
Abstract
Interleukin (IL)-24 is an excellent therapeutic gene for cancer therapy. In this work, IL-24 was inserted into Ad.sp-E1A(Δ24), an oncolytic adenovirus with a 24-bp deletion in the E1A gene, which was driven by the survivin promoter to form Ad.sp-E1A(Δ24)-IL-24. Ad.sp-E1A(Δ24)-IL-24 has an excellent antitumor effect in vitro for human nasopharyngeal, liver, lung, and cervical carcinoma cell lines but does no or little damage to normal cell lines L-02 and WI38. Furthermore, it achieved nearly complete inhibition (although not elimination) of NCI-H460 lung carcinoma growth in nude mice. The antitumor efficacy of Ad.sp-E1A(Δ24)-IL-24 on NCI-H460 cells was clearly mediated by apoptosis, because it induced caspase-3 and poly(ADP-ribose) polymerase cleavage. This is the first report of Ad.sp-E1A(Δ24)-IL-24 with such an excellent, broad, and specific antitumor effect in vitro and nearly complete inhibition of lung tumor growth in vivo.
Introduction
Interleukin (IL)-24 shows a strong antitumor effect, inhibiting tumor in three ways: (1) by induction of tumor apoptosis and its bystander effect (Su et al., 2005; Sauane et al., 2006, 2008); (2) by inhibition of angiogenesis (Ramesh et al., 2003; Wang et al., 2007; Xie et al., 2008); and (3) by an increase in immunological effect (Miyahara et al., 2006; Commins et al., 2008). The first oncolytic virus used in our laboratory was ZD55. By insertion of the IL-24 gene into ZD55, ZD55-IL-24 was formed. The antitumor effect of ZD55-IL-24 was 100-fold higher than that of the respective gene therapy product Ad-IL-24 in vitro (Zhao et al., 2005; Inoue et al., 2006), which is now in a phase III clinical trial in the United States. Therefore an application for an international patent for ZD55-IL-24 has been made according to the terms of the Patent Cooperation Treaty (PCT). By the combined use of two ZD55 genes, such as ZD55-IL-24 plus ZD55-TRALL, xenografts of colorectal carcinoma, hepatoma, and gastric cancer in nude mice could all be completely eliminated (Zhao et al., 2006).
Survivin is a unique member of the inhibitor of apoptosis (IAP) protein family, which plays a key role in the regulation of apoptosis and cell division (Ambrosini et al., 1997; Adida et al., 1998; Altieri, 2008). Most significantly, survivin is expressed in almost all human tumors but is rarely detectable in normal cells and tissues. Overexpression of survivin has been implicated in lung cancer and other cancers (Blanc-Brude et al., 2003; Falleni et al., 2003; Lu et al., 2004; Karczmarek-Borowska et al., 2005; Warnecke-Eberz et al., 2008). Survivin can be used as an indicator of tumor staging and prognosis. Studies have showed that survivin promoter-driven oncolytic adenovirus exhibited tumor-selective cytotoxicity in vitro and in vivo (Kamizono et al., 2005; Zhu et al., 2006; Wang, 2008), which suggested the survivin promoter on transcriptional targeting is a good candidate for targeting cancer therapy. In addition, oncolytic virus Ad5-E1A(Δ24), with a 24-bp deletion in the E1A region responsible for binding retinoblastoma (Rb) protein and its replication, is restricted to dividing cells or to Rb-inactive arrested cells and exhibits tumor-selective capability (Fueyo et al., 2000). The antitumor activity of the therapeutic gene should safely progress along with the improved tumor-selective adenoviral vector (Lee et al., 1999; Shinoura and Hamada, 2003). When the IL-24 gene was inserted into Ad.sp-E1A(Δ24), Ad.sp-E1A(Δ24)-IL-24 was formed. This is the first report of a double-regulated oncolytic adenovirus (Ad.sp-E1A(Δ24)-IL-24) constructed with a survivin promoter-controlled E1A gene with a 24-bp deletion (Δ24) targeting the Rb pathway to improve safety. Our data show that Ad.sp-E1A(Δ24)-IL-24 can achieve wide-range tumoral targeting to induce dramatic cytotoxicity in tumor cells in vitro. Furthermore, Ad.sp-E1A(Δ24)-IL-24 achieved essentially complete inhibition (although not elimination) of xenografted lung cancer in nude mice.
Materials and Methods
Cell lines and culture conditions
HEK293 (a human embryonic kidney cell line containing the E1A region of adenovirus serotype 5 [Ad5]) was obtained from Microbix Biosystems (Toronto, ON, Canada). All tumor and normal cell lines were from the American Type Culture Collection (ATCC, Manassas, VA) or the Shanghai Cell Collection (Shanghai, China); these included CNE (human nasopharyngeal carcinoma), NCI-H460 (human lung carcinoma), NCI-H1299 (human lung carcinoma), PLC (human hepatocarcinoma), HeLa (human cervical carcinoma), L-02 (human normal liver), and WI38 (human normal fetal lung fibroblast).
CNE, NCI-H460, NCI-H1299, PLC, HeLa, L-02, and HEK293 cell lines were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 4 mM glutamine, penicillin (50 U/ml), and streptomycin (50 mg/ml). The WI38 cell line was cultured in DMEM supplemented with 20% FBS. All the cell lines were cultured at 37°C in 5% CO2.
Construction of plasmids, and generation and purification of adenoviral vectors
Plasmid pCN103 carrying the adenovirus serotype 5 E1A gene with a 24-bp deletion from bp 922 to bp 946, was kindly provided by C. Qian of our institute and subcloned into pXC2 to form pXC2-E1A(Δ24). The human survivin promoter (hSurP) was further subcloned into plasmid pXC2-E1A(Δ24) to form the plasmid pAd.sp-E1A(Δ24). After inserting IL-24 expression cassette into it, pAd.sp-E1A(Δ24)-IL-24 was obtained. The oncolytic viruses Ad.sp-E1A(Δ24) and Ad.sp-E1A(Δ24)-IL-24 were generated by homologous recombination between pAd.sp-E1A(Δ24) or pAd.sp-E1A(Δ24)-IL-24 with the adenovirus packaging plasmid pBHGE3 (Microbix Biosystems) in HEK293 cells. Each recombinant adenovirus was isolated through three rounds of plaque purification in HEK293 cells and purified by ultracentrifugation in a cesium chloride gradient. Moreover, viral titers were determined by TCID50 (median tissue culture infective dose) assay in HEK293 cells. Cells were infected with adenovirus at various doses at 37°C in a humidified atmosphere containing 5% CO2.
Adenoviral progeny assay
To determine viral progeny, tumor cells and normal cells were infected with Ad.sp-E1A(Δ24) and Ad.sp-E1A(Δ24)-IL-24 or with wild-type adenovirus (Wt-Ad) at a multiplicity of infection (MOI) of 10. After 5 hr, medium was removed and cells were washed three times with phosphate-buffered saline (PBS), and then 2 ml of fresh medium was added. Two days postinfection, cells were collected and virus was released by three freeze–thawing cycles and centrifuged to collect the supernatant. Virus production was determined by TCID50 assay in HEK293 cells.
Cell viability assay
Cells were plated in 96-well plates and treated with various adenoviruses. After infection for 4 days at the indicated MOIs, the cell survival rate was evaluated by a standard 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Sigma, St. Louis, MO). Medium was removed and fresh medium containing MTT (0.5 mg/ml) was added to each well. The cells were incubated at 37°C for 4 hr; the supernatant of each well was drawn off carefully and then an equal volume (150 μl) of dimethyl sulfoxide (DMSO) was added to each well and mixed thoroughly on a concentrating table for 10 min. The absorbance from the plates was read at 595 nm with a DNA microplate reader (GENios model; Tecan, Maennedorf, Switzerland).
Cytopathic effect assay
PLC, NCI-H1299, and NCI-H460 tumor cell lines, as well as the L-02 and WI38 normal cell lines, were grown to subconfluence and infected with adenoviruses at various MOIs. Four days after infection, cells were exposed to 2% crystal violet in 20% methanol for 15 min and then washed with distilled water and documented by photography.
Western blot analysis
To determine the expression of various proteins, Western blot analysis was performed as described in a standard protocol (Zhao et al., 2006). Cells were harvested by trypsinization and resuspended in lysis buffer (62.5 mM Tris-HCl [pH 6.8], 2% sodium dodecyl sulfate [SDS], 10 mM glycerol, 1.55% dithiothreitol). The total protein concentration was determined with a bicinchoninic acid (BCA) protein assay kit (Pierce Biotechnology, Rockford, IL) as described in the manufacturer's protocol. Protein samples were then separated by SDS–12% polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes (Pierce Biotechnology). Membranes were blocked in 5% bovine serum albumin (BSA) solution and incubated with primary antibodies, which were then detected with the appropriate secondary fluorescent antibodies and checked with an Odyssey infrared imaging system (LI-COR Biosciences, Lincoln, NE). The primary antibodies used were mouse monoclonal anti-mda-7/IL-24 (GenHunter, Nashville, TN), rabbit polyclonal anti-caspase-3 (Cell Signaling Technology, Danvers, MA), rabbit monoclonal (Epitomics, Burlingame, CA), anti-poly(ADP-ribose) polymerase (PARP) (Santa Cruz Biotechnology, Santa Cruz, CA) anti-E1A (Abcam, Cambridge, UK), and anti-actin (Beyotime, Haimen, China).
Flow cytometric analysis
Cells infected with adenovirus were trypsinized and washed once with complete medium. Aliquots of cells (5 × 105) were resuspended in 500 μl of binding buffer and stained with fluorescein isothiocyanate (FITC)-labeled annexin V (BioVision, Palo Alto, CA) according to the manufacturer's instructions. A fluorescence-activated cell-sorting (FACS; BD Biosciences, San Jose, CA) assay was performed immediately after staining.
Hoechst 33258 staining
NCI-H460 cells were seeded in 6-well culture plates with slides and infected with Ad.sp-E1A(Δ24) or Ad.sp-E1A(Δ24)-IL-24 at an MOI of 5; uninfected cells served as control. After 48 hr, cells were treated with the apoptosis-Hoechst 33258 staining kit (Beyotime) for 5–10 min as described in the manufacturer's protocol, washed with PBS twice, and observed under a fluorescence microscope.
Studies of xenograft tumors in nude mice
All animals used in these experiments were maintained in the institutional facilities in accordance with regulations and standards of the U.S. Department of Agriculture and the National Institutes of Health. Female BALB/c nude mice at 4–5 weeks if age, obtained from the Animal Research Committee of the Institute of Biochemistry and Cell Biology (Shanghai, China), were used in all the experiments. NCI-H460 cells (1 × 107) were injected subcutaneously into the lower right flank of female nude mice. After 6 days, the tumor xenograft model was established. Each group was composed of at least eight animals and tumor growth was monitored and measured weekly with a Vernier caliper. Tumor volume (V) was calculated according to the formula V (mm3) = 1/2 × length (mm) × width (mm)2. When the tumors were 100–130 mm3 in size, mice were randomized into three groups and a daily dose of 5 × 108 plaque-forming units (PFU) of Ad.sp-E1A(Δ24) or Ad.sp-E1A(Δ24)-IL-24 suspended in 100 μl of PBS, or 100 μl of PBS alone, was administrated intratumorally once every other day for a total of four times. Tumors were harvested on day 4 posttreatment for histopathological and transmission electron microscopy (TEM) analysis.
Immunohistochemical study
For immunohistochemical (IHC) analysis, tumors on day 4 posttreatment were harvested and fixed in 4% paraformaldehyde, embedded in paraffin, and cut into 4-mm sections. These sections were stained with mouse monoclonal anti-IL-24 and anti-adenoviral E1A antibodies at 1:300 and 1:50 dilutions, respectively. The slides were then washed with PBS and incubated with the avidin–biotin–peroxidase complex reagent (Vector Laboratories, Burlingame, CA) and detected with diaminobenzidine tetrahydrochloride (Bendrik et al., 2008) solution containing 0.006% hydrogen peroxide. Hematoxylin was used as a counterstain. Tissue sections stained without primary antibodies were used as negative controls.
Transmission electron microscopy analysis
For electron microscopy, tumor samples (1 mm3) were fixed in a phosphate-buffered mixture of 2.5% glutaraldehyde overnight, followed by 1 hr of fixation with 1% osmium tetroxide. The tissues were rinsed in water, dehydrated through a graded series of ethanol and propylene oxide, and embedded in Epon 812 resin (Shell Chemicals, Houston, TX). After examination of semithin sections, areas were selected and subjected to ultrathin sectioning. Sections collected on 200-mesh copper grids were contrasted with lead citrate and uranyl acetate, examined, and photographed with a JEOL 100CX transmission electron microscope (JEOL, Akishima, Japan).
Statistical analysis
The statistical significance of experimental results was calculated by analysis of variance (ANOVA) and Student t test. Data were considered statistically significant at p < 0.05.
Results
Characterization of oncolytic adenovirus Ad.sp-E1A(Δ24)-IL-24 and its selective replication in tumor cells
Adenoviral E1A is the early gene for viral replication in host cells; after deletion of 24 bp, Ad-E1A(Δ24) was formed. The survivin promoter is a promising candidate for the construction of a tumor-specific oncolytic adenovirus. The dual-regulated oncolytic adenovirus Ad.sp-E1A(Δ24)-IL-24 is also driven by the survivin promoter and harbors the antitumor IL-24 gene (Fig. 1A).
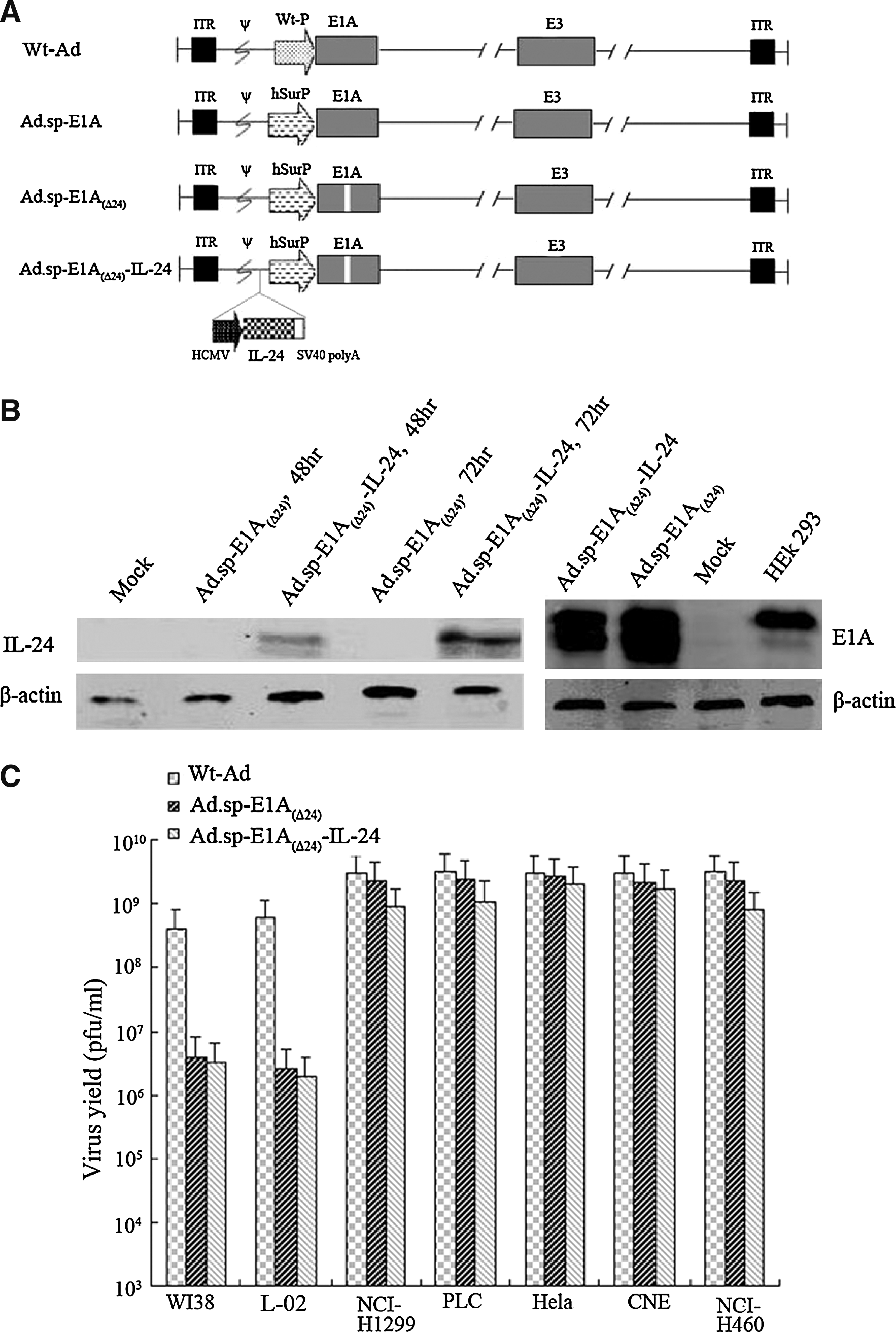
Characterization of oncolytic adenovirus Ad.sp-E1A(Δ24)-IL-24 and its selective replication in tumor cells. (
All the constructs were characterized by Western blot assay. In Fig. 1B, NCI-H460 cells infected with Ad.sp-E1A(Δ24) or Ad.sp-E1A(Δ24)-IL-24, or positive HEK293 cells, expressed E1A protein, whereas mock-infected cells failed to express E1A protein. To examine IL-24 gene expression, NCI-H460 cells were infected with Ad.sp-E1A(Δ24) or Ad.sp-E1A(Δ24)-IL-24 at an MOI of 5 for 48 or 72 hr. Cells were then harvested and subjected to Western blot assay. Transduction of Ad.sp-E1A(Δ24)-IL-24 resulted in obvious IL-24 expression compared with no expression in control or Ad.sp-E1A(Δ24)-transduced cells. Moreover, the IL-24 expression level of cells infected for 72 hr with Ad.sp-E1A(Δ24)-IL-24 was higher than that of cells infected for 48 hr, which indicated that the IL-24 expression level gradually improved with the replication of virus in lung cancer NCI-H460 cells.
To examine whether the transgene could interfere with the selective replicative ability of recombinant adenoviruses in tumor cell lines, a progeny assay was performed in tumor cells (NCI-H1299, PLC, HeLa, CNE, and NCI-H460) and normal cells (L-02 and WI38) infected with Ad.sp-E1A(Δ24), Ad.sp-E1A(Δ24)-IL-24, or Wt-Ad at an MOI of 10. As shown in Fig. 1C, Ad.sp-E1A(Δ24) and Ad.sp-E1A(Δ24)-IL-24 replicated in infected tumor cells to similar levels, which were comparable to that of Wt-Ad. In contrast, the replicative capacity of these viruses was much reduced in normal cells. These data indicated that expression of IL-24 gene did not affect the selective replicative ability of oncolytic adenoviruses.
Cytotoxicity of Ad.sp-E1A(Δ24)-IL-24 and Ad.sp-E1A(Δ24) in tumor cells but not in normal cells
To evaluate the cytotoxicity of recombinant adenovirus, tumor cell lines (NCI-H1299, HeLa, CNE, NCI-H460, and PLC) and normal cell lines (L-02 and WI38) were infected with Ad.sp-E1A(Δ24) or Ad.sp-E1A(Δ24)-IL-24 at MOIs of 0.1, 1, 5, and 10. After 4 days, cytotoxicity was determined by MTT assay. As shown in Fig. 2A, Ad.sp-E1A(Δ24)-IL-24 and the control virus Ad.sp-E1A(Δ24) could induce dose-dependent cytotoxicity in all five tumor cell lines. These showed the broad antitumor effect of Ad.sp-E1A(Δ24)-IL-24. In addition, Ad.sp-E1A(Δ24)-IL-24 was higher in cytotoxicity in tumor cells (PLC, HeLa, NCI-H1299, and NCI-H460) than was Ad.sp-E1A(Δ24) at each infected concentration gradient (MOIs of 0.1, 1, 5, and 10). Moreover, because of the selective replicative ability of the oncolytic adenoviruses Ad.sp-E1A(Δ24) and Ad.sp-E1A(Δ24)-IL-24, there was no obvious cytotoxicity to the two normal cell lines (Fig. 2B). The results indicated that our dual-regulated oncolytic virus could lead to an obvious and broad tumor-specific cytotoxic effect in vitro, and had no or little effect on normal cells.
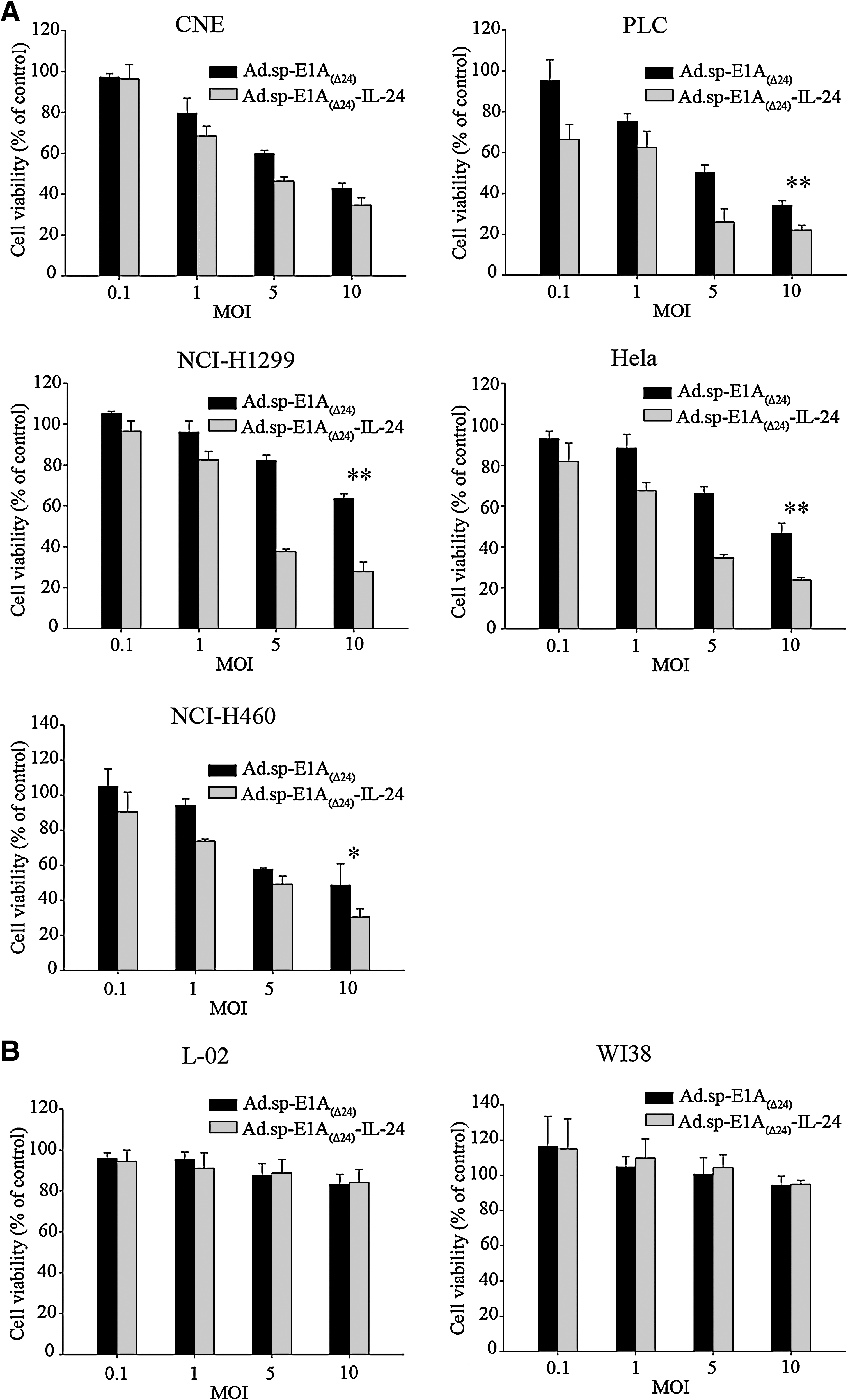
Cytotoxicity of oncolytic adenovirus in tumor cells in vitro. (
Comparison of antitumor activity of gene therapy and targeting gene virotherapy in vitro
To compare the cytopathic capacity of Ad-IL-24 and Ad.sp-E1A(Δ24)-IL-24, a human liver cancer cell line (PLC), lung cancer cell lines (NCI-H1299 and NCI-H460), a normal human lung fibroblast cell line (WI38), and a human liver cell line (L-02) were infected with Ad-IL-24, Ad.sp-E1A(Δ24), or Ad.sp-E1A(Δ24)-IL-24 at MOIs of 0, 0.1, 5, 10, and 100. Cells were stained with crystal violet 4 days later. Significant cytopathic effect was observed in all tumor cell lines infected with Ad.sp-E1A(Δ24)-IL-24, as compared with cells infected with Ad.sp-E1A(Δ24) or Ad-IL-24 (Fig. 3). Moreover, the cytopathic effect of Ad.sp-E1A(Δ24)-IL-24 was superior to that of Ad.sp-E1A(Δ24) and was about 100 times greater than that of Ad-IL-24 in the NCI-H1299 and NCI-H460 cell lines. For all three viruses, no obvious cytopathic effect was detected in the two normal cell lines (WI38 and L-02) at the same MOIs compared with the treated tumor cell groups, although significant cytotoxicity was observed in WI38 and L-02 cells at the highest MOI of 100. These results suggest that Ad.sp-E1A(Δ24)-IL-24 can selectively replicate in various tumor cells and inhibit the growth of tumor cells more effectively than Ad.sp-E1A(Δ24) (oncolytic virotherapy) and Ad-IL-24 (gene therapy).

Tumor-selective cytopathic effect of Ad.sp-E1A(Δ24)-IL-24 and Ad-IL-24. Tumor cells (PLC, NCI-H1299, and NCI-H460) and normal cells (L-02 and WI38) were seeded in 24-well plates at a density of 5 × 104 cells for each well and infected with Ad.sp-E1A(Δ24), Ad.sp-E1A(Δ24)-IL-24, or Ad-IL-24 at the indicated MOIs. Seven days later, cells were stained with crystal violet. Color images available online at
To further evaluate the kinetics of cytotoxicity induced by Ad.sp-E1A(Δ24)-IL-24, tumor cells (PLC, NCI-H1299, and NCI-H460) and normal cells (WI38 and L-02) were plated in 96-well plates and infected with the previously mentioned three viruses. As shown in Fig. 4, the cytotoxic effect of Ad.sp-E1A(Δ24)-IL-24 on the cancer cell lines was much more obvious than that of Ad-IL-24 or Ad.sp-E1A(Δ24); this was particularly evident with the longer treatment times. There was little or no cytopathic effect of Ad.sp-E1A(Δ24)-IL-24, Ad-IL-24, and Ad.sp-E1A(Δ24) on normal cells. Similar results were obtained with other tumor cell lines and normal cell lines (data not shown).
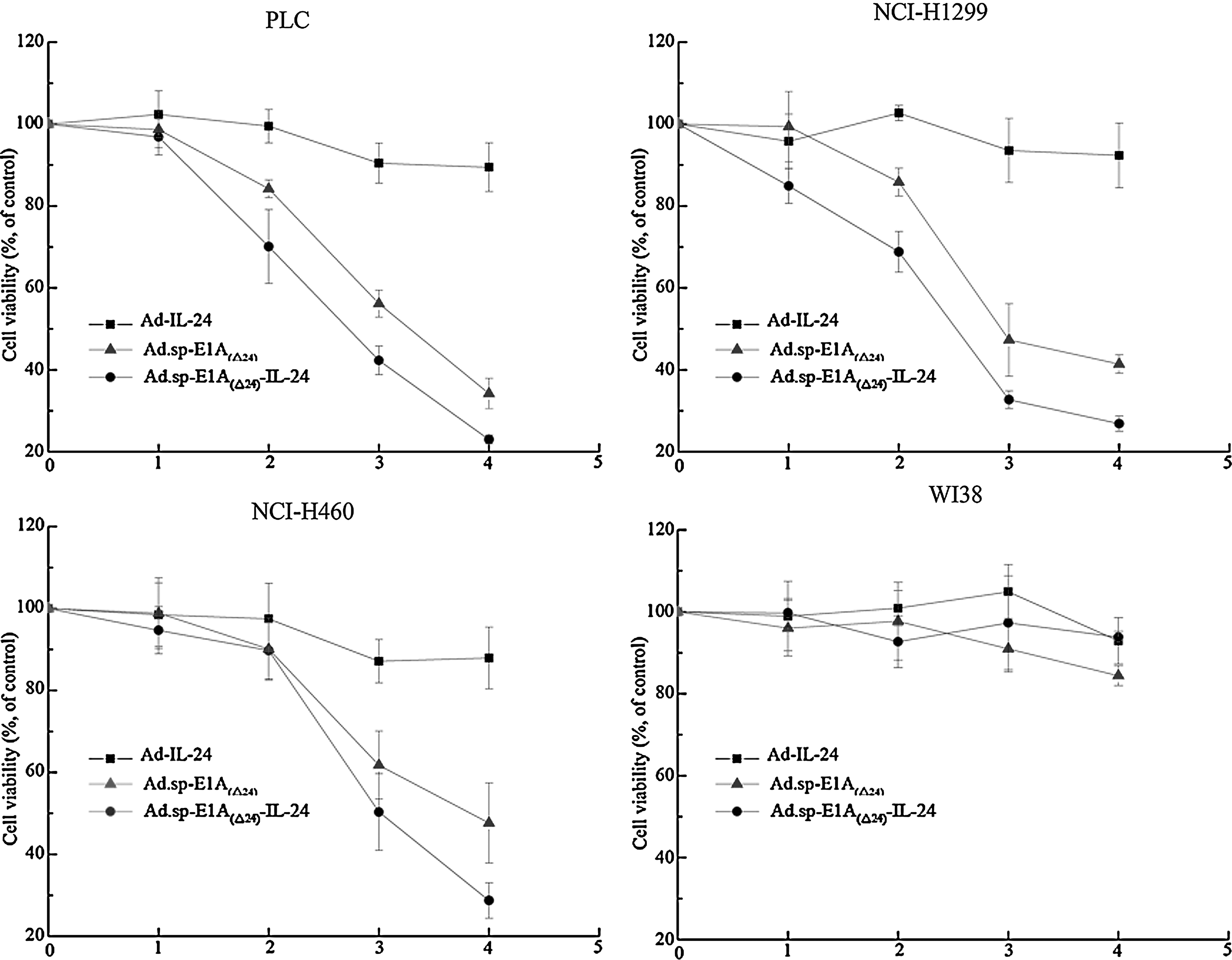
Cell viability was assayed by MTT test. Tumor cells (PLC, NCI-H1299, and NCI-H460) and normal cells (L-02 and WI38) were infected with Ad-IL-24, Ad.sp-E1A(Δ24), or Ad.sp-E1A(Δ24)-IL-24 at an MOI of 10. On days 1, 2, 3, and 4 postinfection, cells were stained with MTT as described in Materials and Methods. Results are presented as means ± SD (error bars) of triplicate experiments and are expressed as a percentage of uninfected control cells.

Mechanism of apoptosis is mediated by Ad.sp-E1A(Δ24)-IL-24 in NCI-H460 cells
We investigated whether Ad.sp-E1A(Δ24)-IL-24 kills tumor cells through apoptosis. Hoechst 33258 staining for apoptotic cells clearly showed that Ad.sp-E1A(Δ24)-IL-24 and Ad.sp-E1A(Δ24) led to remarkable apoptotic morphological changes such as chromatin condensation, nuclear fragmentation, and apoptotic bodies in the NCI-H460 cell line (Fig. 5A). Moreover, to further confirm induction of apoptosis, FACS analysis was performed to test apoptosis induction by Ad.sp-E1A(Δ24) and Ad.sp-E1A(Δ24)-IL-24 in the lung cancer cell line NCI-H460 and the normal cell line WI38. Cells uninfected with adenovirus served as a negative control. The apoptotic effect of Ad.sp-E1A(Δ24)-IL-24 at an MOI of 10 was more significant than that of any other group, even Ad.sp-E1A(Δ24) (Fig. 5B). Although Ad.sp-E1A(Δ24)-IL-24 and Ad.sp-E1A(Δ24) led to similar apoptotic rates at an MOI of 5, a dramatic statistical discrepancy appeared in the induced apoptosis rates of Ad.sp-E1A(Δ24)-IL-24 compared with the mock group (p < 0.01). No apoptosis was detected in normal cells infected with either Ad.sp-E1A(Δ24) or Ad.sp-E1A(Δ24)-IL-24 alone. These results suggest that the induction of apoptosis in tumor cells was due to replication of the oncolytic virus and the expression of IL-24, whereas the few apoptotic effects in normal cells indicated that both Ad.sp-E1A(Δ24) and Ad.sp-E1A(Δ24)-IL-24 are safe. Furthermore, activation of the caspase pathway was examined to explore the mechanism of tumor cell apoptosis. As shown in Fig. 5C, the cleavage of caspase-3 and poly(ADP-ribose) polymerase (PARP) was obviously increased in NCI-H460 cells treated with Ad.sp-E1A(Δ24)-IL-24 compared with Ad.sp-E1A(Δ24) or the control, which indicates the activated process of apoptosis.
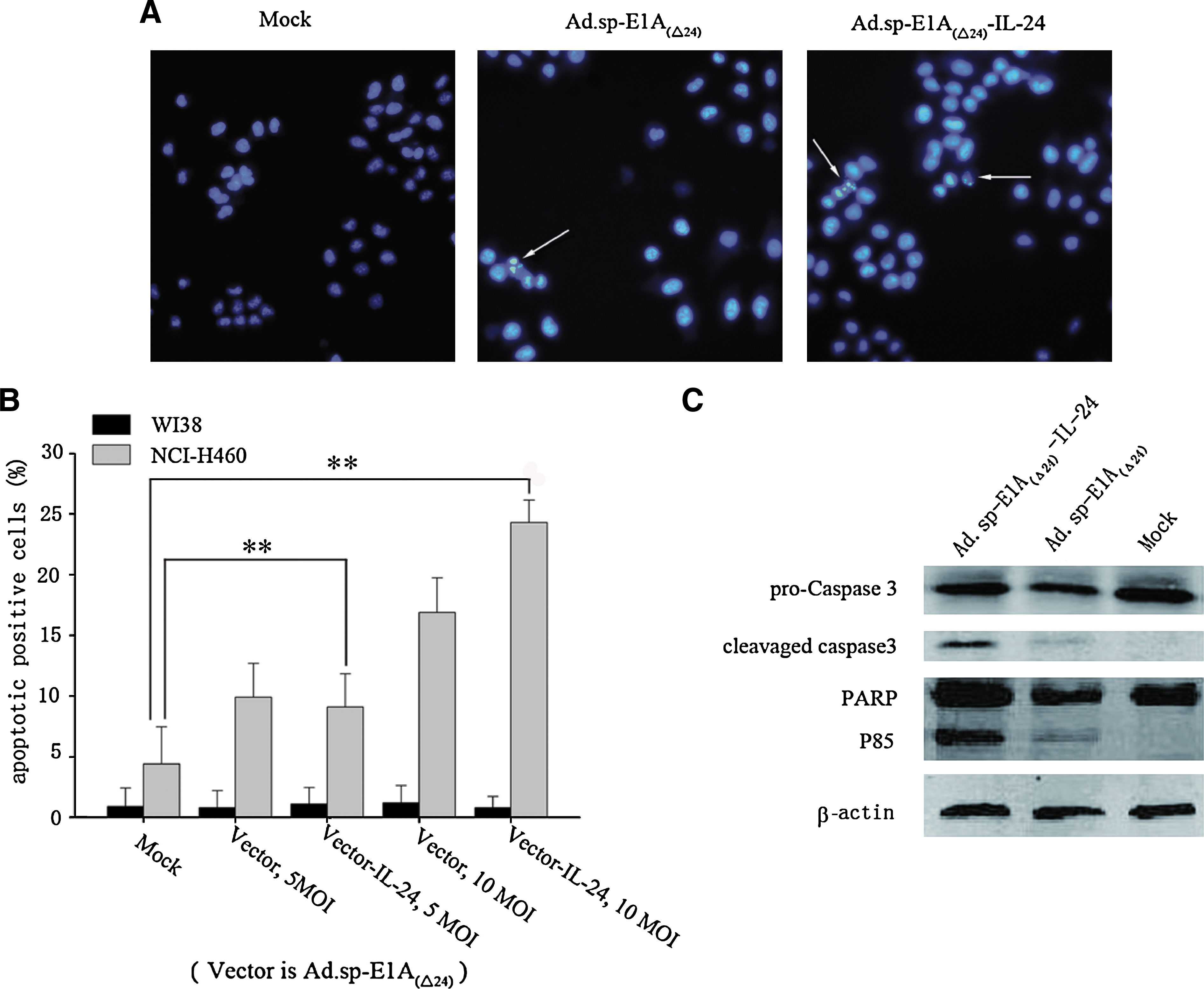
Apoptosis detection in tumor cells and activation of caspase-3 mediated by Ad.sp-E1A(Δ24)-IL-24. (
Potent antitumor efficacy of Ad.sp-E1A(Δ24)-IL-24 in nude mice
An NCI-H460 human lung tumor xenograft model was established in nude mice. When the tumors grew to 100–130 mm3, PBS, Ad.sp-E1A(Δ24), or Ad.sp-E1A(Δ24)-IL-24 (5 × 108 PFU/dose) in 100 μl WAS injected intratumorally every other day for a total of four times. Animals treated with Ad.sp-E1A(Δ24)-IL-24 exhibited significant suppression of tumor growth compared with control PBS-treated animals (p < 0.005) and Ad.sp-E1A(Δ24)-treated animals (p < 0.005). The average tumor volume on injection with Ad.sp-E1A(Δ24)-IL-24 was 136.4 mm3 at 43 days, which continued to decrease to below 95 mm3, even lower than the original volume of 100 mm3; this indicates a complete inhibition of NCI-H460 human lung tumor growth, whereas the tumor volume of PBS-treated or Ad.sp-E1A(Δ24)-treated mice was 2467.3 and 545.8 mm3, respectively. In addition, the Ad.sp-E1A(Δ24)-treated group also showed notable antitumor efficacy as compared with the PBS-treated group (p < 0.05) (Fig. 6A).

Antitumor efficacy of Ad.sp-E1A(Δ24)-IL-24 in nude mice. Female BALB/c nude mice were subcutaneously inoculated with NCI-H460 cells (1 × 107). When tumors reached a size of 100–130 mm3, the animals were treated with an intratumoral injection of Ad.sp-E1A(Δ24) or Ad.sp-E1A(Δ24)-IL-24, or PBS. (
Survival of animals was also monitored by the Kaplan–Meier method. Animals treated with Ad.sp-E1A(Δ24)-IL-24 all survived (eight of eight), whereas in the other groups zero of eight survived (PBS) and four of eight survived [Ad.sp-E1A(Δ24)] (Fig. 6B). In addition, the volume of tumors treated with Ad.sp-E1A(Δ24)-IL-24 sequentially regressed until day 72, at which time the tumor volume was 95 mm3, less than the volume of about 100 mm3 before treatment.
Morphologic evidence of Ad.sp-E1A(Δ24)-IL-24 effect on tumor growth inhibition
Hematoxylin and eosin (H&E) staining of subcutaneous tumor sections demonstrated that tumor injections of Ad.sp-E1A(Δ24)-IL-24 caused profound cell death and symptoms of necrosis in the tumor mass. Vessel growth was significantly suppressed by treatment with Ad.sp-E1A(Δ24)-IL-24, showing its antiangiogenesis effect (Fig. 7A). IHC analysis also showed E1A protein mediated by oncolytic adenovirus Ad.sp-E1A(Δ24) and Ad.sp-E1A(Δ24)-IL-24 in lung tumor tissues, but not with PBS treatment (Fig. 7B). In addition, there was local high expression of IL-24 in tumor cells infected with Ad.sp-E1A(Δ24)-IL-24 (Fig. 7C).

Both pathological detection by H&E and IHC staining assay and morphological observation by TEM analysis were performed. Subcutaneous NCI-H460 tumors receiving various treatments were harvested 4 days after infection with viruses and tumor sections were treated as described in Materials and Methods. (
Morphologic observation by TEM analysis showed the viral replication of Ad.sp-E1A(Δ24) and Ad.sp-E1A(Δ24)-IL-24 (Fig. 7D). Mature virus and submature adenoviral particles can be seen (marked as M and S, respectively, in Fig. 7D). The significant apoptosis in tumor tissues injected with Ad.sp-E1A(Δ24)-IL-24 or Ad.sp-E1A(Δ24) was detected as the classic characteristics of apoptosis, including nuclear collapse and an increased nuclear-to-cytoplasmic ratio; also, the appearance of nucleus deformation and heterochromatin, chromatin condensed in lumps at the inner side of the nuclear envelope were observed (Fig. 7E).
The preceding results suggested that the inhibitory effect on tumor growth was due to IL-24-induced apoptosis, antiangiogenesis, and viral propagation.
Discussion
Tumor-selective oncolytic virus has been used as a vector for gene delivery to improve cancer therapy. The E1A gene is an essential gene to be transcribed for efficient adenoviral replication in host cells. Replacing the native E1A promoter with a tumor-specific promoter, such as the prostate-specific antigen promoter (Rodriguez et al., 1997), the α-fetoprotein promoter (Hallenbeck et al., 1999), the midkine promoter (Adachi et al., 2001), the tyrosinase promoter (Nettelbeck et al., 2002), the pancreas promoter CCKAR (Xie et al., 2007), and so on, revealed that it was an efficient strategy by which to improve the targeting of adenovirus to tumor.
Survivin, one of the inhibitor of apoptosis (IAP) proteins, functions as a key regulator of mitosis and programmed cell death. Survivin was described as an inhibitor of caspase-9 (Mita et al., 2008). However, research studies have shown that the role of survivin in cancer pathogenesis is not limited to apoptosis inhibition but also involves regulation of the mitotic spindle (Li et al., 1998; Petersen and Hagan, 2003; Beltrami et al., 2004) and the inhibition of angiogenesis (Nassar et al., 2008) and chemoresistance (Virrey et al., 2008). Survivin gene expression is transcriptionally repressed by wild-type p53 and can be deregulated in cancer by several mechanisms, including gene amplification, increased promoter activity, and loss of p53 function (Raj et al., 2008; Yang et al., 2008). Studies revealed that the survivin promoter is superior even to the telomerase reverse transcriptase (TERT) promoter at both the responsive promoter activity and as a cancer-specific biomarker in the quantities of different kinds of tumors (Kamizono et al., 2005). Another targeting is the deletion of 24 bp in E1A conservative region 2 (CR2), which make adenovirus available to target the Rb dysfunctional signal pathways in many cancers (Fueyo et al., 2000). Thus, the 1100-bp human survivin promoter was chosen as the cancer-specific promoter to drive the 24-bp-deleted E1A gene to form the double-regulated adenoviral vector Ad.sp-E1A(Δ24) in this study, although a shorter survivin promoter (Li and Altieri, 1999a,b) has also been used. The results showed that vector Ad.sp-E1A(Δ24) can replicate and induce cytotoxicity in various tumors but rarely in normal cells, as shown in Figs. 2, 3, and 4.
IL-24 is a cytokine belonging to the IL-10 family. It is a cancer-specific apoptosis-inducing gene with a broad antitumor spectrum, inducing apoptosis in a variety of tumor types including melanomas, gliomas, and cancers of the breast, colon, lung, cervix, pancreas, and prostate (Gupta et al., 2006; Fisher et al., 2007). Ad-IL-24 has been employed to treat ovarian cancer cells and has led to tumor growth suppression in vivo (Gopalan et al., 2007); and INGN 241 (Gopalkrishnan, 2002; Cunningham et al., 2005; Tong et al., 2005; Eager et al., 2008) is in a phase III clinical trial. The mechanism of IL-24 action is to induce apoptosis (Zhao et al., 2005, 2006). Ad.sp-E1A(Δ24)-IL-24 can induce apoptosis by activated caspase-3 and the cleavage of PARP in tumor cells but rarely damages normal cells (Fig. 5), which was also proved by Lebedeva and colleagues (Lebedeva et al., 2002). In addition, IL-24 can induce a direct antitumor effect through its “bystander antitumor effect” and resistance to antiapoptotic proteins Bcl-2 and Bcl-xL (Su et al., 2005). Ad-IL-24 has antiangiogenic activity. Ad-IL-24 can downregulate angiogenesis-associated vascular endothelial growth factor and CD31 for reduction of angiogenesis to suppress hepatocellular carcinoma growth in vivo. Also, Ad-IL-24 functions antiangiogenically to decrease CD31/PECAM expression in a lung cancer model. Secreted IL-24 has been shown to have potent antiangiogenic activity both in vitro and in vivo (Saeki et al., 2002; Wang et al., 2007). Ad-IL-24 also has immune stimulatory activity, inducing high-level secretion of some helper T cell type 1 (Th1) cytokines such as IL-6, tumor necrosis factor (TNF)-α, and interferon (IFN)-γ and low-level secretion of IL-1β, IL-12, and granulocyte-macrophage colony-stimulating factor (GM-CSF) from human peripheral blood mononuclear cells (PBMCs), which functions as a Th1 cytokine by activating the immune system to improve antitumor efficacy (Caudell et al., 2002; Chada et al., 2004; Wu et al., 2007). The strong effect of IL-24 for inducing apoptosis, antiangiogenesis, and immunity led to the strong antitumor effect of Ad.sp-E1A(Δ24)-IL-24 presented in this paper (Fig. 6). Our data showed that IL-24 has a wide-ranging ability to kill tumor cells (Fig. 2). Moreover, comparison of the antitumor activity of gene therapy and targeting gene-virotherapy in vitro by MTT and crystal violet assays revealed that the antitumor activity of the gene–virus therapeutic drug Ad.sp-E1A(Δ24)-IL-24 is much higher (almost 100-fold) than that of regular Ad-IL-24 gene therapy in PLC and NCI-H1299 cells (Figs. 3 and 4); this showed that targeting gene-virotherapy is a more efficient mode of cancer therapy.
In summary, Ad.sp-E1A(Δ24)-IL-24 was successfully generated with a dual-regulated oncolytic adenovirus harboring IL-24. In in vitro experiments, Ad.sp-E1A(Δ24)-IL-24 was shown to efficiently express the IL-24 gene in tumor cells and to show an obvious broad tumor-specific cytotoxicity effect, but not in normal cells. In vivo, xenograft tumors were completely inhibited until they were smaller than they were before Ad.sp-E1A(Δ24)-IL-24 treatment, indicating that Ad.sp-E1A(Δ24)-IL-24 might be a potent antitumor agent.
Footnotes
Acknowledgments
The authors thank Prof. Cheng Qian for kindly providing plasmid pCN103, Dr. Miao Ding for kindly providing adenovirus Ad-IL-24, and also Xi Jun Liu, Jing Zhang, and Jing Qian for their kind help in cell culturing. This work was supported by the Zhejiang Science and Technology Support Plan (no. 2007C33027), the National Nature Science Foundation of China (nos. 30623003 and 30800093), the National Basic Research Program of China (973 Program; no. 2004 CB51804), Key Project funding from the Chinese Academy of Science (no. KSCX2-YW-R-09, R-04), and by Zhejiang Sci-Tech University grant 0616033 to Prof. Xin Yuan Liu.
Author Disclosure Statement
No competing financial interests exist.