
Abstract
Growth hormone (GH) deficiency (GHD) causes somatic growth impairment. GH has a short half-life and therefore it must be administered by daily subcutaneous injections. Adeno-associated viral (AAV) vectors have been used to deliver genes to animals, and double-stranded AAV (dsAAV) vectors provide widespread and stable transgene expression. In the present study we tested whether an intramuscular injection of dsAAV vector expressing GH under the control of a muscle creatine kinase regulatory cassette would ensure sufficient systemic GH delivery in conjunction with muscle-specific expression. Virus-injected GHD mice showed a significant (p < 0.05) increase in body length and body weight, without reaching full normalization, and significant (p < 0.05) reduction in absolute and relative visceral fat. Quantitative RT-PCR showed preferential GH expression in skeletal muscles that was confirmed by qualitative fluorescence analysis in mice injected with a similar virus expressing green fluorescent protein. The present study shows that systemic GH delivery to GHD animals is possible via a single intramuscular injection of dsAAV carrying a muscle-specific GH-expressing regulatory cassette.
Introduction
Somatic growth requires the presence of growth hormone (GH), which is secreted into the circulation by the somatotroph cells of the anterior pituitary gland. GH exerts its effects both directly and indirectly through the action of locally produced and circulating insulin-like growth factor-I (IGF-I) (Le Roith et al., 2001). Growth impairment and metabolic abnormalities present in children with GH deficiency (GHD) are reversed by replacement treatment with GH (rt-GH) (Bryant et al., 2002). The short half-life of GH (Giustina and Veldhuis, 1998) requires that patients be treated with daily subcutaneous injections (Saggese et al., 1998). We have developed a mouse model of isolated growth hormone deficiency (IGHD) by generalized ablation (knockout, KO) of the GH-releasing hormone gene (GHRHKO) (Alba and Salvatori, 2004) and we have showed that, in these animals, both somatic growth and body composition were normalized by recombinant mouse GH injections (Alba et al., 2005).
Viral vectors have been used to effectively deliver proteins over a long period of time. Gene therapy approaches have been tested in GHD animals, using recombinant adenovirus vector (Hahn et al., 1996; Marmary et al., 1999) or single-stranded adeno-associated viral (ssAAV) vectors expressing GH (Rivera et al., 1999). Double-stranded adeno-associated viral (dsAAV) vectors offer several advantages over ssAAV vectors, as they can incorporate dsDNA and are generally characterized by low toxicity while providing long-term gene transfer, strong infectivity, and stable expression (Wang et al., 2003).
We have previously used the intraperitoneal injection of dsAAV vector (serotype 8) to deliver GH under the control of a ubiquitous cytomegalovirus (CMV) promoter to GHRHKO mice, which reached full growth normalization 4 weeks after infection (Sagazio et al., 2008). GH mRNA detection in liver, cardiac, and skeletal muscles was consistent with the GH gene being under the control of a nonspecific promoter, and the lack of GH expression in other tissues was related to the tissue tropism of the virus itself (Wang et al., 2005). However, widespread gene expression can result in toxicity (Weeratna et al., 2001; Wang et al., 2004). In addition, direct liver exposure to recombinant AAV vector in humans has caused destruction of transduced hepatocytes by a cell-mediated immune response elicited by antigens of the AAV capsid, resulting in both a decline in gene expression and transient transaminitis (Manno et al., 2006). In addition, the risks due to disproportionate GH production, as seen in subjects with GH-secreting pituitary adenomas, are well documented (Melmed, 2006). For these reasons, the use of a site of injection that does not expose the liver to high doses of virus and of a promoter with nonuniversal expression would be desirable.
Skeletal muscles are attracting interest as administration sites for gene therapy, as they are easily accessible, composed of myofibers with a long half-life, well vascularized, and suitable for production and secretion of proteins (Marshall and Leiden, 1998). Regulatory cassettes derived from promoter and enhancer portions of muscle-specific genes—especially muscle creatine kinase (M-CK) (Donoviel et al., 1996; Shield et al., 1996; Hauser et al., 2000)—have been used to avoid or strongly limit gene expression in nonmuscle tissues (Cordier et al., 2001; Salva et al., 2007). An additional safety benefit of CK regulatory cassettes is that they function only in terminally differentiated postmitotic striated muscle cells. This greatly lessens the possibility that the unintended and rare genomic integration of a CK regulatory cassette into a replicating myoblast or other proliferative cell types would cause deleterious expression from adjacent genes. A large number of CK cassettes have been developed with different selectivity and potency, finding applications in the expression of genes in skeletal and heart muscles for the treatment of muscular dystrophies in mice (Wang et al., 2000; Salva et al., 2007) and murine Pompe's disease (Sun et al., 2008). Newer cassettes, although being compact in size (between 500 and 700 bp long), are stronger than viral promoters (Salva et al., 2007; Wang et al., 2008).
In the present study we tested whether selectively expressing GH in a single injected quadriceps muscle, using a dsAAV vector to deliver GH under the control of a newly designed CK-based regulatory cassette (CK8) (Q. Nguyen and S.D. Hauschka, unpublished data) would provide sufficient systemic GH to influence growth. To this end, we used the CK8 cassette because of its compact size and its higher activity compared with previously developed cassettes (Hauser et al., 2000; Salva et al., 2007). We show that this promoter is able to cause enough systemic GH delivery to significantly increase longitudinal growth in GHD mice.
Materials and Methods
Generation of dsAAV8-CK-GFP and dsAAV8-CK-GH vectors
The dsAAV8-CMV-green fluorescent protein (dsAAV8-CMV-GFP) and dsAAV8-CMV-GH vectors were described earlier (Xiao et al., 1998; Gao et al., 2002; Sagazio et al., 2008). The CK8 regulatory cassette was constructed by ligating an additional copy of the modified mouse M-CK enhancer region from the CK7 cassette (Salva et al., 2007) onto the 5′ end of the CK7 cassette. A full description of this small regulatory cassette is reported elsewhere (Nguyen and Hauschka, unpublished data). The CK8 cassette was cloned into dsAAV8-CMV-GFP (previously digested with MluI and PstI to release the CMV fragment) to give the dsAAV8-CK8-GFP vector. This was used to observe fluorescence in tissues that express the product of the plasmid construct.
Full-length mouse GH cDNA was amplified by polymerase chain reaction (PCR) from pituitary cDNA of a C57BL/6 mouse and cloned into dsAAV8-CK8-GFP (previously digested with KpnI and NotI to release the GFP fragment) to give the dsAAV8-CK8-GH plasmid. The orientation of the two constructs was confirmed by endonuclease digestion and DNA sequencing.
Recombinant viral stocks were produced by the adeno-virus free, triple-plasmid cotransfection method (Xiao et al., 1998). Briefly, twenty 150-mm dishes of human 293 cells (80% confluent) were prepared. A total of 60 μg of plasmid DNA (20 μg of dsAAV8-CK8-GFP or dsAAV8-CK8-GH plus 10 μg of pAAV8 plus 30 μg of pHelper) was dissolved in 2 ml of 0.25 M CaCl2 and quickly mixed with 2 ml of HBS buffer (50 mM HEPES, 280 mM NaCl, and 1.5 mM Na2HPO4, 10 mM KCl, 12 mM dextrose; pH 7.2), and then added to the cells of each dish. After 16 hr of transfection, the medium was replaced with fresh Dulbecco's modified Eagle's medium (GIBCO brand DMEM; Invitrogen, Carlsbad, CA) containing 10% fetal bovine serum and antibiotics. At 2 days posttransfection, the cells were harvested, resuspended in 15 ml of the supernatant, and subjected to four cycles of freeze–thawing. Cell debris was removed by low-speed centrifugation. The AAV particles were subsequently purified by two rounds of CsCl gradient ultracentrifugation. The titration of viral genome particle number was performed by quantitative DNA dot–blot method (Snyder et al., 1996).
Animals and vector administration
Only male animals were used for our experiments. Homozygous GHRHKO mice were treated with placebo (saline) or viral particles, whereas heterozygous (HTZ) male carriers were used as normal controls to guarantee a similar genetic background (mixed C57BL/6–129/Sv) (Alba and Salvatori, 2004). Viruses were injected by intramuscular injection in the right quadriceps muscle at day 10 of age. The low-dose (LD) group (n = 7) received 0.5 × 1011 dsAAV8-CK8-GH viral particles. The high-dose (HD) group (n = 7) received twice the number of viral particles (1 × 1011). The CMV group (n = 7) received 1 × 1011 particles of dsAAV8-CMV-GH via the same route. The GFP group (n = 7) received 1 × 1011 particles of dsAAV8-CK8-GFP. The placebo group (Plac) (n = 7) consisted of GHRHKO mice that received intramuscular placebo injections.
Pups were weaned at week 4 of age and they were killed at week 6 of age. Mice were housed in a controlled environment with 14-hr light/10-hr dark cycles at 21°C and 23% humidity, standard mouse/rat food (Prolab RMH2500; PMI Nutrition International, Brentwood, MO), and water ad libitum. All procedures were approved by the Johns Hopkins Institutional Animal Care Committee.
Auxological data
Mice were examined on a weekly basis for 6 weeks after birth. Total body weight (TBW) and body length (nose-to-anus distance, N-A) were measured with a daily-calibrated electronic balance (Scout Pro balance, SP601; Ohaus, Pine Brook, NJ) and with an electronic digital caliper (Fisher brand traceable digital caliper; Thermo Fisher Scientific, Hampton, NH), respectively. At the end of the study animals were anesthetized with Avertin (tribromoethanol; Sigma-Aldrich, St. Louis, MO) and blood for hormone measurements (∼200 μl) was collected by retroorbital bleeding. Sera were stored at −20°C until the day of assay. Euthanasia was performed by halothane (Sigma-Aldrich) overdose, and heart, liver, spleen, right kidney, right testis, right lung, right gastrocnemius (site of injection), and left quadriceps muscles were harvested after sacrifice. Internal organs were weighed with an Adventurer Pro analytical balance (AV264; Ohaus). Right femur and right tibia were cleaned and their length was measured with an electronic caliper. The perirenal and epididymal fat pads were pooled and weighed (visceral fat, VF) as well as fat pads from the interscapular and axillary region, thighs, and inguinal region (subcutaneous fat, SF). Lean mass (LM) was measured by weighing animals without tail, skin, adipose tissue, and internal organs. LM, VF, SF, and weights of internal organs were normalized to TBW according to the following equation: [weight (g)/TBW (g)] × 100.
Detection of GH RNA expression by quantitative RT-PCR
Total RNA was extracted with TRIzol reagent (Invitrogen) according to the manufacturer's recommendations. About 25–50 mg of tissue per mouse was used. Total RNA was spectrophotometrically quantified at 260 nm (DU 640 spectrophotometer; Beckman Coulter, Fullerton, CA) and 1 μg of total RNA was used to produce cDNA with reverse transcriptase (Moloney murine leukemia virus [M-MLV] reverse transcriptase; Promega, Madison, WI).
Real-time quantitative RT-PCR (RT-qPCR) was performed in a iQ5 multicolor real-time PCR detection system (Bio-Rad, Hercules, CA) and a master mix was prepared with iQ SYBR green supermix (Bio-Rad), a 1 mM concentration of each primer, and 2 μl of 1:5 diluted reverse transcriptase (RT) mix in a 25-μl reaction volume. After selection of the most adequate annealing temperature, sample assays and nontemplate controls were produced in duplicate for each gene. Primers used for the detection of GH mRNA were as follows: 5′-TACCTGCGGGTCATGAAG-3′ (forward) and 5′-GATGCATCTTAATTTTATTAGGAC-3′ (reverse). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control.
The following experimental run protocol was used: quantification program consisting of 40 cycles of 95°C for 10 sec, 58°C for 30 sec, and 72°C for 30 sec, ending with a melting program ranging from 55 to 95°C with a heating rate of 0.5°C/10 sec and continuous fluorescence measurement.
Serum hormone measurements
Serum GH was determined by radioimmunoassay (RIA) (rat GH RIA, RGH-45HK; Linco-Millipore, Billerica, MA). The assay included quality controls and standard samples provided by the manufacturer. Each sample was assayed in duplicate. The intraassay variability was determined to be 5.2%.
Serum IGF-I was determined with a mouse/rat IGF-I kit (DSL-2900; Diagnostic Systems Laboratories [DSL]/Beckman Coulter, Webster, TX) in accordance with the manufacturer's recommendations. The assay included quality controls and standard samples provided by the manufacturer. Each sample was assayed in duplicate. The intraassay variability was determined to be 2.2%.
Histological analysis
Tissues were fixed in 4% paraformaldehyde (PFA, pH 7.3), washed in 0.2 M phosphate buffer, embedded in Tissue-Tek O.C.T. compound (Sakura Finetek USA, Torrance, CA), and stored at −20°C. Seven-micrometer transverse sections were cut from frozen tissues. Tissue sections were placed on glass slides and VECTASHIELD mounting medium (Vector Laboratories, Burlingame, CA) containing 4′,6-diamidino-2-phenylindole (DAPI) was applied. GFP expression was directly evaluated under a fluorescence microscope. Tissues from mice in the Plac group were prepared according to the same procedure and used as controls.
Statistical analysis
Data analysis was performed by analysis of variance (ANOVA), using the SPSS statistical package (SPSS, Chicago, IL), with post hoc analysis according to the Bonferroni method. Data were considered statistically significant at p < 0.05.
Results
Auxological data
GHRHKO mice injected with dsAAV8-CMV-GH or dsAAV8-CK8-GH virus exhibited a significant increase in length and total body weight (TBW), but they did not attain the full size of normal 6-week control mice (Figs. 1 and 2). As expected, mice treated with dsAAV8-CK8-GFP were comparable to untreated mice. GHRHKO mice injected with the high dose of dsAAV8-CK8-GH virus and with dsAAV8-CMV-GH exhibited normal femoral and tibial lengths, whereas mice injected with the low dose of dsAAV8-CK8-GH virus had tibial and femoral lengths that were intermediate between those of HTZ and Plac mice (Fig. 3). Body composition in LD, HD, and CMV mice also changed after AAV-mediated GH delivery (Fig. 4). Absolute visceral fat (VF) was significantly decreased in mice treated with low and high doses of dsAAV8-CK8-GH virus as well as with dsAAV8-CMV-GH compared with Plac and GFP mice, whereas absolute subcutaneous fat (SF) was comparable among all the groups. Relative SF was also decreased in LD and HD mice compared with Plac and GFP mice, whereas relative VF was decreased in LD, HD, and CMV mice compared with Plac and GFP mice. Absolute and relative lean body mass (LM) appeared to change in virus-treated mice, but the difference from Plac was not statistically significant.
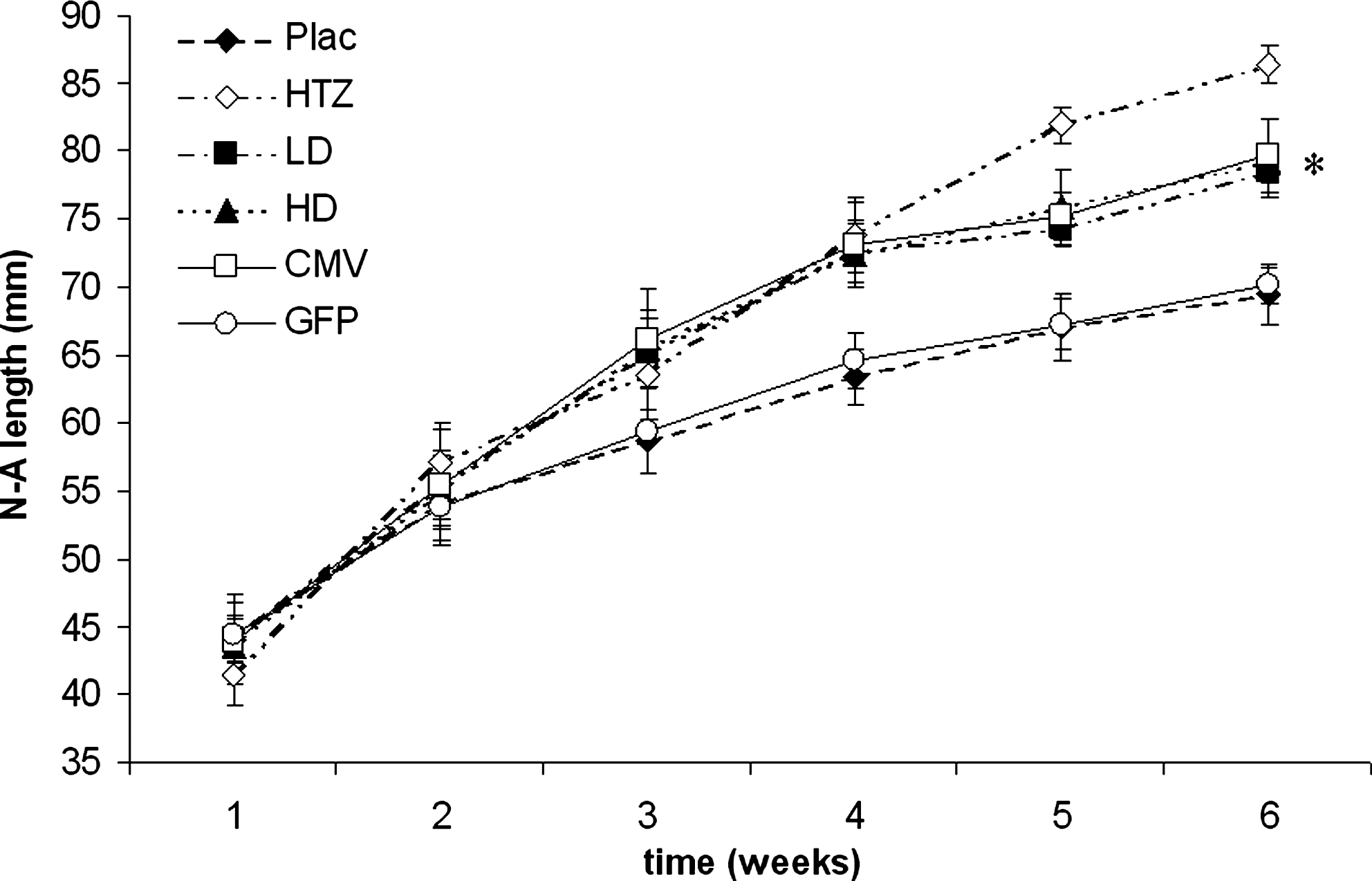
Body length (N-A, nose-to-anus distance, in millimeters) was assessed weekly and is expressed as means ± SD. LD, HD, and CMV mice reached a body length significantly higher than that of untreated mice (*p < 0.05 vs. Plac and GFP). CMV, cytomegalovirus; GFP, green fluorescent protein; HD, high dose; HTZ, heterozygous; LD, low dose; Plac, placebo.
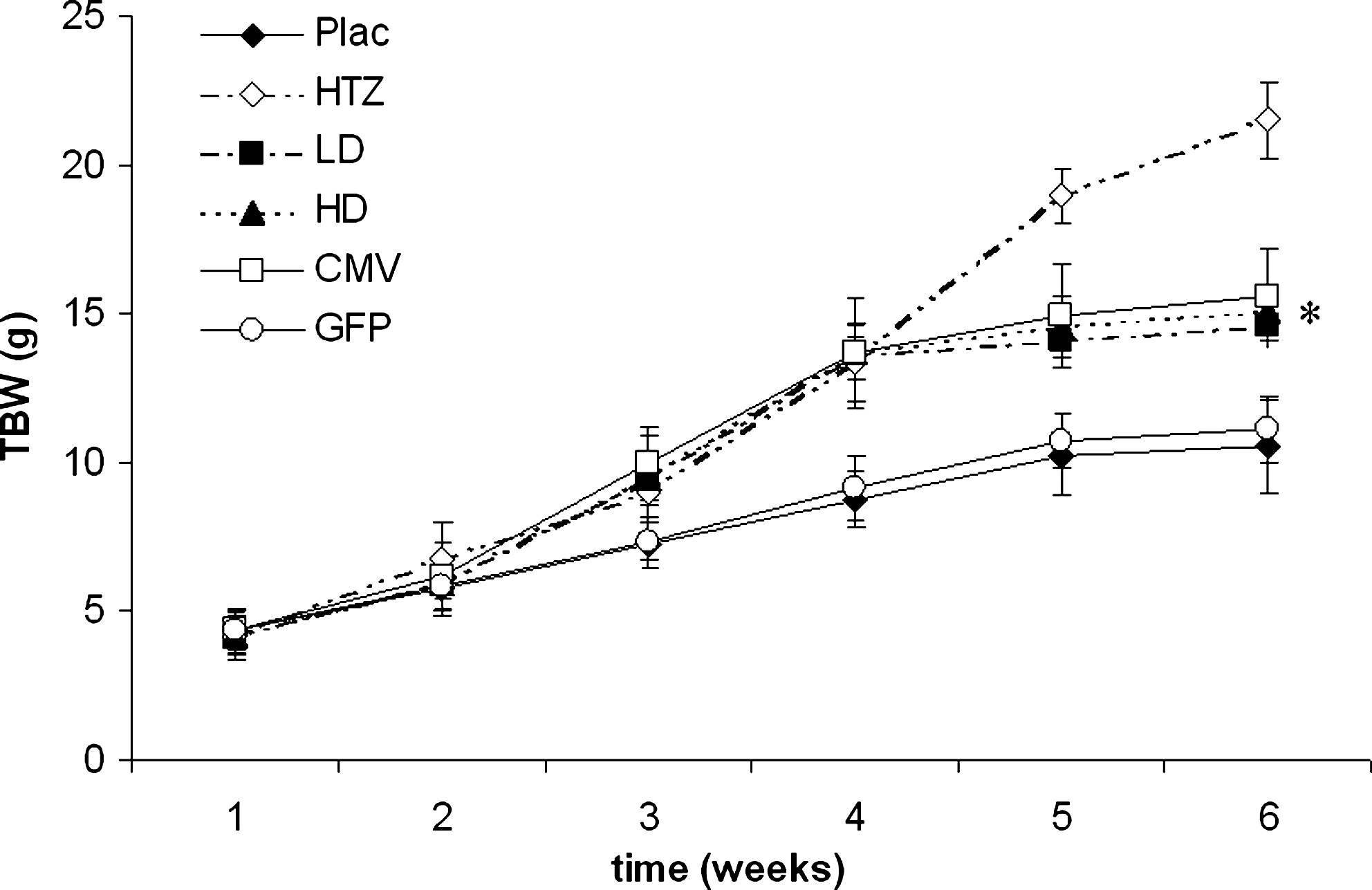
Total body weight (TBW, in grams) was assessed weekly and it is expressed as means ± SD. LD, HD, and CMV mice reached a TBW significantly higher than that of untreated mice (*p < 0.05 vs. Plac and GFP).
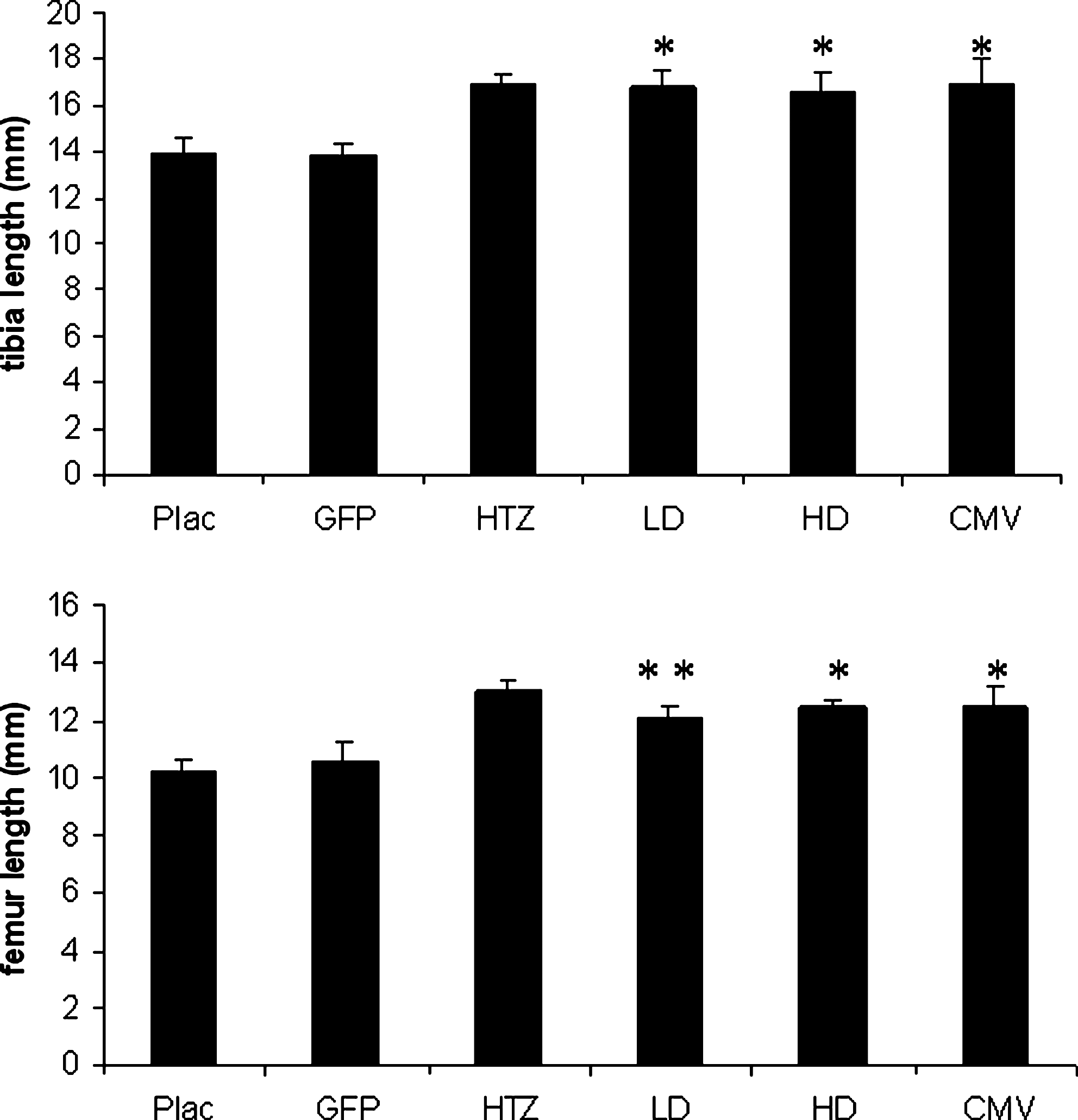
Tibial and femoral length (in millimeters) at time of death (means ± SD). LD, HD, and CMV tibias were significantly longer than those of Plac and GFP mice (*p < 0.01 vs. Plac). HD and CMV femurs appeared normalized compared with HTZ mice (*p < 0.01 vs. Plac) whereas LD femurs were significantly different from both Plac and HTZ femurs (**p < 0.02 vs. Plac and HTZ).
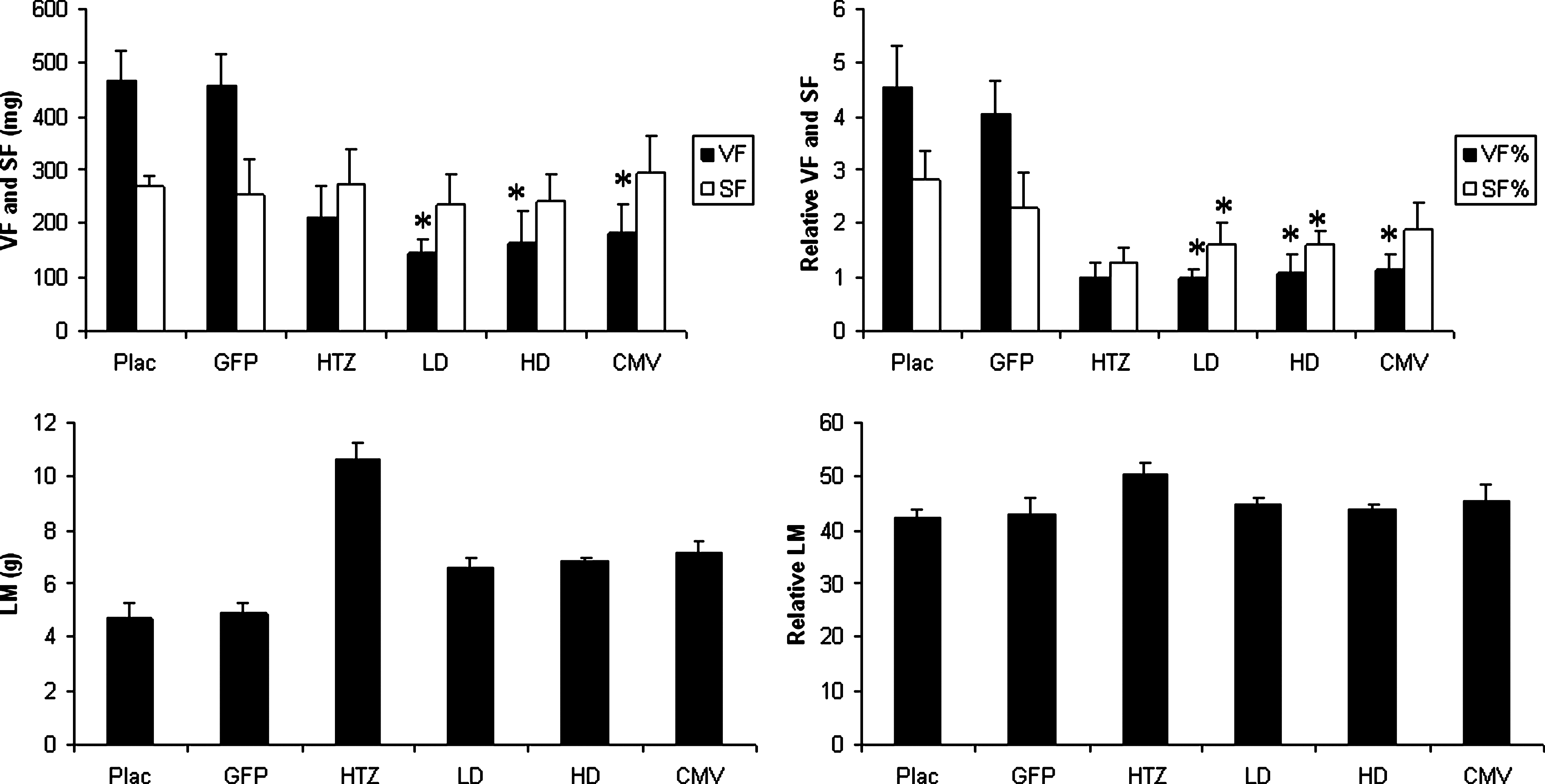
Body composition at the time of death (means ± SD). Absolute visceral fat (VF) was lower in LD, HD, and CMV mice compared with Plac mice (*p < 0.01 vs. Plac). Relative VF and subcutaneous fat (SF) were lower in LD and HD mice compared with Plac mice whereas only relative VF was lower in CMV mice (*p < 0.01 vs. Plac). Absolute and relative lean mass (LM) were not significantly different in treated and untreated mice.
Weights (measured as percentage of total body weight) of heart, kidney, testis, and lung did not differ between groups, showing a remarkable relation to body size. Liver size was lower in Plac and GFP animals than in HTZ animals, and remained lower in virus-injected animals, although HD and CMV animals had larger livers compared with placebo animals. Spleen size was lower in GFP and Plac mice and normal in LD, HD, and CMV mice when compared with HTZ mice (Table 1).
Weights of Selected Organs Normalized to Total Body Weight at the Time of Death a
Abbreviations: CMV, cytomegalovirus group; GFP, green fluorescent protein group; HD, high-dose group: HTZ, heterozygous group; LD, low-dose group; plac, placebo group; rx, right-side organ.
Values are expressed as means ± SD.
p < 0.01 versus HTZ.
p < 0.01 versus Plac.
GH mRNA expression
Quantitative RT-PCR showed that GH expression in quadriceps muscle of HD animals was significantly higher than in HD gastrocnemius and cardiac muscles, kidney, and liver (Fig. 5). GH expression in gastrocnemius muscle, cardiac muscle, kidney, and liver of HD animals was not significantly different from that in placebo mice. CMV mice showed significantly higher GH expression in quadriceps muscle compared with all other tissues. Also, GH expression in liver and gastrocnemius muscle of CMV mice was higher than in Plac tissues.
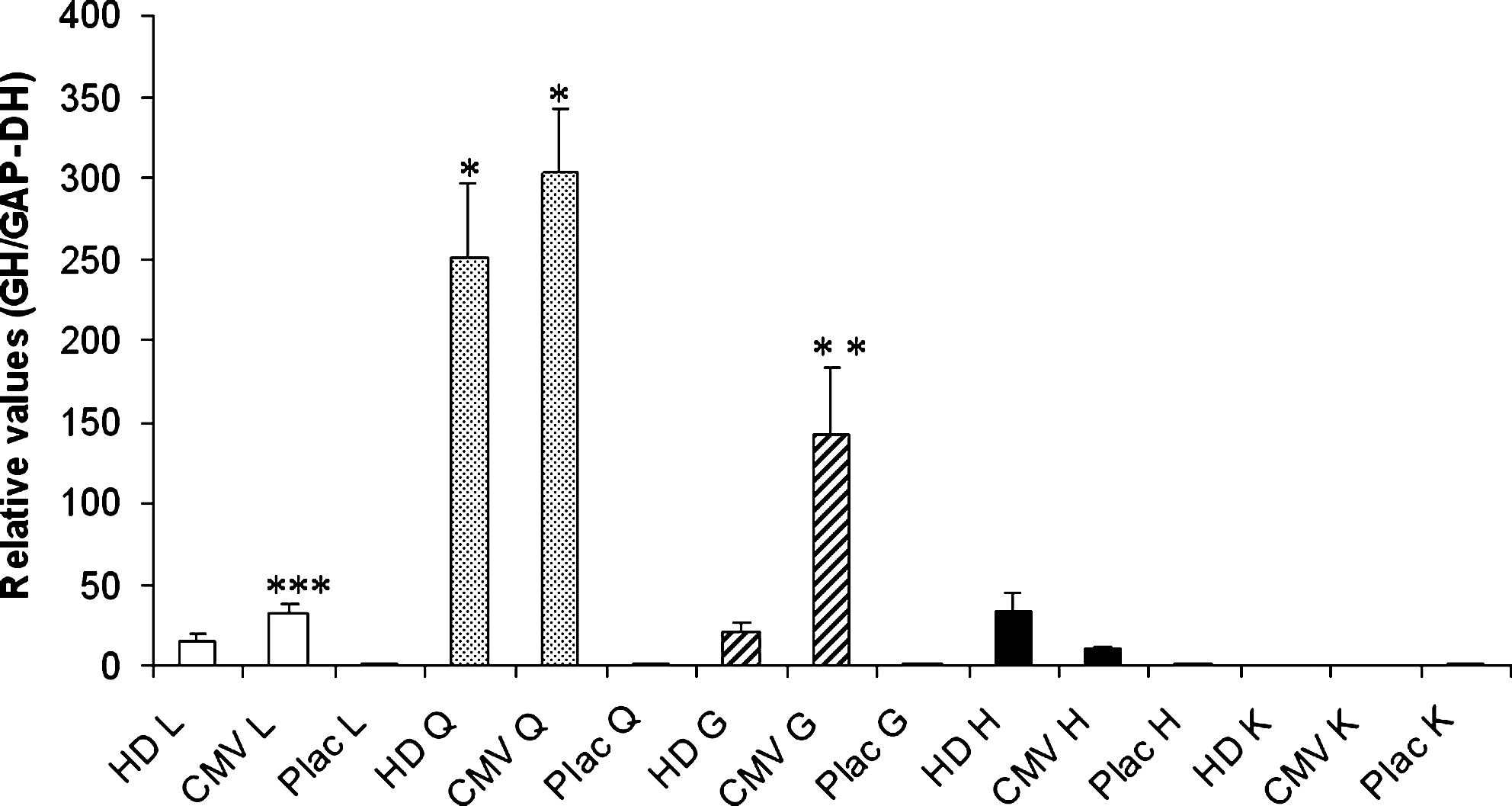
Growth hormone (GH) mRNA detected by quantitative RT-PCR (means ± SEM) in HD and CMV mice compared with Plac mice (liver, L; quadriceps, Q; gastrocnemius, G; heart, H; kidney, K). GH expression in HD Q tissue was significantly higher than in other HD tissues (*p < 0.01 vs. all HD tissues) and GH expression in CMV Q tissue was significantly higher than in other CMV tissues (*p < 0.05 vs. all CMV tissues). GH expression in CMV G tissue was higher than in HD and Plac G tissues (**p < 0.05). GH expression in CMV L tissue was higher than in Plac L tissue (***p < 0.01). GH expression in H tissue was not significantly different among the three groups.
Serum GH and IGF-I levels
At 6 weeks of age random serum GH levels in mice that had received either a low or high dose of dsAAV8-CK8-GH were not significantly higher than in Plac mice, whereas the GH levels in mice injected with dsAAV8-CMV-GH were higher than in all other groups. Levels of GH did not appear to be different among the HTZ, Plac, and GPF groups (Fig. 6A).

(
Serum IGF-I levels in LD, HD, and CMV mice were not significantly different from the Plac and GFP groups, and they were much lower compared with HTZ mice (Fig. 6B).
Histological analysis
Tissue sections from GHRHKO mice treated with dsAAV8-CK8-GFP showed that GFP was expressed in quadriceps, gastrocnemius, and heart muscles, whereas no detectable expression was observed in liver and kidney. No fluorescence was observed in tissues from the control group (Fig. 7). Because vectors were injected only into the right quadriceps, detection of GFP expression in the heart and gastrocnemius indicates vector leakage during or after the injection procedure.
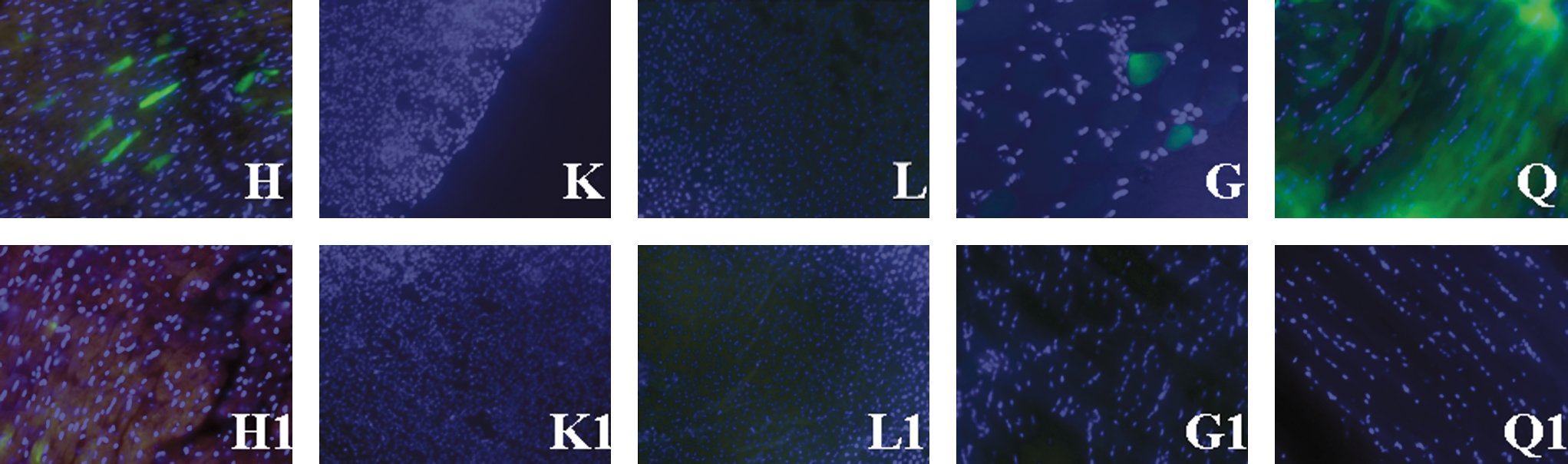
Fluorescence microscopy. At the end of the study dsAAV8-CK-GFP-injected GHRHKO mice showed fluorescence in heart (H), gastrocnemius (G), and quadriceps (Q) muscles. No fluorescence was detected in liver (L) and kidney (K) of GFP mice. No fluorescence was observed in tissues from Plac mice (heart, H1; kidney, K1; liver, L1; gastrocnemius, G1; quadriceps, Q1). Blue coloring is the result of DAPI nuclear staining. All photographs of the tissues were taken with a 2-sec exposure time under a fluorescence microscope. Color images available online at
Discussion
In this work we studied a dsAAV vector expressing GH under the control of a high-activity regulatory cassette derived from modified portions of the muscle creatine kinase (M-CK) promoter, specifically looking at delivery and muscle-specific expression of GH in GHD mice. We had used the same vector at similar dosage in our previous study, although in that study the virus had been administered intraperitoneally and GH was driven by a ubiquitous CMV promoter (Sagazio et al., 2008). We chose the intramuscular route of administration to increase the muscular targeting and to mimic a possible future therapeutic route, eliminating exposure of the liver to a high dose of virus. The inclusion of control HTZ mice provided the ability to compare treated and untreated GHRHKO mice with animals with a normal growth pattern on a similar genetic background (Alba and Salvatori, 2004).
Our data show that GHD mice treated with dsAAV-CK8-GH had a significant increase in length and body weight compared with untreated mice. GH expression promoted longitudinal growth and weight gain in mice that received both the low dose (LD) and high dose (HD) of vector, as they attained size phenotypes that were intermediate between those of placebo and HTZ mice.
The weight of most interior organs (heart, kidney, testis, and lung) of animals of each group did not differ significantly from each other, showing a good relationship with body size. The exception is represented by liver and spleen; the fact that placebo-treated animal have lower organ size compared with heterozygous mice suggests that in mice GH may have a specific effect on establishing the mass of an organ. Although GH caused normalization of spleen mass at both virus doses, liver—albeit heavier than in Plac animals—did not fully normalize, suggesting that liver growth requires higher GH levels than does spleen. The observation that liver growth requires GH fits nicely with our previous work in which GHD mice were treated with daily injections of recombinant mouse GH (Alba et al., 2005), as well as with the work of others (Khamaisi et al., 2007).
Interestingly, mice injected with the CMV-driven construct did not reach full growth normalization, presenting identical growth to that of LD and HD mice. This is in contrast with our previous study, in which the dsAAV-CMV-GH viral vector (at the same dosage, but administered intraperitoneally) was effective in fully normalizing growth (Sagazio et al., 2008). We hypothesize that the different administration route affected the bioavailability of the viral particles, as has been previously reported (Wang et al., 2005).
Bone length and body composition appeared to be affected by the treatment but to variable degrees. Whereas tibial and femoral lengths were normalized in all groups, apart from tibias in LD mice (Fig. 3), fat pools and lean mass did not fully normalize (Fig. 4). These findings suggest that serum GH reached levels that promoted growth without attaining full normalization, and possibly that tibias require higher levels of GH to reach optimal growth.
GH expression based on GH mRNA was detected in muscular tissues of HD and CMV mice, with the expression in quadriceps muscle (site of injection) significantly higher than in the gastrocnemius (Fig. 5). This finding supports the idea that the viral particles, injected in the quadriceps, infected the surrounding tissue to a great extent and led to mostly localized expression of GH. On the other hand, HD mice did not exhibit a significant increase in GH expression in nonmuscle tissues, whereas CMV mice showed expression in the liver. The CK8 regulatory cassette, rather than the route of administration, was therefore essential for muscle-specific GH expression, avoiding hepatic protein expression.
Quantitative RT-PCR results were confirmed by qualitative histological fluorescence studies performed on GFP-infected mice. As GFP was under the control of the same CK8 regulatory cassette, we expected its pattern of expression to be similar to GH expression. Indeed, fluorescence was more intense in quadriceps and heart muscle compared with gastrocnemius muscle, whereas it was undetectable in liver and kidney, confirming the muscle tissue-specific expression of GH via the CK8 regulatory cassette.
Interestingly, neither GH nor IGF-I serum levels were significantly increased in HD or LD mice, whereas serum GH levels were increased in mice injected with vectors containing the CMV-driven construct. As a consequence of lower levels of systemic GH and IGF-I, no abnormal increase in spleen mass—a trait often associated with recombinant GH administration (Vickers et al., 2002; Sagazio et al., 2008)—was observed in this study. It is noticeable that serum GH levels in HTZ mice were not higher than those in Plac or GFP mice, seemingly contradicting our previous findings (Alba and Salvatori, 2004). This may be because we measured nonstimulated GH levels, for which there may be overlap between normal and GHD animals.
Serum GH levels in the CMV-treated animals was higher than in HTZ animals, but circulating IGF-I was not increased. The discrepancy between serum GH and IGF-I levels is not surprising, as serum IGF-I levels are due mainly to liver production, and we have previously noticed differences between serum IGF-I levels and the effects of GH or GHRH on growth in GHRHKO mice (Alba et al., 2005). Accordingly, mice with liver-specific ablation of the IGF-I gene have essentially normal body growth despite extremely low serum IGF-I levels (Sjögren et al., 2000).
Several approaches to GH gene therapy have been developed, using different types of viral carriers (Hahn et al., 1996; Marmary et al., 1999) and different promoters (Rivera et al., 1999). We tested a striated muscle regulatory cassette for the selective expression of GH in skeletal muscles. Although GH production at the vector doses we used was not sufficient to fully normalize growth in GHD mice, vector-mediated expression of GH did promote whole body and bone longitudinal growth as well as influenced body composition. Moreover, GH expression was detected preferentially in skeletal muscle, confirming the use of CK-based regulatory cassettes as a possible tool in these types of therapeutic applications. The heart expression in HD mice could represent a possible pitfall of this approach, as persistent high levels of GH in the heart may have damaging effects on heart structure. However, careful administration of therapeutic vectors in the much larger skeletal muscles of humans might greatly decrease the possibility of vector leakage and subsequent uptake by heart muscle cells. Interestingly, the serum GH levels detected in the current studies were much lower than previously observed after intraperitoneal injection of a double-stranded AAV vector expressing GH via the CMV promoter (Sagazio et al., 2008), and significantly lower than was detected after the intramuscular injection of a CMV-driven GH-expressing virus. This is a promising result, as it is likely to reduce the risks of GH overexpression. In conclusion, although the strategy of vector-mediated GH therapy is still far from a clinical application, the present study is a promising step toward future systemic delivery of GH using a gene therapy approach that limits GH expression to skeletal muscles.
Footnotes
Acknowledgments
Supported in part by grant 1R21 DK073175 (to R.S.), and by NIH grants 2R01 AR18860 and 1P01 NS046788, and by the Muscular Dystrophy Association (to S.D.H.). The authors thank Dr. Mehboob Hussain (Division of Metabolism, Department of Pediatrics, Johns Hopkins University) for help with immunofluorescence studies.
Authors Disclosure Statement
No competing financial interest exists.