
Abstract
Extension of in vivo nucleic acid transfection techniques and increased information about those transfection properties and side effects are urgently needed to advance biological research and drug therapy. Tissue pressure-mediated transfection, involving lightly pressing the target tissue after intravenous injection of plasmid DNA or small-interfering RNA (siRNA), is a promising approach because of its high transfection efficiency and resulting low tissue damage. In this study, the gene expression/silencing properties and proinflammatory cytokine production associated with tissue pressure-mediated transfection were evaluated to extend its application. We have found that tissue pressure-mediated transfection can be applied to plasmid DNA and siRNA transfection to the spleen and siRNA transfection to the liver. In addition, we have demonstrated that these methods induce little production of proinflammatory cytokines, including tumor necrosis factor-α, interleukin (IL)-6, IL-12, and interferon-γ. Moreover, we succeeded in controlling and quantifying the degree of pressure on the spleen and kidney and found that 0.59 N/cm2 is sufficient for efficient and highly reproducible plasmid DNA transfection to the spleen and kidney in mice. Tissue pressure-mediated transfection of the kidney, liver, and spleen exhibits well-balanced characteristics including (1) simple and convenient manipulation, (2) tissue-specific, effective broad transfection properties, and (3) a low inflammatory response. Therefore, this information could be useful for a molecular-level mechanism analysis of diseases at an individual level in mammals, exploration of therapeutic target molecules and evaluation of gene therapy and nucleic acid-based therapy approaches, as well as potential clinical applications.
Introduction
Various recombinant virus and synthetic carrier-mediated transfection methods have been reported (Brown et al., 2001; Kay et al., 2001); however, the transfection strategy with naked plasmid DNA or siRNA has the advantages of convenient preparation and handling methods and a lack of toxicity associated with the transfection agents. The simplest approach is direct injection of plasmid DNA or siRNA solution into local tissues, whereby transfection is limited to the injection site (Hickman et al., 1994; Thanaketpaisarn et al., 2005). Transfection via blood flow is superior to direct injection from the point of view of transfection to many cells in the target tissue. Although normal intravenous injection of plasmid DNA or siRNA solution produces a low transfection efficiency (Hashida et al., 2001), application of physical forces to tissues, such as hydrodynamic (Liu et al., 1999; Zhang et al., 1999) and electric (Liu and Huang, 2002a; Sakai et al., 2005) forces, markedly enhances transfection efficacy. Regrettably, these methods have one crucial disadvantage, namely, they can cause severe tissue injury (Lefesvre et al., 2002; Liu and Huang, 2002b).
In their pioneering report, Liu and Huang (2002b) described that liver-specific gene expression could be obtained without liver toxicity by a simple and convenient method involving massaging (i.e., pressing multiple times) the abdomen after intravenous injection of plasmid DNA and, in addition, that administration of plasmid DNA encoding hepatocyte growth factor prevented hepatic failure. We have shown that this unique phenomenon also applies to siRNA as well as plasmid DNA delivery to the kidney, namely, that siRNA and plasmid DNA could be transfected by pressing the kidney after intravenous injection of siRNA and plasmid DNA (renal pressure-mediated transfection method), without marked renal dysfunction (Mukai et al., 2008). Because these tissue pressure-mediated transfection methods possess useful characteristics as experimental and clinical tools, there is a need for systematic investigation of their extended application to siRNA, which is both a convenient biological research tool and an important drug candidate (Dorsett and Tuschl, 2004; Kawakami and Hashida, 2007), as well as to plasmid DNA delivery to a variety of tissues. Proinflammatory cytokine production following plasmid DNA and siRNA transfection was reported to affect therapeutic effects (Kleinman et al., 2008; Robbins et al., 2008) and gene expression (Li et al., 1999). However, there is no report evaluating proinflammatory cytokine production subsequent to tissue pressure-mediated transfection.
Taking these factors into consideration, we focused on assembling information about the tissue pressure-mediated transfection method in this study: (1) exploring the extendability of this method, (2) controlling the tissue pressure, and (3) evaluating proinflammatory cytokine production. We investigated the gene expression and silencing properties of plasmid DNA and siRNA delivered by tissue pressure-mediated transfection of kidney, liver, spleen, lung, heart, muscle, and tumor, using the luciferase gene as the reporter because it is simple to handle and can be detected with high sensitivity. In addition, the proinflammatory cytokine concentration in tissue lysate as well as serum, including tumor necrosis factor (TNF)-α, interleukin (IL)-6, IL-12, and interferon (IFN)-γ, was evaluated.
Materials and Methods
Plasmid DNA
The previously constructed cytomegalovirus (CMV) immediate-early promoter-driven plasmid encoding complementary luciferase DNA (pCMV-Luc) (Kawakami et al., 2000) and a human ferritin composite promoter-driven plasmid encoding complementary luciferase DNA (pVIVO2-Luc) (Mukai et al., 2008) were used. pCMV-Luc and pVIVO2-Luc were amplified in Escherichia coli DH5α (Toyobo, Osaka, Japan), isolated, and purified with an Endofree plasmid giga kit (Qiagen, Hilden, Germany) or a JETSTAR2.0 plasmid giga kit (GenoMed, Löhne, Germany). The endotoxin in plasmid DNA solution was removed by repeated extractions with Triton X-114, including mixing 1 vol% of Triton X-114 (Nacalai Tesque, Kyoto, Japan) with the plasmid DNA solution containing endotoxin, cooling for 15 min on ice, heating for 15 min at 56°C, and separating the supernatant by centrifugation at 500 × g for 20 min at a temperature exceeding 25°C.
siRNAs
siRNAs were chemically synthesized by Gene Design (Osaka, Japan) or Hayashi Kasei (Osaka, Japan). The siRNA sequence targeting pGL3 firefly luciferase corresponded to the coding region 433–453 (5′-CTTACGCTGAGTACTTCGAAA-3′). The sense and antisense strands of the siRNA were 5′-CUUACGCUGAGUACUUCGAdTdT-3′ (sense) and 5′-UCGAAGUACUCAGCGUAAGdTdT-3′ (antisense). The sense and antisense strands of the scrambled siRNA were 5′-CUUACGCUGUCAUGAUCGAdTdT-3′ (sense) and 5′-UCGAUCAUGACAGCGUAAGdTdT-3′ (antisense).
Cell line
Murine colon 26 adenocarcinoma (Colon 26) cells were obtained from the Cancer Chemotherapy Center of the Japanese Foundation for Cancer Research (Tokyo, Japan) and cultured in RPMI 1640 medium (Nissui Pharmaceutical, Tokyo, Japan) supplemented with 10% heat-inactivated fetal bovine serum, 0.15% NaHCO3, penicillin (100 IU/ml), streptomycin (100 μg/ml), and 2 mM
Animals
Female ICR mice (5 weeks old) and female BALB/c mice (4 weeks old) were purchased from the Shizuoka Agricultural Cooperative Association for Laboratory Animals (Shizuoka, Japan). Tumor-implanted mice were obtained by subcutaneous injection of 3 × 105 Colon 26 cells in 100 μl of Hanks' balanced salt solution (Nissui Pharmaceutical) into the left back of BALB/c mice and were used when the tumors had reached diameters of 7–9 mm after approximately 1 week. All animal experiments were carried out in accordance with the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by the U.S. National Institutes of Health (Bethesda, MD) and the Guideline for Animal Experiments of Kyoto University (Kyoto, Japan).
Plasmid DNA and siRNA transfection by tissue pressure-mediated transfection
In a typical case of tissue pressure-mediated transfection, 100 μg of pCMV-Luc in 200 μl of saline was intravenously injected into anesthetized mice and the target tissue was immediately pressed. In the case of the kidney and spleen, the right kidney and spleen were exposed by a midline incision, held between the thumb and index finger, and then lightly pressed; afterward the abdomen was sutured. In the case of the liver, lung, and heart, the abdomen and chest were held between the thumb and the index and middle fingers of both hands and then pressed without exposure of the tissue. In the case of muscle and tumor, the gastrocnemius muscle of the right hind leg and tumor implanted in the left back were held between the thumb and index finger; pressure was then exerted over the skin.
In the case of direct splenic injection and the combination method of direct splenic injection and electroporation (control experiments), the modified method described previously was used (Thanaketpaisarn et al., 2005). pCMV-Luc in 30 μl of saline was injected directly into the spleen. The parameters of the electric pulses included 5 msec/pulse, 12 pulses, 2 Hz, and 130 V. The electrode gap was 2 mm.
For the siRNA transfection experiment, mice were intravenously coinjected with 100 μg of pCMV-Luc and luciferase-targeting siRNA or scrambled siRNA, immediately followed by pressing the abdomen (liver) or spleen. As a control experiment, 30 μg (liver) or 10 μg (spleen) of luciferase-targeting siRNA was intravenously injected 1 hr after administration of 100 μg of pCMV-Luc by hepatic or splenic pressure-mediated transfection.
Luciferase assay
The luciferase assay was performed as described previously (Kawakami et al., 2000). Relative light units were normalized to milligrams of protein and converted to nanograms, using a standard curve generated with a luciferase enzyme standard (Toyo Ink, Tokyo, Japan).
Imaging of luciferase activity
Anesthetized mice were injected intraperitoneally with
Controlling and quantifying the degree of pressure against spleen and kidney
The pressure control device shown in Fig. 1A was made by setting the spring (UY4-30; Misumi Group, Tokyo, Japan; NO15; Suyama Spring Mfg., Tokyo, Japan) in the barrel of the syringe (Terumo, Tokyo, Japan) and attaching a polypropylene cap with an area of 1 cm2 (low-pressure chromatography fitting 731-8221; Bio-Rad Laboratories, Hercules, CA) at the tip of the syringe. When using this device, we held the tissue between the cap of the device and a spatula and then pushed the plunger against the spatula as shown in Fig. 1B. The pressure value was estimated from the measured displacement of the spring and its force constant. Also, the device was calibrated, using an electronic balance, before experimentation.
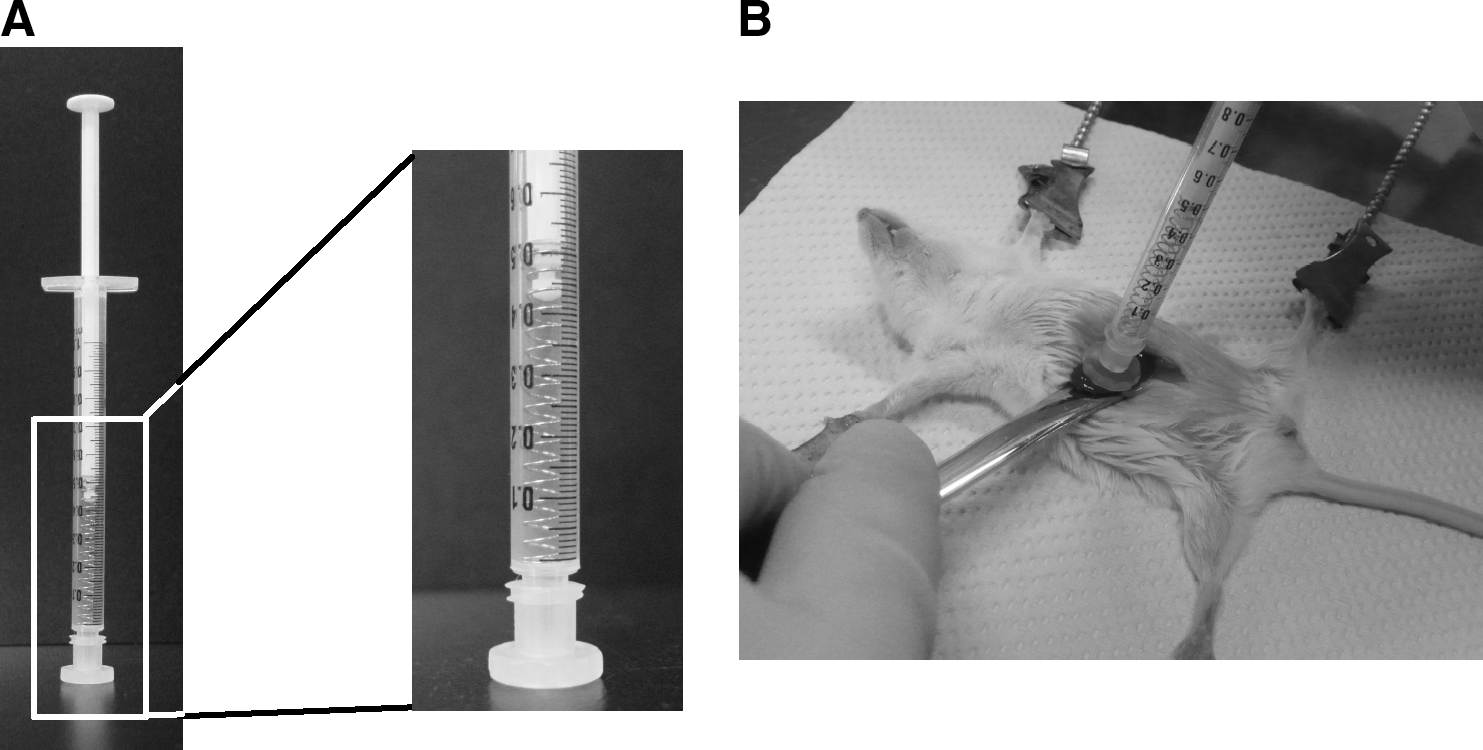
Method of controlling and quantifying the degree of pressure. (
Measurement of proinflammatory cytokine concentrations in serum and tissue lysate
Blood was collected from the inferior vena cava and then allowed to coagulate overnight at 4°C. Serum was isolated as the supernatant fraction after centrifugation at 10,000 × g for 10 min. Serum samples were immediately stored at −80°C until enzyme-linked immunosorbent assay (ELISA) was carried out. Tissues were excised after blood removal by slow saline injection from the left ventricle of the heart, washed twice in ice-cold saline, and stored at −80°C until ELISA was carried out. The tissues were then homogenized in 1 ml of PBS containing a protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO) and phenylmethylsulfonyl fluoride (100 mg/ml) (Sigma-Aldrich), using an Omni tissue homogenizer (Omni International, Marietta, GA). The supernatant fraction after centrifugation at 10,000 × g for 10 min at 4°C was immediately examined. TNF-α, IL-6, IL-12, and IFN-γ concentrations in serum and tissue lysate were determined by ELISA Ready-SET-Go! (eBioscience, San Diego, CA) for serum and a Quantikine kit (R&D Systems, Minneapolis, MN) for tissue lysate. Cationic liposome-mediated plasmid DNA transfection was carried out as a positive control. N-[1-(2,3-Dioleyloxy)propyl]-N,N,N-trimethylammonium chloride (DOTMA)-cholesterol liposomes (molar ratio, 1:1) were prepared as reported (Kawakami et al., 2000). Lipoplex was prepared by adding an equal volume of cationic liposomes to pCMV-Luc in 5% dextrose at a charge ratio (−/+) of 1.0:3.1 and at a pCMV-Luc dose of 30 μg and then stored at room temperature for 30 min. Mice were intravenously injected with prepared lipoplex.
Measurement of alanine transaminase concentration in serum
Thirty micrograms of siRNA targeting luciferase was transfected into anesthetized mice by the hepatic pressure-mediated transfection method. Blood was collected 3, 6, 12, and 24 hr after administration and serum was isolated as described previously. The alanine transaminase concentration was measured with a Transaminase C II test kit (Wako Pure Chemical Industries, Osaka, Japan).
Statistical analysis
GraphPad InStat 3.0 (GraphPad Software, San Diego, CA) was used. Statistical significance was determined by paired or unpaired Student t test for two groups. Analysis of variance (ANOVA) was performed for multiple comparisons among different groups, followed by the Tukey-Kramer test. All p values are two-tailed and p < 0.05 was considered to be indicative of statistical significance.
Results
Investigation of extended application of tissue pressure-mediated transfection method to deliver plasmid DNA to various tissues
We first examined luciferase gene expression of pressed tissue by luciferase assay 6 hr after mice had received an intravenous injection of 100 μg of pCMV-Luc, immediately followed by lightly pressing the tissues, including right kidney, liver, spleen, heart, lung, muscle, and tumor, in order to investigate the extended application of the tissue pressure-mediated transfection method in plasmid DNA transfection to various tissues. Pressing the liver and the lung and heart was replaced by pressing the abdomen and chest, respectively, as described in Materials and Methods. In this experiment, the tissue was pressed four times (1 sec each time), which is the optimal condition reported by Liu and Huang in the case of liver (Liu and Huang, 2002b). As shown in Fig. 2, high luciferase gene expression of the pressed tissues was obtained by pressing the right kidney, liver, and spleen; the levels were 4.7, 3.2, and 0.15 ng/mg protein, respectively. This level of gene expression in the kidney and liver corresponds to that obtained in previous reports (Liu and Huang, 2002b; Mukai et al., 2008). In contrast, when pressing the other tissues, luciferase levels of the pressed tissues were less than 4 × 10−4 ng/mg protein (Fig. 2). Also, in the case of only intravenous administration of pCMV-Luc without pressing, little luciferase activity was expressed in any of the tissues assayed (data not shown), in agreement with previous findings (Li and Huang, 1997; Kawakami et al., 2000).

Investigation of extended application of the tissue pressure-mediated transfection method in plasmid DNA delivery to various tissues (including right kidney, liver, spleen, lung, heart, muscle, and tumor). Shown are the luciferase levels of pressed tissue 6 hr after intravenous injection of 100 μg of pCMV-Luc, immediately followed by pressing the target tissue four times (1 sec each). Each value represents the mean and SD (n = 3 [spleen and tumor], 4 [liver, lung, heart, and muscle], or 7 [kidney]).
Effect of pressure conditions, including repetition and duration of each pressing, on gene expression level
To determine the optimal pressure conditions, specifically the repetition and duration of pressure on splenic and renal tissue, the luciferase level of pressed spleen and right kidney at 6 hr was estimated when pressure was applied from one to four times for a period of 1 or 3 sec (for each pressing) after injection of 100 μg of pCMV-Luc. In both cases, the luciferase levels were nearly constant under all these conditions (Fig. 3A and B). Therefore, the following experiments were performed with the tissue being pressed once for 1 sec (splenic and renal transfection) or four times for 1 sec each (hepatic transfection).
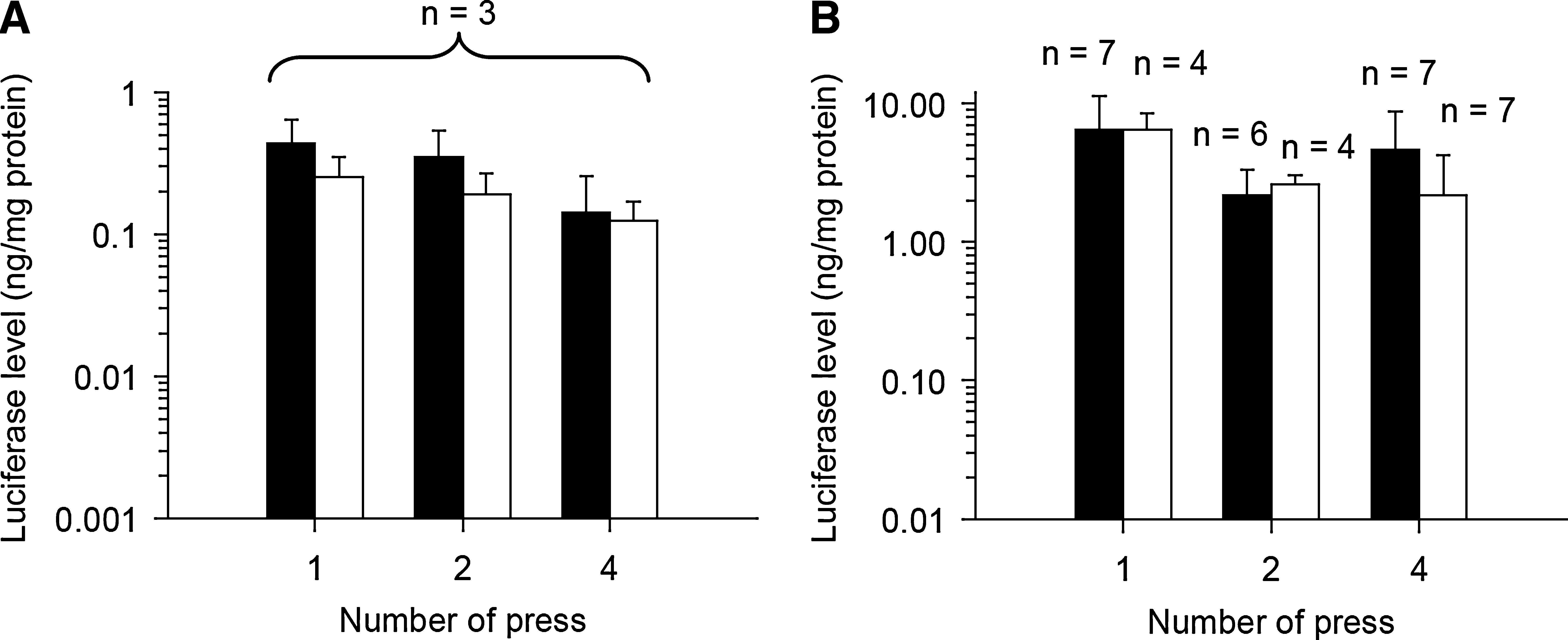
Effect of pressure conditions, including the number and duration of each pressing, on gene expression level. Shown are luciferase levels of pressed (
Spleen-specific plasmid DNA transfection by splenic pressure-mediated transfection
To investigate the gene expression properties associated with plasmid DNA delivery by the splenic pressure-mediated transfection method, luciferase gene expression was estimated by luciferase assay and imaging 6 hr after administration of 100 μg of pCMV-Luc. As shown in 4A–C, luciferase gene expression was specifically obtained in the spleen. The luciferase level in the spleen was about 0.45 ng/mg protein and the luciferase levels in the other organs, including liver, lung, heart, and kidneys, were less than 4 × 10−5 ng/mg protein (Fig. 4A). In addition, when pVIVO2-Luc was alternatively administered to confirm the lack of promoter dependence, luciferase was also specifically expressed in the spleen and its level was 0.18 ± 0.04 ng/mg protein. Also, to examine the efficiency of the splenic pressure-mediated transfection method, direct splenic injection was performed and a combination method with electroporation that has been used for gene immunotherapeutic studies (Maloy et al., 2001; Ho et al., 2006) was carried out as a control. The luciferase level achieved by the splenic pressure-mediated transfection method was almost equal to those produced by direct splenic injection and the combination method with electroporation (Fig. 4D). Moreover, to confirm broad transfection to the spleen by the splenic pressure-mediated transfection method, isolated spleen 6 hr after pCMV-Luc administration was separated into two parts (upper and lower) and the luciferase level in each part was measured. There was no difference between luciferase levels of the two parts (Fig. 4E, left). In contrast, when pCMV-Luc was directly injected into the lower pole of the spleen, the luciferase level in the lower part of the spleen was significantly (about 10-fold) higher than that in the upper part (Fig. 4E, right).
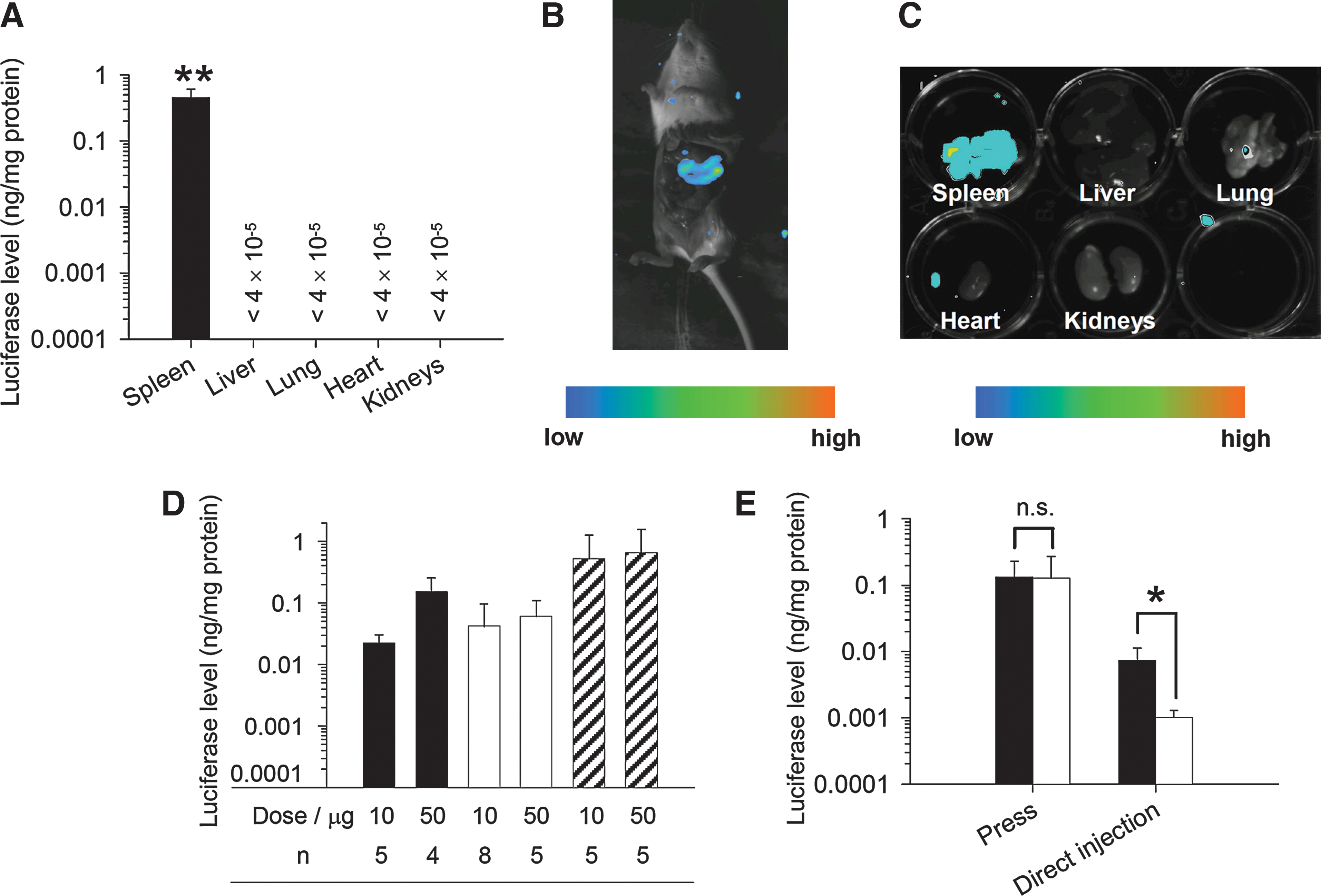
Spleen-specific plasmid DNA transfection by the splenic pressure-mediated transfection method. Shown are (
siRNA delivery to liver and spleen by hepatic and splenic pressure-mediated transfection
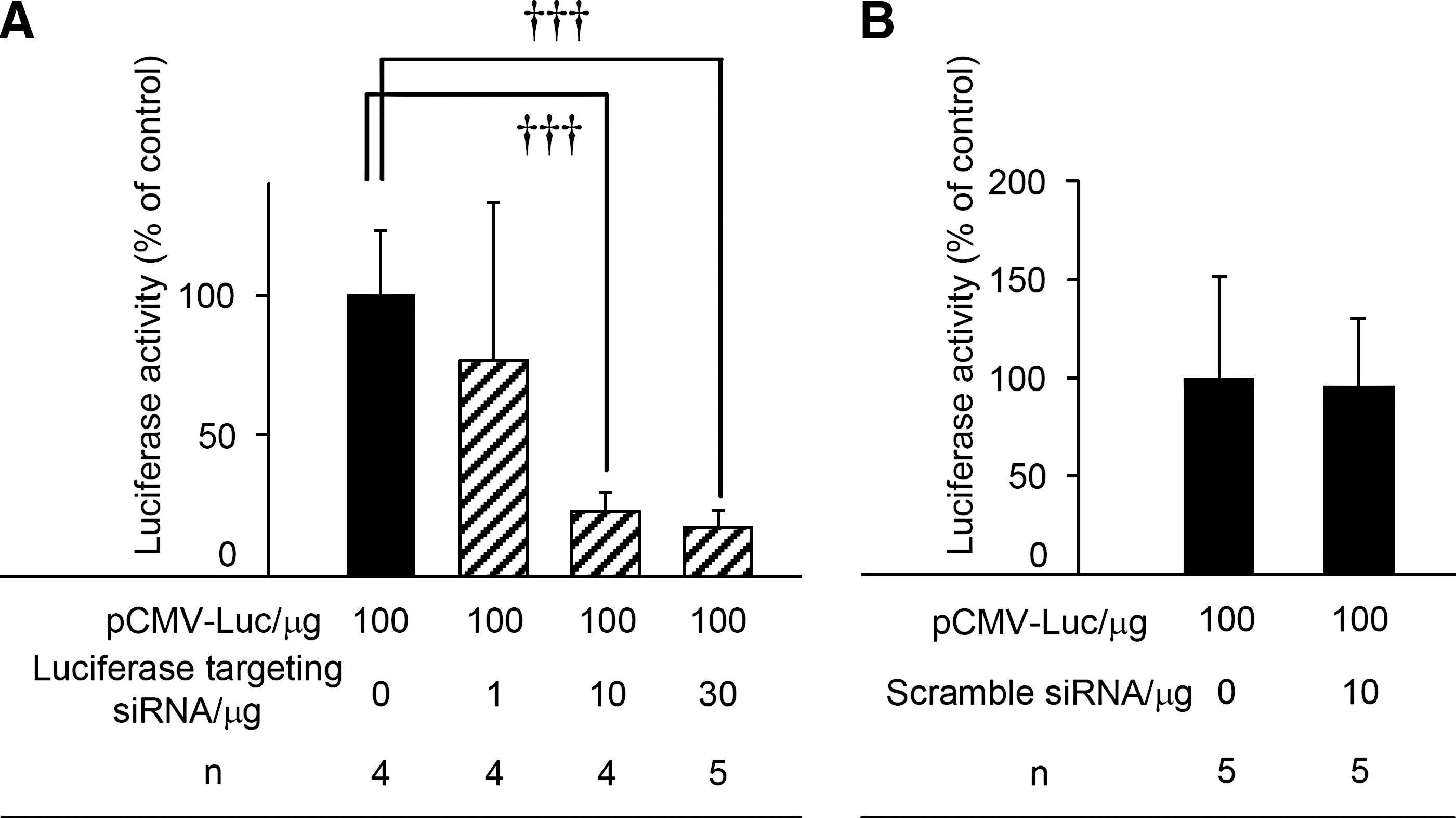
To investigate the application of this method to hepatic and splenic siRNA transfection, gene silencing after siRNA administration by the hepatic or splenic pressure-mediated transfection method was estimated on the basis of luciferase expression, because the assay system is simple to carry out and quantify. When coadministration of pCMV-Luc and luciferase-targeting siRNA was carried out by the hepatic or splenic pressure-mediated transfection method, the luciferase level in the liver and spleen was siRNA dose-dependently suppressed, compared with only pCMV-Luc administration (Fig. 5A and C, and Fig. 6A). When the dose of siRNA was more than 30 μg (liver) and 10 μg (spleen), about 80% suppression was observed. In contrast, after scrambled siRNA administration, no marked suppression of luciferase gene expression was observed in either case (Figs. 5B and 6B). Also, when luciferase-targeting siRNA was given intravenously 1 hr after pCMV-Luc administration by the hepatic or splenic pressure-mediated transfection method to confirm that siRNA is not transfected only by intravenous injection, the same level of luciferase activity was obtained after 6 hr compared with the administration of only pCMV-Luc (data not shown).

siRNA delivery to the liver by the hepatic pressure-mediated transfection method. Shown are luciferase levels 6 hr after coadministration of pCMV-Luc and (
siRNA delivery to the spleen by the splenic pressure-mediated transfection method. Shown are luciferase levels 6 hr after coadministration of pCMV-Luc and (
Effect of degree of pressure on gene expression level
Next, we tried to control and quantify the degree of pressure, using a simple homemade device described in Materials and Methods, and to examine the effect of the degree of pressure on the gene expression level after administration of 100 μg of pCMV-Luc by the splenic and renal pressure-mediated transfection methods, which are easy to perform (the pressure can be well controlled because of the size and shape of the tissues). As shown in Fig. 7A and B, the luciferase level in pressed spleen and right kidney increased in a pressure-dependent manner to 0.59 N/cm2 and then saturated. The saturated levels produced by splenic and renal pressure-mediated transfection are approximately 0.3 and 5 ng/mg protein, respectively, corresponding to results produced by pressing with the fingers (Figs. 2 –4).
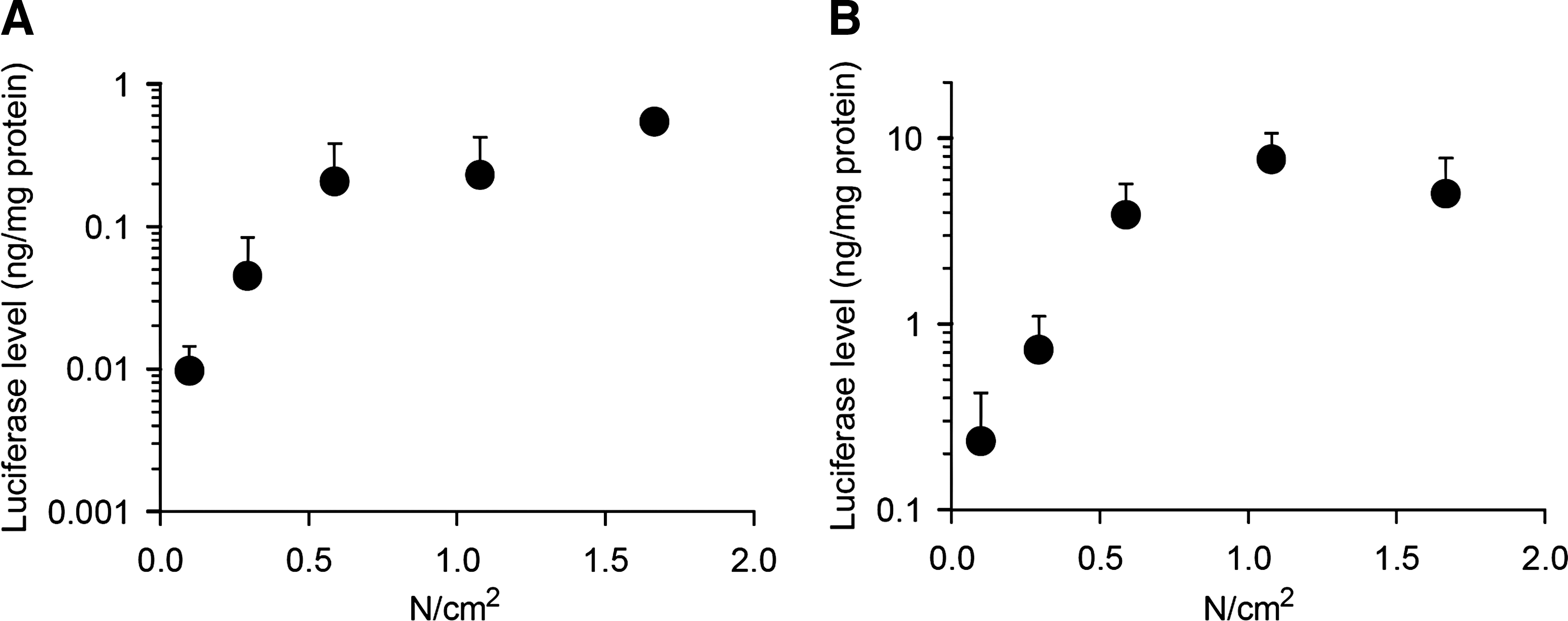
Effect of the degree of pressure on gene expression level. Shown are luciferase levels in (
Evaluation of proinflammatory cytokine production by tissue pressure-mediated transfection
To evaluate proinflammatory cytokine production by tissue pressure-mediated transfection, we evaluated the TNF-α, IL-6, IL-12, and IFN-γ concentrations in serum in a time-dependent manner after 100 μg of pCMV-Luc was administered by renal, hepatic, and splenic pressure-mediated transfection. In the case of renal and splenic pressure-mediated transfection, the right kidney and spleen were pressed at 0.59 N/cm2, using the pressure control device described previously. Cationic liposome-mediated transfection was performed as a positive control. As shown in Fig. 8, TNF-α, IL-6, IL-12, and IFN-γ in serum exhibited no striking changes, compared with the no-treatment group, at 3, 6, and 12 hr, although the IL-6 level was slightly increased in the case of renal plasmid DNA transfection. Moreover, to obtain more direct evidence of only minor proinflammatory cytokine production by tissue pressure-mediated transfection, proinflammatory cytokine concentrations in pressed tissue lysate including right kidney, left lobe of liver, and spleen were measured at 3 hr. TNF-α, IL-6, IL-12, and IFN-γ concentrations also exhibited no marked changes, compared with the no-treatment group (Fig. 9).

Evaluation of proinflammatory cytokine concentrations in serum induced by tissue pressure-mediated transfection. (

Evaluation of proinflammatory cytokine concentrations in tissue lysate induced by tissue pressure-mediated transfection methods. Shown are (
Discussion
Today, we can routinely transfect a variety of cells with nucleic acids, including plasmid DNA and siRNA, using commercially available in vitro transfection reagents. However, in the case of in vivo applications, the nucleic acid transfection technique is restricted and, therefore, extension of the in vivo nucleic acid transfection techniques and information about their transfection properties and side effects are urgently needed in order to advance biological research and drug therapy development.
As shown in Figs. 2 and 4, we demonstrated that plasmid DNA can be transfected to the spleen by the splenic pressure-mediated transfection method, which involves pressing the spleen after intravenous injection of plasmid DNA. Gene expression was tissue specific and widely distributed in the pressed spleen (Fig. 4), in agreement with previous reports of hepatic and renal plasmid DNA transfection (Liu and Huang, 2002b; Mukai et al., 2008). We then compared the gene expression level in the spleen by this method (0.45 ng/mg protein) with that of other splenic transfection methods, including direct splenic injection and the combination method of direct splenic injection and electroporation, using a CMV early-immediate promoter-driven luciferase gene as the reporter in order to evaluate the efficiency of the splenic pressure-mediated transfection method. The luciferase level achieved by the splenic pressure-mediated transfection method was almost equal to that of the direct splenic injection and a combination with electroporation, which have been used for many gene immunotherapeutic studies (Maloy et al., 2001; Ho et al., 2006) (Fig. 4D). Also, recombinant adenovirus, which is the most representative viral vector, was reported to accumulate and provide a high level of gene expression in the spleen after intravenous injection. The gene expression level was about 1 ng/mg protein (Sakurai et al., 2007) and almost equal to that obtained by the splenic pressure-mediated transfection method. In addition, gene expression could be widely obtained in the spleen by this method, although gene expression by direct splenic injection was limited to the vicinity of the injection site (Fig. 4E). These results led us to believe that splenic pressure-mediated transfection produces broad, spleen-specific gene expression comparable to that achieved by the other methods described previously.
The spleen, one of the secondary lymphoid organs, contains abundant immunocompetent cells, including dendritic cells, macrophages, T cells, and B cells, and antigen presentation to lymphocytes and differentiation to mature lymphocytes take place in the spleen (Mebius and Kraal, 2005). Therefore, the spleen is one of the most important target organs for DNA vaccination and cytokine gene therapy. The clinical results of DNA vaccination in HIV-1- and malaria-infected patients show that conventional intradermal and intramuscular administration are insufficient to produce a therapeutic effect (Calarota et al., 1998; Wang et al., 1998) and, therefore, exploration of more efficient administration routes, including the spleen-targeting method, has been carried out. For example, Maloy and colleagues reported that direct splenic injection produces a 100-fold induction of the cytotoxic T lymphocyte response, compared with intradermal and intramuscular injection (Maloy et al., 2001). Similarly, it has been reported that a combination of direct splenic injection and electroporation is superior to muscular electroporation for IL-4 gene therapy of rat experimental allergic encephalomyelitis (Ho et al., 2006). Judging from the gene expression properties discussed previously, the spleen pressure-mediated transfection method may be useful for DNA vaccination and cytokine gene therapy.
siRNA is a convenient biological research tool and simultaneously an important drug candidate because of its sequence-specific suppression of target gene expression (Dorsett and Tuschl, 2004). We previously reported that siRNA may be transfected to the kidney by the renal pressure-mediated transfection method (Mukai et al., 2008). Pressure-mediated transfection methods in various tissues are expected to be suitable for routine screening using siRNA-mediated knockdown under in vivo conditions. Here, we showed that these tissue pressure-mediated transfection methods can be applied in siRNA transfection to the liver and spleen (Figs. 5 and 6). This is the first reported case of spleen-targeting siRNA transfection. As for hepatic siRNA transfection, many investigations have been carried out using the hydrodynamics-based method (Herweijer and Wolff, 2007); however, severe liver toxicity in mice is a major problem (Liu and Huang, 2002b). In contrast, the hepatic pressure-mediated transfection method could transfect siRNA to the liver without increasing the serum alanine transaminase level, which is an indicator of liver toxicity (data not shown). This result corresponds to the hepatic plasmid DNA transfection reported by Liu and Huang (2002b). For siRNA transfection to the liver, therefore, the hepatic pressure-mediated transfection method is an alternative to the hydrodynamics-based method.
Proinflammatory cytokine production is an important side effect caused by the in vivo nucleic acid transfection method (Li et al., 1999; Kleinman et al., 2008; Robbins et al., 2008). As far as intravenous cationic liposomes and polymer-mediated transfection methods as representative synthetic carriers are concerned, many groups have reported that proinflammatory cytokine production, including TNF-α, IL-6, IL-12, and IFN-γ, is induced mainly by Kupffer cell Toll-like receptor 9-mediated recognition of the CpG motif of the plasmid DNA backbone (Dow et al., 1999; Whitmore et al., 1999; Sakurai et al., 2002; Yasuda et al., 2005). This needs to be improved insofar as the relationship to liver toxicity is concerned (Tousignant et al., 2000). The recombinant adenovirus-mediated transfection method similarly induces TNF-α and IL-6 production and subsequent liver toxicity (Lieber et al., 1997). Moreover, importantly, Kleinman and colleagues showed that the angiogenesis suppression activity of siRNA targeting vascular endothelial growth factor A or its receptor VEGFR1 is derived from a nonspecific interferon response in a murine model of choroidal neovascularization (Kleinman et al., 2008). A similar result has been reported in the case of the antiviral activity of antiinfluenza siRNA (Robbins et al., 2008). In this, proinflammatory cytokine production caused by transfection is a disruptive factor for the correct analysis of diseases and for evaluation of gene therapy and nucleic acid-based therapy approaches. Here, we demonstrated that tissue pressure-mediated transfection methods induce few inflammatory responses, including TNF-α, IL-6, IL-12, and IFN-γ production (Figs. 8 and 9).
In the case of tissue pressure-mediated transfection, tissues were pressed with the fingers and, thus, the degree of pressure was not quantitatively controlled. In this study, we were successful in controlling and quantifying the degree of pressure against the spleen and kidney by means of a simple homemade device and found that 0.59 N/cm2 is sufficient for efficient and highly reproducible plasmid DNA transfection to the spleen and kidney of mice (Figs. 1 and 7). This simple device will be useful for further experiments involving pressure-mediated transfection methods.
In conclusion, we have found that tissue pressure-mediated transfection methods involving lightly pressing the target tissue after intravenous injection of plasmid DNA or siRNA can be applied to plasmid DNA and siRNA transfection of the spleen and to siRNA transfection of the liver. In addition, we demonstrated that these methods induce little proinflammatory cytokine production involving TNF-α, IL-6, IL-12, and IFN-γ. Moreover, we succeeded in controlling and quantifying the degree of pressure against the spleen and kidney and found that 0.59 N/cm2 is sufficient for efficient and highly reproducible plasmid DNA transfection of the spleen and kidney of mice. Tissue pressure-mediated transfection of the kidney, liver, and spleen offers well-balanced characteristics including (1) simple and convenient manipulation, (2) tissue-specific, high and broad transfection properties, and (3) a low inflammatory response. This information could be useful for the molecular-level mechanism analysis of diseases at an individual level in mammals, exploration of therapeutic target molecules and evaluation of gene therapy and nucleic acid-based therapy approaches, as well as clinical applications.
Footnotes
Acknowledgments
This work was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan, and by Health and Labor Sciences Research Grants for Research on Advanced Medical Technology from the Ministry of Health, Labor, and Welfare of Japan.
Author Disclosure Statement
No competing financial interests exist.