 15.6 months. Interestingly, studies of much larger groups of patients showed progression-free survival times of 5.5 months with the angiogenesis inhibitor sorafenib (Escudier et al., 2007) and 5 months with IFN-α (Motzer et al., 2007); however, the side effects of these therapies were much greater. Progression was detected at various sites and new lesions appeared in various organs. It should be noted that although 13 patients had lung metastases at study entry, further tumor progression at this site was observed in only 1 patient (MR-4). In addition, the progression of multiple lung lesions in this patient was slow. It has been observed in other studies that lung metastases may respond better to immunotherapies than metastases at other sites (Han et al., 2003).
Immune monitoring revealed that vaccine-induced T cell responses could be detected in a majority of the patients. Strong reactivity to the control viral CEF pool was detected in prevaccination samples of all evaluated patients, indicating that their memory T cells were capable of responding to antigenic stimulation despite extensive tumor burden that is often associated with a general state of immune suppression (Frankenberger et al., 2007). Responses to CEF peptides were retained in seven of nine patients postvaccination at 22 weeks. However, spot numbers were lower in three patients at 36 weeks compared with values obtained at weeks 1, 6, and 14, indicating some loss of immune potential. There were also surprisingly high responses to vaccine lysates in the prevaccination samples and these increased in nine patients through vaccination. Because the responding lymphocytes in the ELISPOT assays were not separated into natural killer (NK) and T cell fractions, it is possible that some of this reactivity was due to NK cells, which were shown previously to respond to RCC-26 cells in the presence of IL-2 (Schendel et al., 2000; Falk et al., 2002; Frankenberger et al., 2005b). On the other hand, there is substantial evidence that patients with RCC spontaneously develop T cell responses to their tumors. Thus, some of the response to lysate stimulation may reflect responses of effector memory T cells. Because RCC-26 cells overexpress a large number of various TAAs, such T cell responses may be specific for TAAs that are shared by various RCCs. Further evidence of the presence of preexisting effector memory T cells in some patients was reflected in the ELISPOT responses detected to the pools and single peptides in prevaccine PBMCs.
More than half of the patients showed vaccination-associated increases in T cell reactivity to surrogate peptides that were derived from TAAs overexpressed by the vaccine cells and commonly shared by RCC (Stevanovic, 2002). The patterns of response to the single peptides and peptide pools were diverse, indicating that an immunodominant epitope was not present in this set of surrogate peptides. Furthermore, several patients responded to two or more specific peptides or pools, indicating that several epitopes expressed by the vaccine cells could elicit ELISPOT responses in these patients with advanced disease. However, no correlation was found between degree of immune response and stabilization of disease as reflected by time to progression or survival times. Further studies are needed to elucidate the individual specificity of responses occurring to the two peptide pools. This will then provide a more complete picture of the specificity and complexity of T cell responses, including the role of epitope spreading, occurring as a result of vaccination.
Although no clinical responses according to RECIST were detected, the median survival time seen in this small patient group was long, considering their clinical status at study entry. Because of the minimal side effects, vaccination was well tolerated and could be delivered on an outpatient basis throughout the study course. The use of a generic vaccine strategy for selected HLA-A*0201-positive patients contributed substantially to the feasibility of employing an escalating, multidose vaccine strategy because vaccine cells were not limiting. This generic approach also allowed a standardized immune-monitoring approach to be used for all study patients. On the basis of the preliminary results presented here, it can be concluded that the majority of patients acquired increased T cell reactivity through vaccination to several epitopes derived from shared TAAs. Such T cell responses may be able to elicit significant immune attacks against autologous tumor lesions in better clinical settings. This could include, for example, combining vaccination with the application of treatment strategies that reduce regulatory cells that can dampen ongoing immune responses. Furthermore, combination therapies employing vaccination and new molecular inhibitors to block tumor-driven angiogenesis and retard tumor progression may allow better immune responses to develop over longer periods of time.
The demonstrated feasibility of applying this vaccine in a group of patients at fairly low cost, with substantial savings in time and logistics as compared with individualized patient vaccines—alongside safety, low toxicity, and substantial immune-stimulatory potential—supports performing further evaluations of this vaccine in combination therapies and as adjuvant therapy in patients with minimal residual disease.
Footnotes
Acknowledgments
This study was supported by the Federal Ministry of Education and Research (01 GE 9624/1) and the German National Research Foundation (SFB-455 and SFB-TR36). The authors thank Roswitha Fischer, Heidi Herbig, and Birgit Stadlbauer for excellent technical support in immune monitoring.
Author Disclosure Statement
No competing financial interests exist.