
Abstract
In vivo electroporation of plasmid DNA (DNA-EP) is an efficient and safe method for vaccines. It results in increased DNA uptake, enhances protein expression, and augments immune responses to the target antigen in a variety of species. To further improve the efficacy of DNA-EP, we evaluated small interfering RNA (siRNA) sequences targeting apoptotic genes as an adjuvant to cancer vaccine. Bak1 or Casp8 siRNA was coadministered with plasmid DNA encoding the extracellular and transmembrane domains of rat HER2 ECD.TM to BALB-neuT mice, which spontaneously develop HER2/neu-positive mammary tumors. The combination regimen significantly reduced spontaneous tumor progression in BALB-neuT mice, in an advanced disease setting, when compared with DNA-EP alone. The antitumor effect was associated with a noteworthy antibody isotype switch from IgG1 to IgG2a, when siRNA was coadministered with DNA-EP. CD8+ T cell responses increased significantly, as did the number of responders to vaccination. Coimmunization of siRNA and DNA-EP at the same physical location was essential for the enhanced therapeutic effect. Silencing of the targeted genes was confirmed by in vitro Western blots. siRNA sequences targeting apoptotic genes Bax and Fas did not improve tumor protection in this mouse model when compared with DNA-EP alone. These data demonstrate that some siRNA sequences can act in concert with DNA-EP to control HER2/neu-positive mammary carcinoma. These observations emphasize the potential of siRNA as adjuvant for therapeutic DNA vaccines.
Introduction
Enhancement of immunologic potency is one of the key objectives of DNA vaccine research today and this aim is currently being pursued through a variety of different yet complementary strategies (Yu and Finn, 2006). Among those being tested, in vivo electroporation of plasmid DNA (DNA-EP) has been shown to be a safe method resulting in greater DNA uptake, leading to enhanced protein expression in the treated muscle (Cappelletti et al., 2003). This has been associated with an increase in immune responses to the target antigen in a variety of species (Widera et al., 2000; Zucchelli et al., 2000; Babiuk et al., 2002; Scheerlinck et al., 2004; Aurisicchio et al., 2007). A number of adjuvants such as cytokines, costimulatory molecules and chemotherapeutic agents are being tested both in the laboratory and the clinic (Comes et al., 2006; Hodge et al., 2006; Overwijk et al., 2006; Taieb et al., 2006; Arcuri et al., 2008). Among them are small interfering RNA sequences, which are being used to harness the ability of the immune system to respond and develop a stable, long-lasting immune response against tumor antigens (Shen et al., 2004).
The process of posttranscriptional double-stranded RNA (dsRNA)-dependent gene silencing is called RNA interference, or RNAi (Zamore et al., 2000). RNAi is triggered by the presence of 21- to 23-nucleotide-long dsRNA in the cell and results in the rapid destruction of the mRNA containing identical or nearly identical sequences. In nature, dsRNA is recognized by Toll-like receptor-3 (TLR3). On recognition, TLR3 induces the activation of NF-κB to increase production of type I interferons, which signal other cells to activate their antiviral defenses (Alexopoulou et al., 2001). RNAi can be induced in vitro by synthetic 20- to 25-nucleotide RNA sequences (small interfering RNA, or siRNA) (Elbashir et al., 2001). RNAi in adult mice with siRNAs targeting luciferase is well characterized (McCaffrey et al., 2002). Data suggest that siRNAs could be used for induction of cytokines such as interferon-α by plasmacytoid dendritic cells via TLR7 signaling (Hornung et al., 2005). siRNA-mediated knockdown of Notch ligands in primary CD4+ T cells and dendritic cells enhanced cytokine production in mice (Stallwood, 2005). Silencing of SOCS1 (suppressor of cytokine signaling) in dendritic cells enhanced anti-HIV immunity in mice (Song et al., 2006). Transduction of antigen-presenting cells (APCs) with siRNAs targeting apoptotic genes Bak and Bax has been shown to enhance cancer vaccine potency (Peng et al., 2005). Coadministration of DNA vaccines encoding human papillomavirus type 16 E7 with siRNA targeting the same proapoptotic genes prolonged the lives of antigen-expressing dendritic cells in the draining lymph nodes, enhanced antigen-specific CD8+ T cell responses, and elicited potent antitumor effects against an E7-expressing tumor model in vaccinated mice (Kim et al., 2005). Both Bak1 and Bax belong to the Bcl-2 family of genes and are potent regulators of cell death (Karbowski et al., 2006). Other genes such as Casp8 and Fas in the apoptotic pathway have also been targeted (Wesche-Soldato et al., 2005). These studies provided the groundwork for the testing of siRNA as adjuvants in our study.
For cancer vaccines, the most widely used preclinical models for immunologic and antitumoral studies are transgenic rodents expressing human tumor-associated antigens (TAAs) because they show central and/or peripheral tolerance to them (Lucchini et al., 1992; Clarke et al., 1998). Among them, HER2/neu is a robust mouse model. The HER2/neu oncoprotein is a Tyr-kinase receptor overexpressed in several human tumors and associated with poor prognosis (Slamon et al., 1987, 1989). Female BALB/c mice transgenic for the mutated rat HER2 gene (BALB-neuT) expressed specifically in the mammary glands are an optimal model for HER2-mediated breast cancer (Boggio et al., 1998; Lollini et al., 2006). DNA vaccination has been shown to be effective as a prophylactic method in BALB-neuT mice. Numerous studies have demonstrated the prevention of spontaneous development of mammary tumors occurring in these mice at an early age (Lollini et al., 2006). The combination of cytokines or immunoregulatory molecules, together with HER2/neu-encoding DNA plasmid, has been shown to increase the effectiveness of the HER2/neu vaccines (Nanni et al., 2003; De Giovanni et al., 2004; Quaglino et al., 2005). We have shown that a TLR7 agonist, SMO36020, is a potent vaccine adjuvant in transgenic mouse models (Dharmapuri et al., 2008).
In this study, we evaluated DNA-EP vaccination in combination with siRNAs in a well-characterized mouse model for the development of HER2/neu cancer vaccines. We demonstrate that siRNAs targeting the proapoptotic genes Bak1 and Casp8 significantly impact tumor protection in BALB-neuT mice when combined with DNA-EP and characterize the immunologic parameters associated with this effect. This combination treatment delays tumor progression in an advanced and more aggressive disease setting. In vitro data confirmed that silencing was target specific. Hence, we propose that specific siRNA sequences can act as immunological adjuvants and synergize with DNA vaccination.
Materials and Methods
Mice and DNA immunizations
Female BALB/c mice (H-2d
) transgenic for the transforming neuT oncogene (neuT+/neuT+) under the transcriptional control of the mouse mammary tumor virus (MMTV) promoter and transgene negative (neuT–/neuT–) were bred under specific pathogen-free conditions by Charles River (Calco, Italy). Mice were screened for the presence of the transgene by duplex polymerase chain reaction (PCR) as previously described (Gallo et al., 2005). The ECD.TM region (comprising the first 679 amino acid residues) of a codon-optimized version of the rat neu gene was cloned under the control of the human cytomegalovirus (CMV) promoter in the plasmid pV1J/rHER2ECD.TM. In general, 5 μg of each siRNA duplex was mixed with 50 μg of DNA and administered intramuscularly with electroporation (EP) at the same site unless indicated otherwise. DNA and siRNA were injected into the quadriceps muscle (in 50 μl of physiological solution) and electrostimulation was performed as previously described (Rizzuto et al., 1999). Ten trains of 1000 bipolar pulses (130 V, 75 mA, 200 μsec/phase) were delivered by an electroporator through electrodes inserted into the mouse quadriceps (instrument and software manufactured by Eurika, Crotone, Italy). The siRNA sequences used were synthesized by Sigma-Aldrich (St. Louis, MO). The sequences used were as follows (5′ → 3′, only sense sequences described): Bak1-1, UGCCUACGAACUCUUCACCUU, Bax, UAUGGAGCUGCAGAGGAUGUU; nonspecific or control, NNAUUGUAUGCGAUCGCAGAC (Kim et al., 2005); SOCS1, CUACCUGAGUUCCUUCCCCUU (Shen et al., 2004); Casp8, UGCCCUCAAGUUCCUGUGCUUGGAC; and Fas, AAAGUGGCCCAUUUAACAGGC (Wesche-Soldato et al., 2005). Bak1-2 (GAGACUGCAUGACCUACUUUU), Bak1-3 (CCAAGAUCGCCTCCAGCCUUU), and Bak1-4 (GGUCUUUCGAAGCUACGUUUU) were identified using standard databases (
Evaluation of spontaneous tumor growth in BALB-neuT mice
BALB-neuT females were inspected weekly and tumor masses were measured with calipers in two perpendicular diameters. Progressively growing masses >3 mm in mean diameter were regarded as tumors. Growth was monitored until all mammary glands displayed a palpable tumor or tumor diameter exceeded 10 mm, at which time mice were killed for ethical reasons. Except where otherwise specified, surviving BALB-neuT mice were killed at week 30.
Intracellular cytokine staining
Interferon (IFN)-γ and tumor necrosis factor (TNF)-α production was measured as previously described (Gallo et al., 2005; Mennuni et al., 2005; Giannetti et al., 2006). Briefly, 1–2 million mouse peripheral blood mononuclear cells (PBMCs) were isolated from blood by standard protocols in RPMI 1640 medium. Cells were incubated overnight at 37°C with appropriate amounts (5–6 μg/ml) of a known immunodominant CD8+ epitope of rat Neu (TYVPANASL) and brefeldin A (1 μg/ml; BD Sciences, San Jose, CA). Cells were washed and stained with surface antibodies APC (allophycocyanin)–anti-mouse CD3ɛ, PE (phycoerythrin)–anti-mouse CD4, and PerCP (peridinin chlorophyll protein)-anti-mouse CD8α. Lymphocytes were then permeabilized and incubated with fluorescein isothiocyanate (FITC)-conjugated IFN-γ or TNF-α antibody (all antibodies from BD Sciences). Cells were analyzed with a FACSCalibur flow cytometer, using CellQuest software (BD Biosciences).
Titration of anti-Neu humoral response
Antibodies against Neu were measured by enzyme-linked immunosorbent assay (ELISA). Ninety-six-well plates were coated with anti-hFc antibody (Pierce Biotechnology, Rockford, IL) overnight at 4°C. Supernatant from a HEK-293 cells expressing a soluble fusion protein, rat neuECD-hFc, was layered over the antibody followed by sera from DNA-EP-immunized mice. Alkaline phosphatase-linked antibodies against total IgG1 or IgG2a were used for detection (BD Sciences). Plates were read on an ELISA reader to determine the optical density (OD) at 405–620 nm. Serum dilution was plotted versus optical density values and fitted with a Michaelis–Menten curve using KaleidaGraph (version 3.5; Synergy Software, Reading, PA).
Western blotting
HeLa cells were transfected with various concentrations of siRNA with Oligofectamine (Invitrogen, Carlsbad, CA). Cells were treated with staurosporine to induce apoptosis (Hartel et al., 2003). Total proteins were isolated by standard protocols, quantitated, and run on sodium dodecyl sulfate (SDS)–polyacrylamide gels. Bak1 and Casp8 proteins were detected by respective monoclonal antibodies (Cambridge Antibody, Cambridge, UK) by standard methods.
Statistical analysis
Statistical analysis was performed as described previously (Cipriani et al., 2008). Tumor-free survival or time to first tumor appearance, tumor multiplicity (average number of tumors at 26 weeks), and time to advanced disease (time to palpable tumors in 50% of the mammary glands) were the measures of analysis by log-rank test. Data for which p values were less than 0.05 were considered statistically significant.
Results
Target-specific silencing predicts adjuvant potency
In a pilot in vitro study, the silencing efficacy of four siRNA sequences was assessed. HeLa cells were transfected with siRNA duplexes. The effect on Bak1 and Casp8 protein expression showed that the most effective siRNAs, Bak1-1 and Casp8, silenced their targets at low concentrations, starting from 10 pmol (Fig. 1A). Other siRNAs that targeted Bak1 were less effective. Bak1-2, for instance, needed to be used at 50 pmol to show any silencing effect as detected by Western blotting. This experiment also showed that Bak1 siRNAs could be ranked in order of silencing efficacy (from highest to lowest) as Bak1-1 > Bak1-3 > Bak1-2. Casp8 siRNA showed silencing efficacy similar to Bak1-1 at 10 pmol.
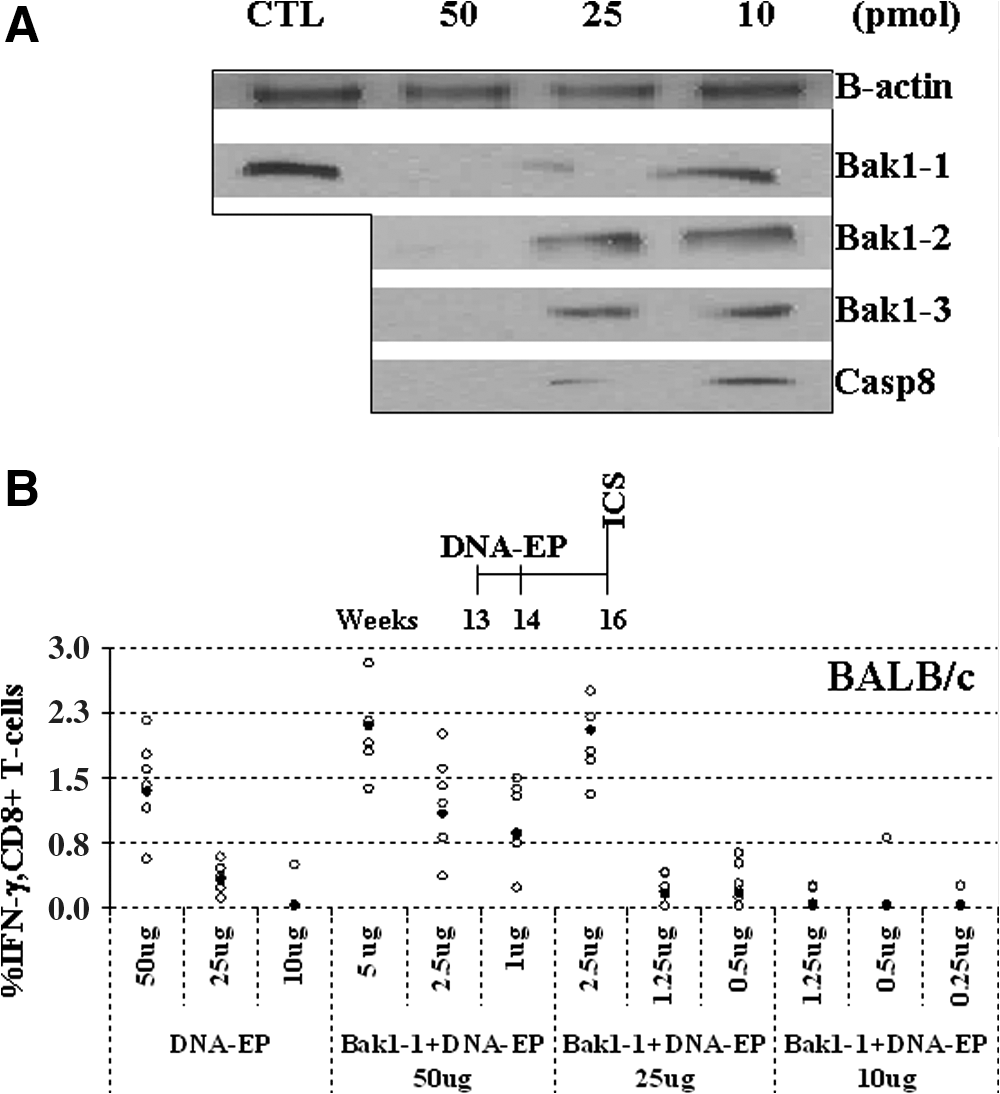
In vitro silencing with small interfering RNA (siRNA) and impact of antiapoptotic siRNA on the immune response elicited by electroporation of plasmid DNA (DNA-EP) in BALB/c mice. (
Bak1-1 siRNA increases immune response in BALB/c mice
In vivo studies were performed in 13-week-old BALB/c mice, using plasmid-encoded rat neu ECD.TM as antigen. They were immunized twice (1-week interval) with various amounts of DNA and siRNA followed by EP (DNA:siRNA, from 10:1 to 2:1 in micrograms). The T cell response was analyzed 2 weeks after the second vaccination. Figure 1B shows that 50 μg of DNA-EP per se elicited the highest T cell response as compared with lower amounts of DNA as measured by IFN-γ intracellular cytokine staining (ICS) (p < 0.05). Addition of 5 μg of Bak1-1 siRNA duplex increased the T cell response to more than that with DNA-EP alone. The reactivity observed in response to 25 μg of DNA-EP plus 2.5 μg of Bak1-1 siRNA (mean, 2%; 100% responders defined as those that have at least 0.1% cytokine-secreting CD8+ cells) relative to that of 25 μg of DNA-EP plus 1.25 μg of Bak1-1 siRNA with almost no responders was significantly higher (p < 0.05). This suggested that Bak1-1 siRNA could be having an effect on cell-mediated immunity and could be used at a 1:10 ratio with DNA. As 50 μg of DNA-EP elicited the highest immune response (without siRNA), all further experiments were carried out with 50 μg of DNA-EP and 5 μg of siRNA duplexes. Five micrograms of nonspecific siRNA had no effect on the immune response elicited by 50 μg of DNA-EP (data not shown).
Bak1-1 and Casp8 siRNAs show adjuvant activity with DNA-EP in BALB-neuT mice
The antitumor effect of siRNA/DNA-EP was assessed in BALB-neuT transgenic mice. Five micrograms of siRNA duplex was mixed with 50 μg of DNA (pV1J-neuECD.TM) and delivered by EP into BALB-neuT mice. Figure 2 shows the antitumor response after DNA-EP and siRNA immunization at 13 and 14 weeks of age, represented as Kaplan–Meier survival curves. At this age, mice display advanced atypical hyperplasia in their mammary glands. Untreated mice begin to develop palpable tumors by 15 weeks of age and all animals have tumors by week 21 (Boggio et al., 1998). Two DNA-EP injections delivered with a nonspecific siRNA at 13 and 14 weeks of age did not significantly slow tumor development compared with untreated mice. By week 22, all the mice developed tumors and had at least eight per mouse. When DNA-EP was supplemented with siRNAs targeting the proapoptotic gene Bak1 there was a distinct increase in the number of tumor-free mice. siRNA Bak1-1 elevated the number of tumor-free mice to 50% and the mean tumor number stabilized at four per mouse at 30 weeks of age (Fig. 2A). On the other hand, siRNAs targeting Bax and SOCS1 had no effect and the relative lack of tumor protection was similar to that of the DNA-EP and nonspecific siRNA (control) group. Bax is a proapoptotic gene and SOCS1 plays a prominent role in cytokine signaling. Published data suggested that they could be suitable targets but did not prove to be so in this mouse model. Mice that were treated with any of the siRNAs alone showed similar rates of tumor development as the saline control and indicated that siRNA per se did not show any nonspecific antitumor activity (data not shown).
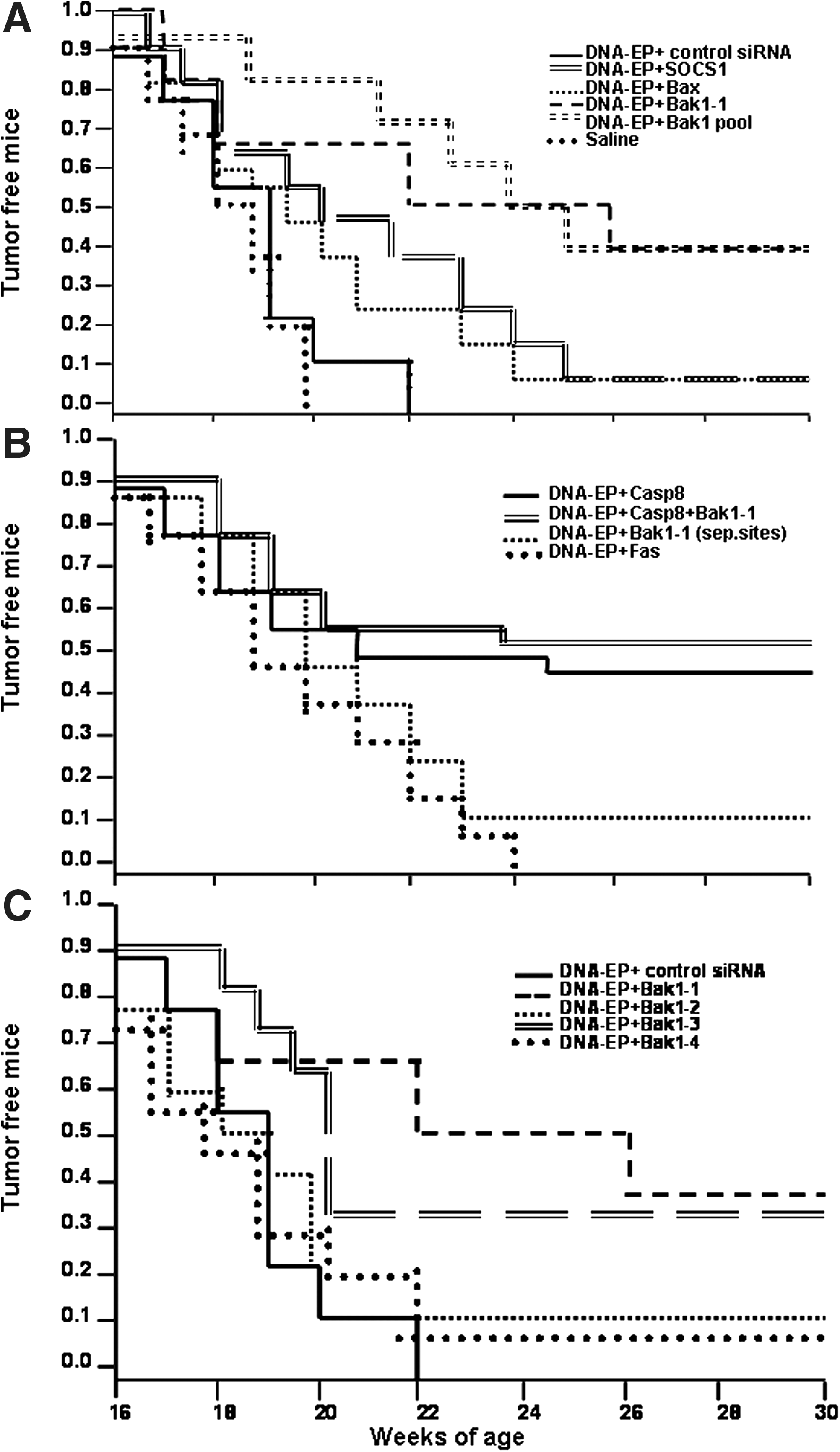
Impact of antiapoptotic siRNA on antitumor response elicited by DNA-EP in BALB-neuT mice. Mice were vaccinated with 50 μg of DNA and 5 μg of siRNA, both delivered intramuscularly with EP at 13 and 14 weeks of age. Mice that received two siRNAs simultaneously were administered 5 μg of each siRNA mixed with 50 μg of DNA. Tumor development was monitored weekly up to 30 weeks of age. (
The administration of siRNA targeting another apoptotic gene, Casp8, showed a similar effect as Bak1-1 (Fig. 2B). Almost 50% of the mice remained tumor free at 30 weeks and animals had three or four tumors each. When both were administered simultaneously with DNA-EP, there were no further significant additive effects over what was observed when they were used individually. As DNA-EP delivered with a nonspecific siRNA was equivalent to DNA-EP, it indicated that most likely targeted genes were being silenced in the other groups. siRNA targeting Fas did not enhance tumor protection when codelivered with DNA-EP. A separate group was immunized with DNA-EP and Bak1-1 siRNA at physically separate locations (on right and left quadriceps). These mice developed tumors at the same rate as those immunized with DNA-EP and nonspecific siRNA. This suggested that coimmunization of both reagents is critical.
To confirm the results obtained and to exclude potential effects mediated by off-target activity, three other siRNAs targeting Bak1 were tested. Of them, Bak1-3 was somewhat effective, with about 30% of the mice remaining tumor free at 30 weeks of age. The two other sequences tested did not improve tumor protection (Fig. 2C). The results were subjected to statistical analysis by log-rank and Wilcoxon tests (Table 1). DNA-EP plus Bak1-1 siRNA, alone or as part of a Bak1 siRNA pool (Fig. 2A), significantly extended tumor-free survival and delayed the time to advanced disease (p < 0.05). With Bak1-3 siRNA, DNA-EP significantly extended tumor-free survival (p < 0.05) but did not delay advanced disease. DNA-EP plus Casp8 siRNA moderately extended tumor-free survival and significantly delayed advanced disease (p = 0.02). DNA-EP plus Bak1-2, Bak1-4, or Fas siRNA did not significantly delay tumor-free survival or the time to advanced disease. Figure 3 shows tumor multiplicity at 26 weeks. Bak1-1 siRNA reduced the number of tumors compared with DNA-EP alone whereas Bak1-3 did not significantly reduce the number of tumors at 26 weeks. Bak1-2, Bak1-4, and Fas siRNA had no impact on tumor counts at 26 weeks. These data showed that Bak1-1 and Casp8 siRNAs were the best candidates for further adjuvant characterization.

Tumor counts at 26 weeks of age in BALB-neuT mice. Mice were vaccinated with 50 μg of DNA and 5 μg of siRNA, both delivered intramuscularly with EP at 13 and 14 weeks of age. Tumor development was monitored weekly and the median tumor count at 26 weeks of age is depicted. The ends of each box represent the 75% confidence interval and the line in the middle indicates the median. The whiskers indicate the 95% confidence interval. Ten mice were analyzed in each group and each siRNA was tested in at least two independent experiments.
Log-rank test, two-sided p value. Bold face values indicate significance.
Wilcoxon test, two-sided p value. Bold face values indicate significance.
Effect on cell-mediated immunity in BALB-neuT mice
To monitor the immunologic effect of Bak1-1 and Casp8 siRNAs in combination with DNA-EP, we measured the cell-mediated immune response by intracellular staining. The percentage of CD8+ T cells that secreted IFN-γ or TNF-α was measured in peripheral blood lymphocytes 2 weeks after the second DNA-EP plus siRNA injection at week 16 and again 7 weeks later. Figure 4A shows that Bak1-1 and Casp8 siRNAs contributed to an increase in number of responders when compared with DNA-EP alone. At week 16, about 50% of mice showed detectable IFN-γ- or TNF-α-secreting T cells. When either Bak1-1 or Casp8 siRNA was administered as adjuvant, the percentage of responders increased to more than 80%. A significant effect was best observed at week 16 (mean, 0.7 and 0.5% IFN-γ for Bak1-1 and Casp8, respectively). When both siRNAs were administered simultaneously, there was again a significant additive effect for the percentage of IFN-γ cells (mean, 0.5%), although not over and above the same with individual siRNAs.
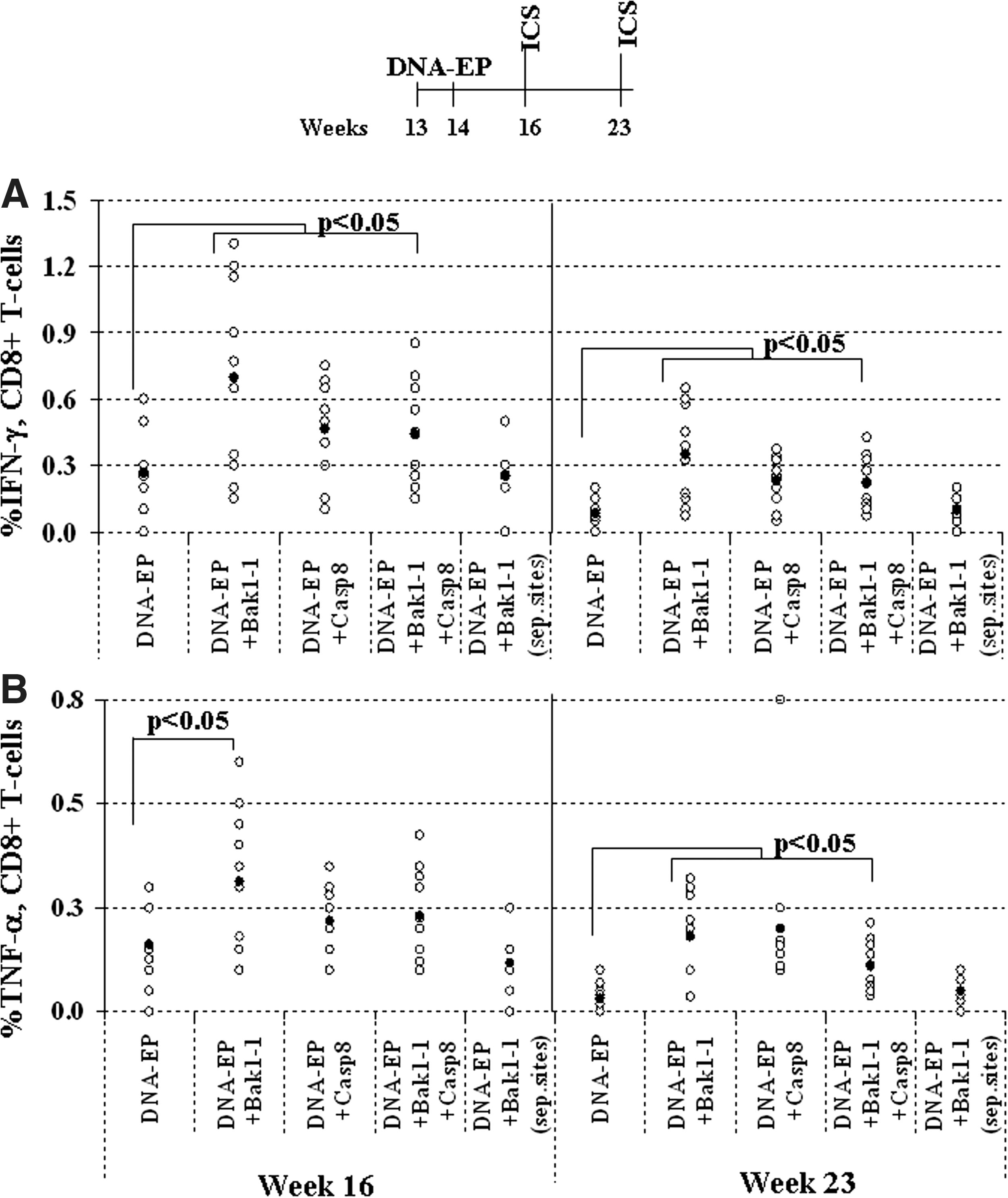
Antiapoptotic siRNA significantly increases cell-mediated immunity after DNA-EP in BALB-neuT mice. (
The same picture prevailed when peripheral blood lymphocytes (PBLs) were analyzed from the same mice at 23 weeks of age. The number of responders with DNA-EP alone dropped steeply whereas the groups that received either Bak1-1 or Casp8 siRNA or both had mean responder values of 0.25–0.4%. For TNF-α, however, a significant enhancement was seen only with the Bak1-1 siRNA at week 16. The values observed overall for TNF-α were lower than those seen with IFN-γ (Fig. 4B; mean, 0.2%). At week 23, cell-mediated immunity dropped across all groups, but that was most likely attributable to the advanced tumor status of the mice. This resulted in lower measurable reactivity in ICS. The differences between DNA-EP alone and the groups that received either or both siRNAs were statistically significant. Data from the group that was immunized with DNA-EP and Bak1-1 at physically separate sites confirmed that the incremental effect on cell-mediated action was due to the DNA-EP and siRNA combination.
Humoral response
Antibodies to p185, the product of the rat HER2/neu oncogene, were measured by ELISA. Figure 5 shows the titer of IgG1 and IgG2a antibodies against rat HER2/neu protein elicited by DNA-EP and its combination with siRNA. Two weeks after the second DNA-EP vaccination, mice treated with Bak1-1 or Casp8 siRNA showed antibodies with a significant isotype switch to IgG2a subclass. Mice treated with DNA-EP and nonspecific siRNA did not show this isotype switch. This dominance of the IgG2a subclass suggested that the humoral response was Th1 (helper T cell type 1) skewed.

Antibody titers as measured by sandwich ELISA show a helper T cell type 1 (Th1)-skewed response. DNA-EP injections with antiapoptotic siRNAs show a dominant IgG2a component that is a characteristic feature of tumor-free BALB-neuT mice. Columns show the titers of IgG1 and IgG2a subtypes in serum at week 18. Displayed data are characteristic of three independent experiments, and sera from 10 mice were measured. Asterisks indicate that differences in data are statistically significant.
Discussion
In this study, we have evaluated the adjuvant properties of naked siRNA duplexes in an aggressive mammary tumor model (BALB-neuT). The data obtained clearly demonstrate that two of the duplexes, Bak1-1 and Casp8, are capable of exerting a strong effect in combination with DNA vaccination, leading to an enhanced CD8+ T cell response, a Th1-skewed antibody response, and increased therapeutic effect.
Most of the siRNA sequences used in this study target genes of the apoptosis pathway and induce efficacious immune responses according to published data. The rationale behind targeting these genes is that delaying apoptosis of antigen-presenting cells (APCs) or dendritic cells (DCs) could prolong antigen stimulation of T and/or B cells. With our vaccination platform, which uses plasmid DNA electroporation into the muscle, the expectation is that siRNAs targeting cell death genes or suppressors of the immune reaction could transfect APCs or muscle cells moving through the injection site (Pardoll and Beckerleg, 1995). Our initial results confirmed the ability of Bak1-1 siRNA to enhance the immunogenicity of a DNA vaccine, pV1J-neuECD.TM, in a xenogeneic vaccination protocol in BALB/c mice (Fig. 1B). We then tested this and other siRNAs in BALB-neuT mice to measure antitumor effects.
The antibody subclass switch, that is, the domination of IgG2a over IgG1 (Fig. 5), strongly indicated that a significant immunologic shift of an effector arm of the B cell response was being influenced by siRNA. A similar IgG2a bias has been observed previously for TLR9 agonists, CpG oligonucleotides, in BALB/c mice (Weeratna et al., 2005) and for a TLR7 agonist (Dharmapuri et al., 2008).
Evidence of clear inhibition of tumor growth was obtained in the BALB-neuT model (Fig. 2). siRNA duplexes acted in combination with a DNA-EP vaccine expressing rat neu ECD.TM and the results were statistically significant (Table 1). The siRNAs targeted Bak1, Bax, Casp8, Fas, and SOCS1. Four different siRNA duplexes targeting Bak1 were also tested. Of them, two siRNAs, Bak1-1 and Casp8, showed the best adjuvant activity. DNA-EP with Bak1-1 siRNA alone or as part of a Bak siRNA pool, significantly extended tumor-free survival, delayed the time to advanced disease, and reduced the number of tumors at 26 weeks compared with DNA alone. DNA-EP plus Casp8 siRNA moderately extended tumor-free survival and significantly delayed advanced disease but did not significantly reduce tumor counts at 26 weeks. The molar ratio of each siRNA to DNA was unchanged when two duplexes (Bak1-1 and Casp8) were used, but this did not result in a significant increase over the efficacy of the individual siRNAs. It is interesting to note that for the two different targets, tumor protection in general was enhanced but there are differences in the measurements of the various parameters. This (and control experiments with nonspecific siRNA) clearly suggested that this adjuvant activity was gene specific and not a general effect observed with siRNAs targeting apoptotic genes. Figure 3 depicts this and also describes the differences between the siRNAs that target Bak1. Figure 1A provides supporting in vitro evidence of target-specific inhibition by siRNA. Three of the Bak1 siRNAs showed distinct differences in their silencing efficacy, suggesting that this could be sequence dependent even for a single-gene target.
In addition, the noteworthy feature of all the experiments in the BALB-neuT model is that mice were vaccinated at 13 and 14 weeks of age, when mice display advanced atypical hyperplasia and nonpalpable multiple carcinoma in situ. These results demonstrate for the first time the efficacy of a vaccination protocol when combined with siRNA in BALB-neuT mice with spontaneous and physiologically complex tumors. We showed the impact of repeated DNA-EP vaccination in this therapeutic window (Cipriani et al., 2008). siRNA as an adjuvant now provides an additional tool to advance this methodology by improving both the antitumor and immune responses with just two DNA-EP immunizations.
The CD8+ T cell response to rHER2ECD.TM when delivered as plasmid DNA is low as shown earlier (Rovero et al., 2000; Quaglino et al., 2004; Cipriani et al., 2008). The range of the T cell response in BALB-neuT mice (Fig. 4A and B) was not significantly enhanced, but Bak1-1 and Casp8 siRNAs contributed to an increase in the number of responders. Increased secretion of cytokines IFN-γ and TNF-α was observed when the two siRNAs were used. This showed that these siRNAs were most likely prolonging the APC life span, enhancing presentation of self-antigen epitopes (HER2/neu in this case), and positively impacting interaction of APCs or muscle DCs with components of the immune system. It has been shown that CD8+ T cells can be activated by a 100-fold lower number of peptide–MHC complexes when compared with CD4+ T cells (Demotz et al., 1990). The prolonged life span could mimic an increase in the apparent epitope density on the APC surface and/or the number of complexes that promote proliferation (Rolla et al., 2006). This could play a role in recovering and selecting antigen-specific T cells within the T cell repertoire in a tolerant setting. Evidence of the antibody class switch was known earlier and here we have presented data that differentiate clearly that the immune reaction is more Th1 skewed when siRNA is coadministered with DNA-EP immunizations.
One of the drawbacks of cancer vaccines developed in mouse models in general has been their inability to elicit immune and antitumor responses in humans. The most likely causes are that in cancer patients tumors are more sophisticated than their murine counterparts and refractory to immunotherapy and chemotherapy. As patients already have detectable tumors at the time of immunization, a therapeutic regimen incorporating multiple vaccinations and long-term adjuvant treatment is a possible remedy. Increasingly, immune response modifiers are being screened and recruited for use in conjunction with DNA vaccines. In this study, we suggest that siRNAs are a class of adjuvants that could in the near future provide a substantial boost to obtaining positive results in clinical trials.
Footnotes
Acknowledgments
The authors acknowledge Dr.Guido Forni for BALB-neuT mice, and Drs. C. Bentsen, I. Pak, A. Fridman, and A. Bagchi for statistical analysis.
Author Disclosure Statement
No competing financial interests exist.