
Abstract
High-level tissue-specific expression of recombinant proteins in muscle is an important issue for several therapeutic applications. To achieve this goal, we generated several constructs containing one to five copies of the upstream enhancer (USE) of 160-bp of the human slow troponin I gene, linked to that gene's minimal promoter. We also tested constructs made with one to four copies of a 100-bp deletion of USE (ΔUSE) reported to drive pan-muscle-specific expression in transgenic mice. These constructs were evaluated by measuring the activity of the reporter gene β-galactosidase (β-gal). In cell culture, these multimerized enhancers retained tissue specificity and their transcriptional strength increased with the number of enhancer copies. In myotube cultures (which still contain nondifferentiated cells), constructs containing four and five USE copies were stronger than the cytomegalovirus (CMV) early enhancer/promoter and slightly weaker than the hybrid CMV enhancer/β-actin (CB) promoter. Those containing three USE, or four ΔUSE copies were similar in strength to CMV. After electrotransfer of plasmid DNA into the mouse tibialis anterior muscle, the strengths of the two constructs (USEx3 and ΔUSEx3) were tested; as measured by β-gal activity in the total muscle lysate and by the number of transduced fibers, they were similar to CMV and CB. Muscle fiber typing, after electrotransfer of the soleus muscle, showed that ΔUSEx3 and USEx3 were active in slow and fast fibers. The tissue specificity of these two constructs was also evaluated by hydrodynamic plasmid injection through the tail vein. Although significant β-gal expression was measured in the liver when CMV was tested, no expression above background level was detected with USEx3 and ΔUSEx3. The strength, muscle specificity, and small size of these transcriptional elements render them very attractive for gene therapy applications.
Introduction
Troponin I (TnI) is an abundant component of the myofibrils of striated muscle. In adult vertebrates, two distinct isoforms, TnIslow and TnIfast, are differentially expressed at the transcriptional level in slow- and fast-twitch muscle fibers, respectively. The regulatory elements controlling the expression of TnIslow are relatively well characterized. A small upstream enhancer (USE) of 160-bp, also referred to as SURE, drives muscle-specific expression, preferentially in slow fibers (Corin et al., 1995; Levitt et al., 1995; Nakayama et al., 1996). Several transcription factors (MeF-2, MyoD/Myogenin, MusTRD1/GTF3, and Sp1) interact with this enhancer (Calvo et al., 2001; Polly et al., 2003; Issa et al., 2006). If a fragment of 100-bp is deleted at the 5′ end of USE, thus removing the binding site for MusTRD1/GTF3, the resulting enhancer (ΔUSE) remains specific for muscle, but with reduction or loss of slow muscle preference, that is, pan-muscle expression (Calvo et al., 2001). In the present study, to develop strong and muscle-specific promoters, we tested a battery of constructs containing several copies of USE or ΔUSE. We showed that multimerization of USE and ΔUSE upstream of the basal human TnIslow promoter generated powerful muscle-specific gene regulatory cassettes. Because of their strength and relatively small size (less than 1-kb), these multimerized TnIslow enhancer constructs could become instrumental for several gene therapy applications aiming at producing high-level and muscle-specific expression of therapeutic proteins.
Materials and Methods
DNA constructs
Plasmids were prepared using standard molecular biology methods. All constructs were verified by DNA sequencing. Plasmid encoding β-galactosidase (β-gal) regulated by the CMV early enhancer/promoter (pCMVβ; GenBank accession no. U02451) was purchased from Clontech Laboratories (Palo Alto, CA). This plasmid contains a 600 bp DNA fragment that encompasses the immediate early enhancer and promoter region (−522 to +73) of the genome of human CMV (Boshart et al., 1985). Also, this plasmid contains a small intron (96 bp) derived from SV40 late 19s/16s mRNA splice donor/splice acceptor and a SV40 polyadenylation signal (poly A). The same intron was present in all constructs tested except for pMCKβ and pCBLacZ. The same poly A was present in all constructs tested. Plasmid (pCBLacZ) encoding β-gal controlled by the CMV enhancer (position −598 to −217) linked to the chicken β-actin promoter and first intron (position −350 to +960) has been described (Molnar et al., 2004). This hybrid CMV/β-actin promoter is also referred to as CAG (Niwa et al., 1991). Plasmid expressing β-gal (pMCKβ) controlled by the murine muscle creatine kinase (MCK) enhancer/promoter fragment (−1354 to +1) was constructed by replacing the CMV enhancer/promoter and intron of pCMVβ with the MCK enhancer/promoter of pAdMCKBecker (Larochelle et al., 2002).
A plasmid (pTnIS(Im)β) expressing β-gal regulated by one copy of USE (−1036 to −873) linked to the human minimal TnIslow promoter and first exon (−95 to +56) was constructed by replacing the CMV enhancer/promoter of pCMVβ with the USE/promoter fragment of TnIsUSE-95X1nucZ (Corin et al., 1995) by EcoRI/KpnI digestion. Constructs containing two to five copies of USE were prepared by inserting additional USE enhancers into pTnIS(Im) β by polymerase chain reaction (PCR) using primers 5′-GTACTACTAGTACCGAGTCTTACGCGTGC-3′ and 5′-GGTACGTCTAGATAAGCTTCCCACTGCCCCCTCCTGC-3′ followed by XbaI/SpeI digestion before ligation. Construct pTnIS(Im)β-ΔUSE, containing one copy of ΔUSE (−940 to −873) linked to the TnIslow basal promoter and first exon, was made by amplifying ΔUSE using primers 5′-GCGGGGAGATCTGGATATCGATATAGGGTGGGTATTATG-3′ and 5′-GGTACGTCTAGATAAGCTTCCCACTGCCCCCTCCTGC-3′. The PCR product was digested with BglII and inserted into pTnIS(Im)β previously digested with BlgII to remove the USE enhancer. Constructs containing two to four copies of ΔUSE were made by inserting additional copies of ΔUSE by PCR. ΔUSEx2 and ΔUSEx4 were constructed using the same primers as for ΔUSEx1 followed by XbaI/SpeI digestion. ΔUSEx3 was constructed using the primers 5′-ATAGGCGCGCCGTACTAGTGACTATAGGGTGGGTATTATG-3′ and 5′-ATAATACGCGTTAAGCTTCCCACTGCCCCCTCCTGC-3′ followed by AscI/MluI digestion. The orientation of the USE and ΔUSE enhancers of all the constructs tested in this study was the natural one (5′ to 3′).
A plasmid (pTnIF(Im)β−) containing three copies of the first intron regulatory element (IRE) of the quail TnIfast gene in the reverse orientation (−) fused upstream to the minimal promoter of that gene and expressing β-gal was generated. The IRE (+658 to +804) was amplified by PCR using construct TnILacZ1 (Hallauer et al., 1993) as template and primers: 5′-CTAGTCGACGGCTGCGTCTGAGGAGACA-3′ and 5′-CTACTCGAGGCCAAGCTCCCTGAGGAA-3′. The PCR product was digested with XhoI and SalI, self-ligated, redigested with XhoI and SalI, and cloned into pBluescript II SK + (Stratagene, LaJolla, CA). The fragment containing the minimal promoter (−204 to +22) of the TnIfast gene (Hallauer et al., 1993) was subcloned from plasmid gCTnIf(−198: +22)Z downstream of the 3xIREs. The CMV enhancer/promoter of pCMVβ was then replaced with the 3xIRE and the minimal promoter.
Cell culture, transfection, and staining
293A (HEK-293), A549, HeLa, and C2C12 cells were obtained from ATCC (Rockville, MD). The cells were grown in Dulbecco's modified Eagle's medium supplemented with 5% or 10% (C2C12) fetal bovine serum. Plasmids were mixed with an equal amount of plasmid expressing the secreted alkaline phosphatase (SEAP) regulated by CMV, and the cells were transfected with Lypofectamine 2000 (Invitrogen, Carlsbad CA). The formation of myotubes was induced 24 hr after transfection as described (Gilbert et al., 2002). The cells were lysed 48 hr (293A, A-549, HeLa) or 11 days (C2C12 myotubes) after transfection. The myotube culture medium was replaced every 2 days. Consequently, the value of SEAP measured at day 11 corresponds to 2-day accumulation. The β-gal and SEAP activity was determined at the time of cell lysis as described previously (Molnar et al., 2004; Gaillet et al., 2007). For each sample, the transfection efficiency was normalized by dividing the β-gal value by the SEAP value. The SEAP value was used solely to normalize the transfection efficacy of the constructs within the same cell type. For each construct, the transfection experiment was repeated at least three times using three culture dishes each time (n = 9). Some cells were also fixed with 0.5% glutaraldehyde, and β-gal expression was then observed by incubation with X-gal (5-bromo-4-chloro-3-indolyl β-D-galatoside) using standard procedures. To evaluate the efficacy of C2C12 muscle differentiation, cultures of myotubes were fixed with 4% paraformaldehyde, permeabilized with 0.1% Triton X-100, and incubated with an antibody against myosin heavy chain (A4.1025, Developmental Studies Hybridoma Bank, University of Iowa) followed by a goat anti-mouse conjugated to Alexa Fluor 488 (Molecular Probes, Eugene, OR). The nuclei were then stained by incubation with Hoechst dye (Molecular probes).
Animal studies and statistical analysis
Animal experiments were performed according to McGill University and Biotechnology Research Institute guidelines for animal care. The tibialis anterior (TA) muscle of adult (5 to 6 weeks old) normal (CD1; Charles River Laboratories, Montreal, QC, Canada) mice were electrotransferred with plasmid and analyzed 10 days later as described previously (Molnar et al., 2004). For the soleus, plasmid was injected into surgically exposed muscle in a 2 μl volume at 1 μg/μl plasmid in PBS. After suturing, paddle electrodes (BTX model 520) adjusted to a 0.5 cm gap were placed on shaved skin on medial and lateral surfaces of the lower leg, using conductive gel (PolyGel plus from Medicaltronic) to improve contact. Eight 87 volt, 20 msec2 wave pulses were then delivered at 1 sec intervals using a BTX model ECM 830 electroporator. Mice were euthanized after 14 days. For fiber typing, serial cryostat sections were stained with monoclonal antibody BA-D5 that is specific for slow myosin heavy chain as described previously (Schiaffino et al., 1989; Hallauer and Hastings, 2002). To study tissue specificity of the TnIslow constructs in vivo, 4- to 6-week-old C57BL/6 mice (Charles River Laboratories) were injected through the tail vein using the hydrodynamic injection method (Liu et al., 1999; Zhang et al., 1999). Briefly, each mouse was injected rapidly (5 to 7 sec) through the tail vein with 2 ml of Ringer's solution containing 50 μg of plasmid DNA. The following plasmids were used: pUC19 (negative control), pCMVβ, and plasmids expressing β-gal regulated by USEx3 or ΔUSEx3. The mice were euthanized 24 hr postinjection. The liver was harvested, and lysed in β-gal buffer for reporter gene assay (Roche Molecular Biochemical, Mainnheim, Germany), and β-gal was measured by luminometry following the manufacturer's recommendations. Transduction efficacy was expressed as relative units of β-gal activity per μg of total protein determined by the Bio-Rad DC Protein Assay (Bio-Rad Laboratories, Hercules, CA). The data were analyzed using an analysis of variance followed by the Fisher's least significant difference procedure to compare the means. Some means were also compared with each other using the Student's t-test. Statistical significance was set at p < 0.05.
Results
Multimerization of USE and ΔUSE confers strong and muscle-specific expression in cell culture
During our initial search for strong muscle-specific promoters, we first compared the strength of USEx1, in which one copy of human USE was fused to the minimal promoter of the human TnIslow gene (Fig. 1A), with the short 1.35-kb MCK promoter (Larochelle et al., 1997, 2002) and a construct containing three copies of the IRE enhancer of the TnIfast (Hallauer and Hastings, 2002) linked to the minimal promoter of that gene. For these experiments, the strengths of the constructs were determined by transient transfection using β-gal as gene reporter in C2C12 myotubes. Because USEx1 was significantly stronger than MCK (Fig. 1B) and TnIfast (data not shown), we investigated whether the promoter strength could be increased further by linking together two or three copies of USE. In addition, because we are interested in expressing recombinant protein in all muscle fibers, we also generated constructs containing one to three copies of ΔUSE, a 100-bp deletion of USE reported to drive pan-muscle expression (Calvo et al., 2001). As demonstrated in Fig. 1C and D, the strengths of constructs made with multiple copies of USE and ΔUSE increased with the number of repeats.
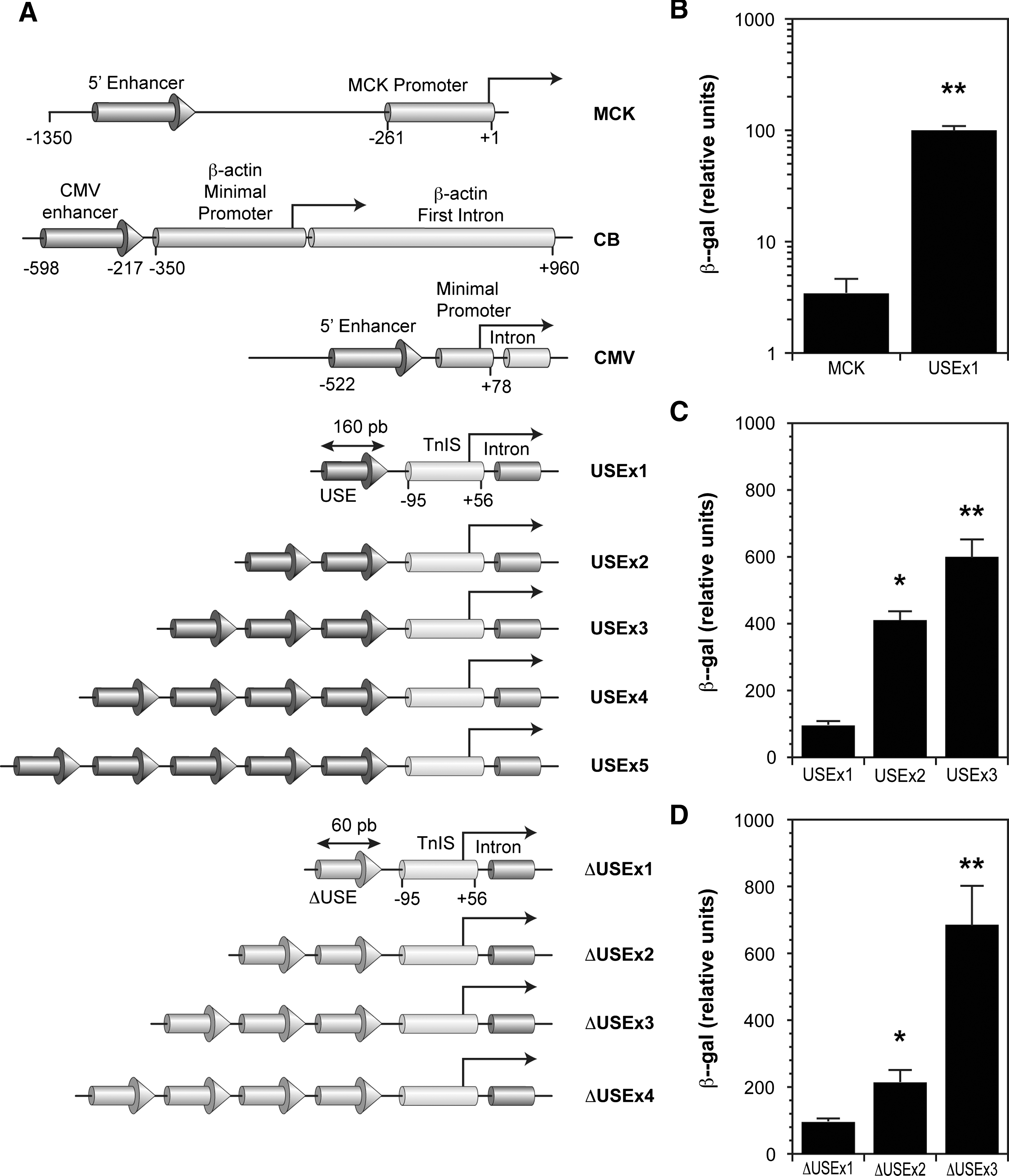
Multimerization of USE and ΔUSE increases the promoter strength in muscle culture. (
The most active constructs of each group (USEx3 and ΔUSEx3) were then cross-compared. For this experiment we also benchmarked their strength with the CMV early enhancer/promoter and with the hybrid CMV enhancer/β-actin (CB) promoter, also known as CAG. The latter promoter was used for comparison because it has been shown to be very strong in muscle cells (Ishii et al., 1999; Gilbert et al., 2002). These four constructs were also compared in nonmuscle cells (293A, HeLa and A549). For each cell type, the data were normalized to the value obtained with CMV, which was arbitrarily set to 100 (Fig. 2A). In differentiated myotubes, the activity of CB was the strongest. The activity of USEx3 was equivalent to CMV, but it was weaker than CB. Because CMV is known to be a very strong promoter, these data indicated that USEx3 drives strong expression in muscle. The activity of ΔUSEx3 was weaker, since it corresponded to 40% of CMV and USEx3. The strength of ΔUSEx3 and USEx3 in myotubes was confirmed by staining the cells for β-gal (Fig. 2B). In nonmuscle cells, the activity of USEx3 and ΔUSEx3 was 50 to 100 times lower than CMV and CB, thus demonstrating their muscle specificity (Fig. 2A).

USEx3 and ΔUSEx3 confer efficient muscle-specific expression in cell culture. (
Strength of USEx3 and ΔUSEx3 in mouse muscle in vivo
We then tested the activities of USEx3 and ΔUSEx3 in mouse muscle in vivo. The TA of adult mice was injected with plasmids encoding β-gal regulated by USEx3 or ΔUSEx3. This was followed by the application of pulses of electric current (electrotransfer) as described previously (Molnar et al., 2004). Electrotransfer is known to dramatically increase the transduction efficiency of naked DNA in muscle. As a control, muscles were also injected in a similar manner with plasmids carrying β-gal regulated by CB or CMV. At 10 days postinjection, the level of β-gal in the injected muscle was determined by luminometry (Fig. 3A). Sections of injected muscles were also processed for β-gal histochemistry to reveal the number of transduced (β-gal positive) muscle fibers (Fig. 3B and C). All of the four constructs tested (CMV, CB, USEx3, and ΔUSEx3) generated similar amounts of β-gal and similar numbers of transduced fibers after electrotransfer. Overall, the transduction efficiency was high because an average of 200 to 300 fibers was positive for β-gal, which represents 15% of the total number of fibers in the TA. Because the TA consists of more than 95% fast fibers (Nakayama et al., 1996), these data suggest that USEx3 and ΔUSEx3 were both active in fast muscle fibers. To confirm that both constructs were active in fast and slow fibers, the soleus muscle (a mixture of slow and fast fibers) (Corin et al., 1995; Nakayama et al., 1996) was electrotransferred with plasmids encoding β-gal regulated by USEx3 and ΔUSEx3. Serial cryostat sections were processed for β-gal histochemistry and stained with a monoclonal antibody against slow myosin heavy chain. β-gal expression was detected in slow as well as in fast muscle fibers, indicating that USEx3 and ΔUSEx3 were active in both types of fibers (Fig. 4). We next compared the strength of USEx3, ΔUSEx3, and CMV in the liver following hydrodynamic injection of plasmid DNA through the tail vein as described previously (Liu et al., 1999; Zhang et al., 1999). β-gal expression was determined by luminometry at 24 hr postinjection because transgene expression using this method is higher at early time points (Liu et al., 1999). In agreement with our cell culture data that showed weak expression in nonmuscle cells, the strength of USEx3 and ΔUSEx3 in the liver was significantly lower compared with CMV (Fig. 5). In fact, the β-gal value obtained with the TnIslow constructs was not significantly different from the value obtained with an empty plasmid.
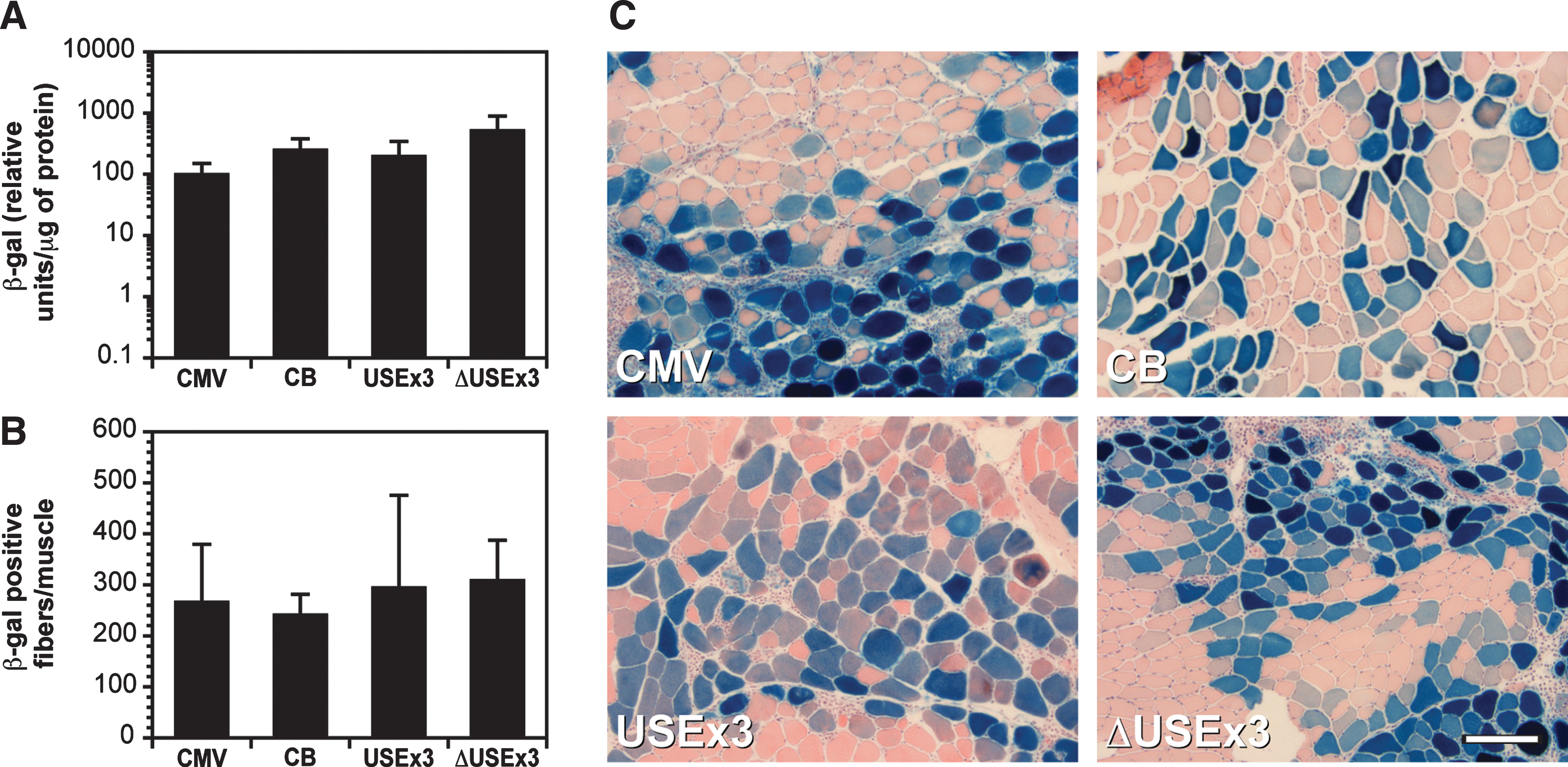
USEx3 and ΔUSEx3 confer efficient expression in mouse muscle in vivo. The tibialis anterior muscle of normal mice was injected with plasmids encoding β-gal under the control of CMV, CB, USEx3, or ΔUSEx3 followed by electrotransfer. The relative amount of β-gal produced per muscle (
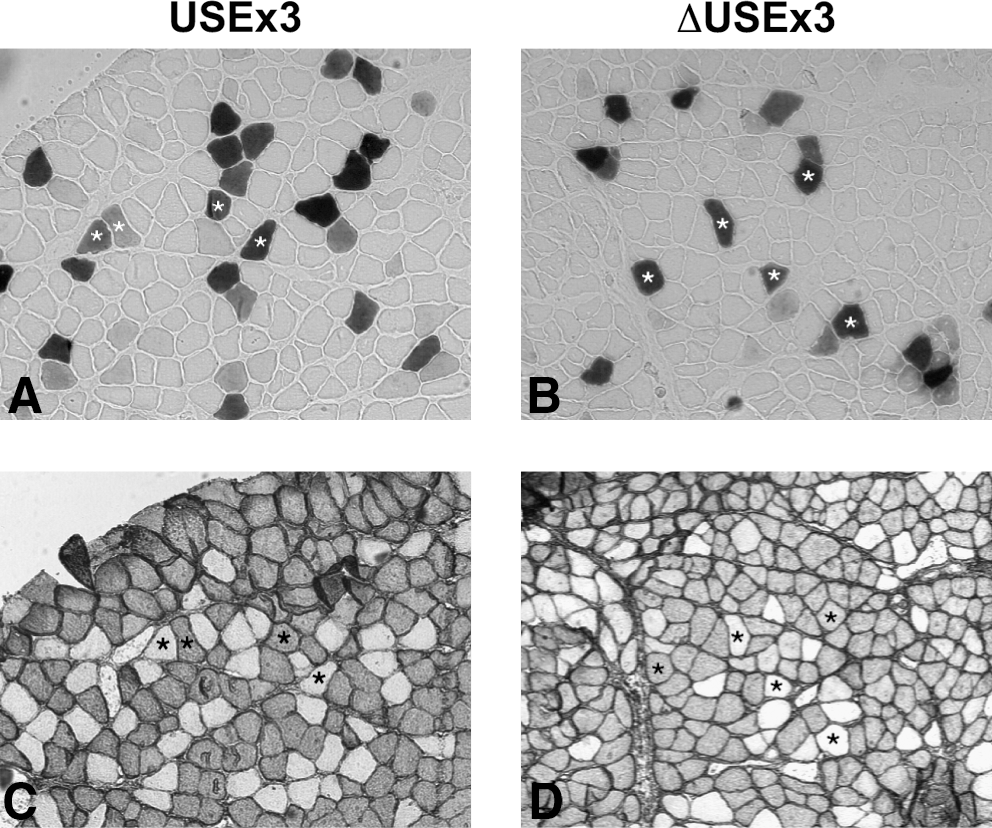
Activity of USEx3 and ΔUSEx3 in fast and slow muscle fiber. The soleus muscle of normal mice was injected with plasmid encoding β-gal regulated by USEx3 (

USEx3 and ΔUSEx3 are weakly active in the liver in vivo. Normal mice were injected through the tail vein using the hydrodynamic method with an empty plasmid (Cont) or with plasmids expressing β-gal regulated by CMV, USEx3, and ΔUSEx3. Twenty-four hours later, the liver was analyzed for β-gal expression by luminometry. The data are the mean ± SEM; n = 5. *Data significantly lower than CMV. USEx3 and ΔUSEx3 were not significantly different from empty plasmid.
Increased muscle-specific expression with additional copies of USE and ΔUSE
To verify whether it was possible to obtain constructs stronger than USEx3 and ΔUSEx3 in muscle cells, we generated constructs containing four or five copies of USE, or four copies of ΔUSE (Fig. 1A). These constructs were tested in cultures of myotubes using plasmids encoding β-gal as described above (Fig. 6A). For comparison, the cells were also transfected with plasmids carrying CB, CMV, USEx3, and ΔUSEx3. The data showed that USEx4 was stronger than USEx3 and CMV, but it was weaker than CB. Increasing the number of USE to five copies did not significantly increase the strength further. In differentiated myotubes, the strength of ΔUSEx4 was comparable to CMV and it was thus better than ΔUSEx3. These promoters were also evaluated in 293A cells (Fig. 6B). Additional copies of USE or ΔUSE did not increase their strength in nonmuscle cells, since USEx4, USEx5, and ΔUSEx4 were 150 to 200 times weaker than CMV in these cells.
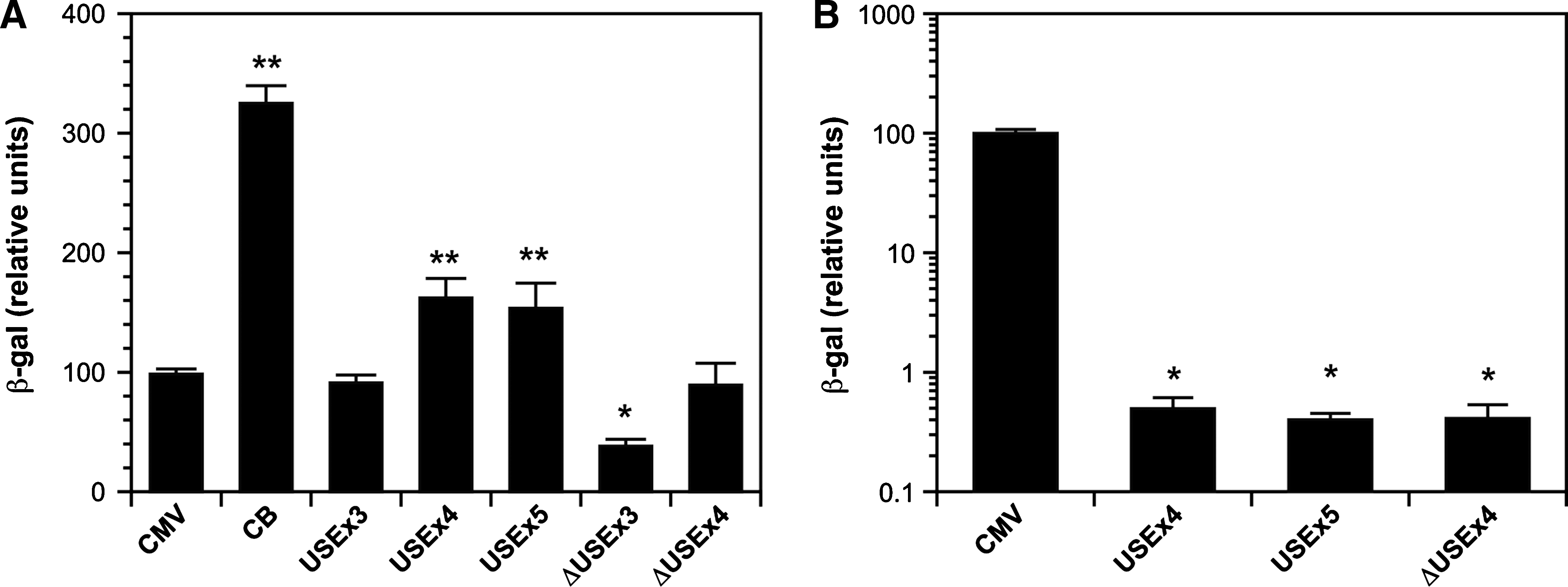
Further increase of muscle-specific expression is possible using four copies of USE or ΔUSE. (
Discussion
In the present study we demonstrated that strong muscle-specific constructs can be generated by multimerizing USE and ΔUSE upstream of the basal TnIslow promoter. We observed a strong correlation between the strength of the construct and the number of enhancer repeats utilized. Maximal strength was obtained by linking together four or five copies of USE enhancer. The strength of USEx5 was not significantly different from USEx4, indicating that an upper limit was reached with four enhancer copies. In myotube cultures, two constructs (USEx4 and USEx5) were more active than CMV and two constructs (USEx3 and ΔUSEx4) had the same activity. None of our constructs, however, were as strong as CB, which has been demonstrated to be very active in skeletal muscle (Ishii et al., 1999; Gilbert et al., 2002). The relative strength of the TnIslow constructs in comparison to CMV and CB is most likely underestimated, because myoblasts were transfected and that CMV and CB are more active in myoblasts than TnIslow constructs (data not shown), which would allow more time for β-gal to accumulate when CMV and CB are used. In addition, even after 11 days of differentiation, 49% of the myonuclei were not in a differentiated state, as measured by staining the cells with an antibody against myosin heavy chain. Consequently, a significant percentage of cells, even if they were successfully transfected with the TnIslow constructs, were not optimally expressing β-gal. We faced a similar situation with the in vivo experiments. Although the bulk of the muscle mass consists of differentiated fibers, nonmuscle cells (satellite cells, cells from connective tissues and capillaries) might be transduced as well. This may increase the relative strength of CMV and CB because they are active in nonmuscle cells, which is not the case for the TnIslow constructs. Despite the limitation of our experimental system, we can state that ΔUSEx3 and USEx3 are strongly active in skeletal muscle, because their strength (β-gal activity and number of transduced fibers) was comparable to CB and CMV.
The series of TnIslow constructs described in this study appear to be muscle specific in that they were very active in muscle (differentiated C2C12 myotubes and TA) and poorly active in nonmuscle cells (293A, HeLa, A549, and liver tissue). Because of their strength and specificity for muscle, these TnIslow constructs possess attractive properties for gene therapy of muscle diseases or for the production of proteins with therapeutic value (blood coagulation factor, erythropoietin, etc.) by the skeletal muscle. Only a few muscle-specific promoters as strong as or better than CMV have been reported in the literature (Li et al., 1999; Salva et al., 2007; Wang et al., 2008). The series of USE and ΔUSE promoters described here represents a valuable alternative to these promoters for achieving strong muscle-specific expression.
In transgenic mice and in vivo gene transfer studies, the USE enhancer has been shown to drive preferential expression in slow-twitch muscle fibers, whereas the ΔUSE enhancer shows little fiber type specificity in transgenic mice (Corin et al., 1995; Nakayama et al., 1996; Calvo et al., 2001). We have evaluated the fiber type specificity of USEx3 and ΔUSEx3 in the soleus muscle that consists of a mixture of fast and slow fibers. As expected, ΔUSEx3 was active in fast as well as slow fibers. Surprisingly, however, USEx3 was also active in both fiber types. This is in agreement with the results obtained in the TA, a muscle that consists largely of fast fibers (Nakayama et al., 1996; Calvo et al., 2001), where both constructs were clearly active with a strength comparable to that of strong viral enhancer/promoter constructs. It is possible that enhancer multimerization or other construct features may account for the unexpected expression of USEx3 in fast, as well in slow, muscle fibers. However, any such effects do not compromise overall specificity for muscle tissue, as assessed in cell culture and in vivo. Thus, not only ΔUSE constructs but also the multimerized USE constructs may be useful for gene therapy applications requiring broad expression in skeletal muscle. Another important practical characteristic of the TnIslow constructs is their small size, which renders them very attractive for vectors with limited transport capacity (such as recombinant AAV). The size (without the intron) of our strongest constructs, USEx4 and ΔUSEx4, is 1000 and 600 bp, respectively. To facilitate their construction, we inserted linkers of 40 to 80 bp between the USE and ΔUSE repeat units. Without such linkers, the size of USEx4 and ΔUSEx4 could be reduced to 800 and 450 bp, respectively.
In conclusion, we showed that strong and muscle-specific expression can be obtained by combining several copies of USE and ΔUSE upstream of the basal TnIslow promoter. The strength of these constructs and their relative small size should make them very useful for various gene therapy applications.
Footnotes
Acknowledgments
We would like to thank Carol Allen, Stephen Prescott, and Antoine W. Caron for their expert technical assistance. This work was supported by grants (ITM 71879 and MOP 81296) from the Canadian Institutes of Health Research (CIHR) to R.G. and by a grant (FRN 57864) from the CIHR/Amyotrophic Lateral Sclerosis Society of Canada/Muscular Dystrophy Canada Neuromuscular Research Partnership program to K.E.M.H. This is an NRC publication number 47828.
Author Disclosure Statement
M.Bl., B.M., and R.G have submitted a patent for the multimerized TnIslow constructs. No competing financial interests exist for Y.Z., M.Be., A.K, P.L.H, K.E.M.H., and G.K.