
Abstract
Recombinant adeno-associated virus (rAAV) production systems capable of meeting clinical or anticipated commercial-scale manufacturing needs have received relatively little scrutiny compared with the intense research activity afforded the in vivo and in vitro evaluation of rAAV for gene transfer. Previously we have reported a highly efficient recombinant herpes simplex virus type 1 (rHSV) complementation system for rAAV production in multiple adherent cell lines; however, production in a scalable format was not demonstrated. Here we report rAAV production by rHSV coinfection of baby hamster kidney (BHK) cells grown in suspension (sBHK cells), using two ICP27-deficient rHSV vectors, one harboring a transgene flanked by the AAV2 inverted terminal repeats and a second bearing the AAV rep2 and capX genes (where X is any rAAV serotype). The rHSV coinfection of sBHK cells produced similar rAAV1/AAT-specific yields (85,400 DNase-resistant particles [DRP]/cell) compared with coinfection of adherent HEK-293 cells (74,600 DRP/cell); however, sBHK cells permitted a 3-fold reduction in the rHSV-rep2/capX vector multiplicity of infection, grew faster than HEK-293 cells, retained specific yields (DRP/cell) at higher cell densities, and had a decreased virus production cycle. Furthermore, sBHK cells were able to produce AAV serotypes 1, 2, 5, and 8 at similar specific yields, using multiple therapeutic genes. rAAV1/AAT production in sBHK cells was scaled to 10-liter disposable bioreactors, using optimized spinner flask infection conditions, and resulted in average volumetric productivities as high as 2.4 × 1014 DRP/liter.
Introduction
The production of rAAV can be accomplished by a variety of methods, whether in suspension or in adherent cell lines. The predominant method involves transient transfection of mammalian cells with multiple plasmids. Historically, three plasmids are employed to deliver all the necessary elements for rAAV production: one plasmid contains the gene of interest flanked by the AAV inverted terminal repeats (ITRs), the minimal cis-acting sequences necessary for AAV replication and packaging; the second plasmid expresses the rep (Rep78, 68, 52, and 40 proteins) and cap (structural proteins VP1, VP2, and VP3 from any AAV serotype) genes; and the third plasmid expresses the adenovirus helper functions (E1a, E1b, E2a, E4orf6, and virus-associated RNA I genes). Multiple variations of the transient transfection method have been pursued, including plasmid transfection to deliver the AAV genes and the gene of interest followed by adenovirus infection, and infection/transfection of stable cell lines expressing either rep and cap or harboring the ITR-flanked gene of interest, and have been reviewed (Buning et al., 2008).
Despite the advantageous characteristics of rAAV and the extensive research into preclinical applications, production scale-up continues to limit rAAV use in large clinical trials that require even moderate doses of vector. Suspension cell culture can facilitate scale-up of viral vector production; however, reports of rAAV production on mammalian cells adapted to grow in suspension have been limited to transient transfection of HEK-293 cells (Park et al., 2006; Durocher et al., 2007; Feng et al., 2007; Hildinger et al., 2007) and amplification of an rAAV carrying the gene of interest on A549 packaging cells that expresses rep and cap genes by coinfection of the rAAV with adenovirus (Farson et al., 2004). In contrast, extensive work has been described using baculovirus infection of insect cells for rAAV production. Baculovirus-based suspension systems employ simultaneous infection of insect cells with three or four baculoviral expression vectors (BEVs) to deliver the ITR-flanked gene of interest and the AAV rep and cap genes (Urabe et al., 2002). Stability of BEV rAAV production systems during serial expansion of helper constructs continues to hamper its clinical applicability, despite the finding that the multiplicity of infection (MOI) input can be significantly reduced (Kohlbrenner et al., 2005; Negrete et al., 2007; Cecchini et al., 2008).
The herpes simplex virus (HSV) coinfection method of rAAV production is an alternative to the baculovirus system, transient transfection, and adenovirus infection. HSV has been widely used to complement rAAV replication and packaging, with the HSV helicase–primase complex (UL5, UL8, and UL52), DNA polymerase, and DNA-binding protein (UL29) genes providing the essential AAV helper functions (Buller et al., 1981; Weindler and Heilbronn, 1991; Alazard-Dany et al., 2009). Although wild-type HSV and several recombinant deletion mutants of HSV have been examined for rAAV production, ICP27-deleted HSV vectors have demonstrated the most promise regarding specific yields in mammalian cells (Conway et al., 1997, 1999; Zhang et al., 1999; Feudner et al., 2001; Mistry et al., 2002; Wustner et al., 2002; Booth et al., 2004; Toublanc et al., 2004; Alazard-Dany et al., 2009). We have previously reported a highly efficient rHSV-based rAAV complementation system that uses two rHSV vectors: one harboring the ITR-flanked gene of interest and the second bearing the rep and cap genes (Kang et al., 2009). The helper vectors were demonstrated to be both stable during serial helper expansion (Kang et al., 2009) and scalable to 3.5-liter packed-bed bioreactors (Knop and Harrell, 2007). The rHSV-based rAAV production platform used adherent HEK-293 cells infected at about 1 × 106 viable cells (VC)/ml in 10-stack Nunclon Δ cell factories (Thermo Fisher Scientific, Waltham, MA) at MOIs of 12 and 2 for rHSV-rep2/capX and rHSV-GOI (with GOI representing any transgene of interest), respectively, and vector was harvested at 52 hr postinfection (Kang et al., 2009). Here we present significant improvements to rAAV production by rHSV coinfection by employing baby hamster kidney cells adapted to grow in suspension (sBHK). This platform provides ease of scale-up due to suspension cell culture, abbreviated cell expansion and vector production cycles due to faster cellular growth rates and metabolism, an increase in volumetric productivity, and a decrease in total rHSV input by >50% during coinfection.
Materials and Methods
Cell culture, herpes simplex virus production, and infectious titer determination
V27, HEK-293, and C12 cells were obtained from the Powell Gene Therapy Center of the University of Florida (Gainesville, FL). Suspension baby hamster kidney (sBHK) cells were purchased from the European Collection of Cell Cultures (ECACC, Porton Down, UK) and HeLaRC32 cells were purchased from the American Type Culture Collection (ATCC, Manassas, VA). All cells were maintained in Dulbecco's modified Eagle's medium (DMEM; HyClone, South Logan, UT) containing 10% fetal bovine serum (FBS; HyClone) and either 1% (v/v) penicillin–streptomycin (Cellgro/Mediatech, Manassas, VA) for sBHK, HEK-293, and HeLaRC32 culture or Geneticin (50 μg/ml; Invitrogen) for V27 and C12 cells.
The rHSV-rep2/capX (with X representing any AAV capsid serotype) and rHSV-GOI vectors were constructed as described previously (Kang et al., 2009). The rHSV vectors were propagated on the ICP27-complementing V27 cell line (Rice and Knipe, 1990), in packed-bed bioreactors or cell factories as previously described (Knop and Harrell, 2007), and infectious vector was harvested 72 hr postinfection by recovering culture supernatant and cell-associated vector. The resulting process fluids were combined and clarified by depth filtration (1.2 μm) and absolute filtration (0.8/0.45 μm), and concentrated by hollow fiber filter (HFF) tangential flow filtration (TFF). The infectious titers (plaque-forming units [PFU]/ml) of rHSV vector stocks were determined by a modified plaque assay. Briefly, V27 cells in 24-well plates were infected with serial dilutions of rHSV. The inoculum was exchanged with culture medium containing 0.2% human γ-globulins (Sigma-Aldrich, St. Louis, MO) at 6 hr postinfection. At 3 days postinfection, the culture medium was aspirated and the cells were fixed with ice-cold methanol. The cells were then incubated with a horseradish peroxidase (HRP)-labeled anti-HSV antibody (Dako, Glostrup, Denmark) and rHSV plaques were visualized by Vector VIP staining (Vector Laboratories, Burlingame, CA) as per the manufacturer's instructions.
rHSV coinfection method
Mammalian cells were simultaneously coinfected with both rHSV vectors. At 2–4 hr postinfection infectious medium was exchanged with serum-free DMEM equivalent to the preinfection culture volume. At the time of harvest, MgCl2 was added to a final concentration of 2 mM, 10% (v/v) Triton X-100 (octyl phenol ethoxylate; J.T. Baker, Phillipsburg, NJ) was added to a final concentration of 1% (v/v), and Benzonase (EMD Chemicals, Gibbstown, NJ) was added to a final concentration of 25–50 U/ml. This mixture was stirred (25-ml spinner flasks) or rocked (disposable bioreactors) at 37°C for 2–4 hr. Before processing, 5 M NaCl was added to a final concentration of 1.0 M and the mixture was frozen at −80°C (25-ml spinner flasks) or clarified (disposable bioreactors).
Assays
DNase-resistant particles (DRP) were quantified by real-time quantitative polymerase chain reaction (qPCR) in an iCycler iQ 96-well block format thermocycler (Bio-Rad, Hercules, CA). Crude samples were incubated in the presence of DNase I (100 U/ml; Promega, Madison, WI) at 37°C for 60 min, followed by proteinase K (Invitrogen, Carlsbad, CA) digestion (10 U/ml) at 50°C for 60 min, and then denatured at 95°C for 30 min. Linearized plasmid pDC-67/+SV402 was used to generate standard curves. The primer–probe set was specific for the simian virus 40 (SV40) poly(A) sequence (rAAV-F, 5′-AGCAATAGCATCACAAATTTCACAA-3′; rAAV-R, 5′-CCAGACATGATAAGATACATTGATGAGTT-3′; rAAV-Pr, 5′-6-FAM-AGCATTTTTTTCACTGCATTCTAGTTGTGGTTTGTC-TAMRA-3′). Amplification of the PCR product was achieved with the following cycling parameters: 1 cycle at 50°C for 2 min, 1 cycle at 95°C for 10 min; 40 cycles of 95°C for 15 sec, and 60°C for 60 sec.
Infectious particle (ip) titering was performed on rAAV-GFP stocks, using a green cell assay. C12 cells (Clark et al., 1995, 1996) were infected with serial dilutions of rAAV-GFP plus saturating concentrations of human adenovirus type 5 (reference material, VR-1516; ATCC) to provide AAV replication helper functions. At 48–72 hr postinfection, the number of fluorescing green cells (each cell representing one infectious event) was counted and used to calculate the titer (ip/ml) of the virus sample.
Infectivity of rAAV particles harboring a gene of interest (rAAV-GOI) was determined in a TCID50 (tissue culture infectious dose at 50%) assay. Eight replicates of rAAV were serially diluted in the presence of human adenovirus type 5 and used to infect HeLaRC32 cells (Salvetti et al., 1998) in a 96-well plate. Three days postinfection, lysis buffer (final concentrations of 1 mM Tris-HCl [pH 8.0], 1 mM EDTA, 0.25% [w/v] deoxycholate, 0.45% [v/v] Tween 20, 0.1% [w/v] sodium dodecyl sulfate, and proteinase K at 0.3 mg/ml [Promega, Madison, WI]) was added to each well then incubated at 37°C for 1 hr, 55°C for 2 hr, and 95°C for 30 min. The lysate from each well (2.5-μl aliquot) was assayed in the DRP qPCR assay described previously. Wells with C t values lower than the value of the lowest quantity of plasmid of the standard curve were scored as positive. TCID50 infectivity per milliliter (TCID50/ml) was calculated with the Kärber equation, using the ratios of positive wells at 10-fold serial dilutions.
rHSV-specific assays for quantifying DNA and protein were executed as previously described and human α1-antitrypsin expression in C57BL/6 mice sera was quantified by enzyme-linked immunosorbent assay (ELISA), as previously described (Kang et al., 2009).
Enumeration of viable cells (VC) and nonviable cells was performed with a Guava EasyCyte mini system (Guava Technologies/Millipore, Hayward, CA) using the Guava ViaCount assay and reagents in accordance with the manufacturer's instructions.
Bioreactor production of rAAV
Disposable bioreactors were controlled with a Sartorius BIOSTAT CultiBag RM 20/50 basic control unit and platform (Sartorius Stedim Systems, Springfield, MO). Cultures were seeded at 2.5–3.5 × 105 VC/ml into disposable bioreactors in DMEM–10% (v/v) FBS, and coinfected with rHSV 24–48 hr postseeding at densities of 1.4–2.2 × 106 VC/ml. Scaled-up runs (5 or 10 liters) were initially seeded at 2 or 4 liters, diluted to 5 or 10 liters at 24 hr postseeding, respectively, and infected 24 hr later. Culture pH was maintained at 7.2 by headspace CO2 addition, dissolved oxygen (DO) was monitored until < 50% saturation, and then headspace oxygen enrichment was used to maintain the DO at 50%. Agitation was maintained at 20 rocks/min at an angle of 6.8° (5 or 10 liters) or 7.2° (1 liter). Temperature was maintained at 37°C. A total headspace gas flow rate of 0.1 volume gas/volume culture/min (vvm) was used. Data acquisition was accomplished with the Sartorius MFCS/DA software package.
Medium exchange at 2–4 hr postinfection was accomplished by centrifugation (640 × g, 4 min), with an integrated perfusion membrane (100 ml/min for volume reduction/perfusion), or via a Sartorius Sartocon Slice 1000 flat sheet TFF holder (Sartorius Stedim Systems) and 0.2- and 0.45-μm nominal pore size Hydrosart membranes (cat. nos. 3051860701W-SG and 3051860601O-SG, respectively). Flat sheet TFF employed recirculation rates of 350–500 ml/min and the transmembrane pressure (TMP) was maintained at approximately 0.5 psig. Volume was reduced 5- to 10-fold and culture was diluted back to the preinfection volume with DMEM lacking FBS.
rAAV vector purification and in vivo evaluation
Crude lysate from disposable bioreactors was clarified by depth filtration (1.2 μm) and absolute filtration (0.8/0.45 μm) and stored for 18–96 hr. Clarified lysate was concentrated and buffer was exchanged with either a Sartorius Sartocon Slice 1000 flat sheet holder using 100-kDa MWCO Hydrosart membranes (cat. no. 3051446801E-SG) or GE Healthcare Life Sciences (Piscataway, NJ) 300-kDa MWCO hollow fiber filter (HFF) devices (cat. nos. UFP-300-C-5A and UFP-300-C-9A). A recirculation rate of 0.5–1.5 liters/min was used and the TMP was maintained at 8.5 ± 1.5 psig.
Concentrated, buffer-exchanged rAAV lysate was purified by two-column chromatography. Capture chromatography employed an anion-exchange column. The anion-exchange column peak was loaded directly onto an affinity column and eluted with a pH gradient. The affinity column eluent was neutralized to pH 7.0–8.0 and buffer was exchanged to lactated Ringer's solution, using a 300-kDa MWCO hollow fiber cartridge (cat. no. UFP-300-E-MM01A; GE Healthcare Life Sciences) for TFF and diafiltration (DF).
Animal experiments with C57BL/6 mice were conducted as previously described (Kang et al., 2009). Purified rAAV1/AAT was administered to C57BL/6 mice (1 × 1011 DRP/mouse) and serum samples were collected from mice 1 month postinjection as previously described (Kang et al., 2009).
Results
BHK cell line and suspension growth
Several cell lines have been widely used for rAAV manufacture, most notably HEK-293 and HeLa cell lines. Despite receiving comparably little use, baby hamster kidney cells (clone 21) have been employed to produce rAAV via rHSV vector coinfection (Booth et al., 2004; Kang et al., 2009). The lineage and safety of mammalian-derived cell lines are important for regulatory filings in support of biotherapeutic manufacturing methods for clinical trials. BHK cells have been shown to generate (Albu and Holmes, 1973) and release (Bergmann and Wolff, 1981) endogenously produced virus-like particles (VLPs); however, the BHK cell line has been used both in a Huntington's disease phase 1 clinical trial to deliver ciliary neurotrophic factor (CNTF) (Bloch et al., 2004) and for the manufacture of recombinant factor VIII to treat hemophilia A. The latter is a U.S. Food and Drug Administration (FDA)-licensed, commercial process (Boedeker, 2001), validating the use of BHK cell-based manufacturing as a viable method for the production of human therapeutics. Therefore, BHK cells adapted to grow in suspension (sBHK) were examined as an rAAV production platform because of the high specific yield of their adherent counterpart and their favorable regulatory profile. sBHK cells were cultured in spinner flasks (80–120 rpm) in DMEM–FBS in water-jacketed incubators (5% CO2, 37°C) and initially maintained between cell densities of 2 × 105 and 1 × 106 VC/ml. The sBHK cells had an average doubling time of 12 ± 2 hr (n = 33) under these conditions, in contrast to adherent HEK-293 cells, which had a doubling time of about 24 hr in T-flasks.
rAAV production optimization: Harvest timing, cell density, and MOI
rAAV production by rHSV coinfection was initially optimized in 25-ml spinner flask cultures by examining rAAV harvest timing after coinfection, infection cell density, and infection MOI. The rHSV constructs used for coinfection production of rAAV on sBHK cells are shown in Fig. 1. sBHK cells in spinner flasks were coinfected with rHSV-rep2/cap2 (MOI of 12) and rHSV-GFP (MOI of 2). Samples were taken at the indicated intervals and the infectious titer was determined. Across the three time points examined, rAAV2/GFP specific yields of 6500–7500 ip/cell were obtained (Fig. 2A), which were similar to previous results with adherent HEK-293 and BHK cells. On the basis of these results, the harvest of rHSV-coinfected sBHK cells could be executed as early as 24 hr postinfection (Fig. 2A), or about two preinfection doubling times for the sBHK cells.
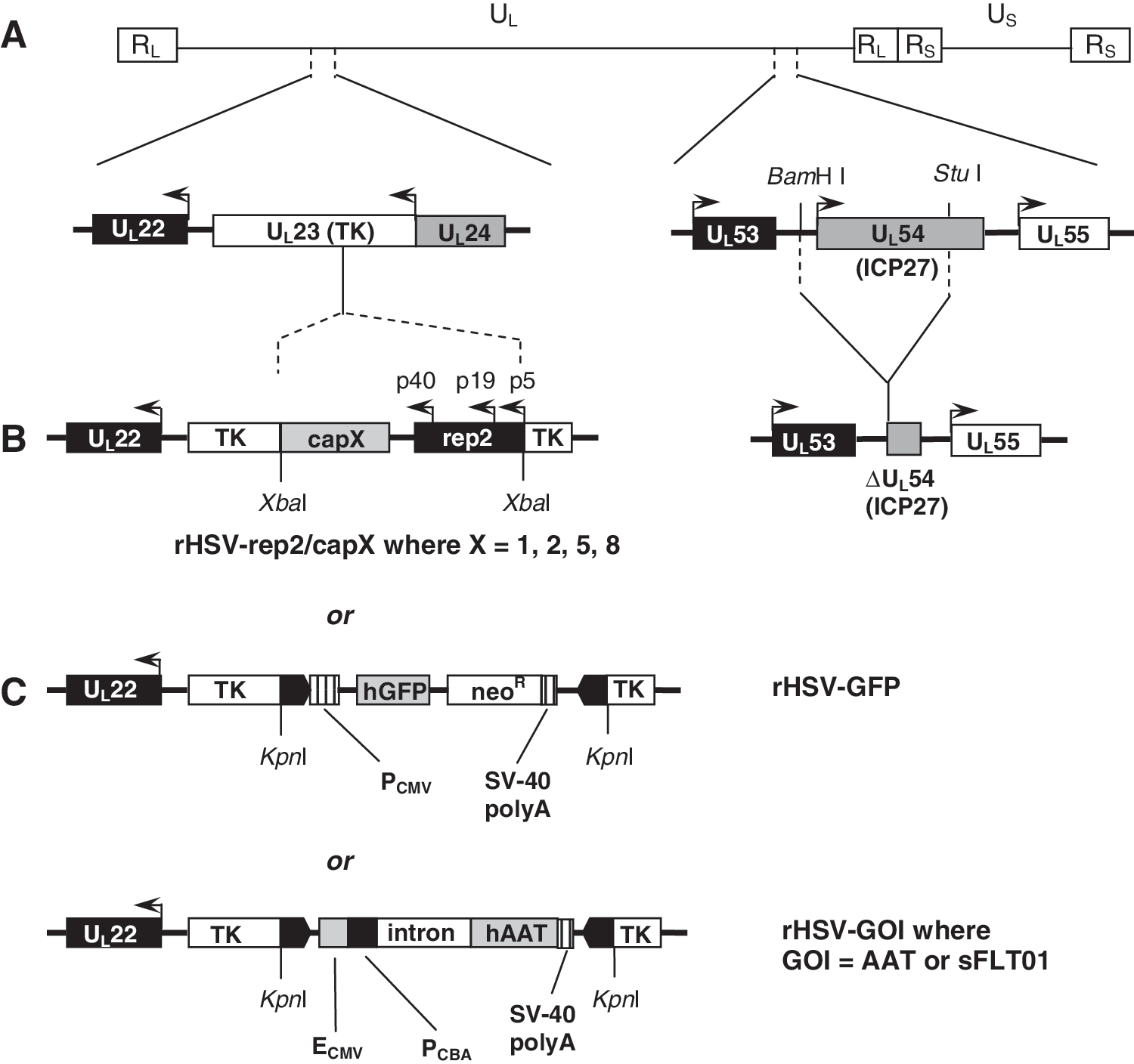
The relevant genetic sequences of rHSV-rep2/capX and rHSV-GOI used for rAAV production. (

Optimization of harvest timing and infection density for rAAV production in sBHK cells. (
We next examined the effect of cell density at the time of infection on rAAV vector production during rHSV coinfection of sBHK cells. sBHK cells were grown in 25-ml spinner cultures to the densities indicated in Fig. 2B and coinfected with rHSV-rep2/cap2 (MOI of 12) and rHSV-GFP (MOI of 2). The cultures were harvested 24 hr postinfection and processed by in situ lysis as described in Material and Methods. The rAAV2/GFP specific yields were insensitive to infection densities between 1.6 × 106 and 3.8 × 106 VC/ml (Fig. 2B), permitting 2- to 3-fold increases in volumetric productivity by increasing the cell density at the time of infection.
Finally, the input MOI for rHSV vector coinfection production of rAAV was examined. The rHSV-rep2/capX MOI of 12 used for HEK-293 cells required potentially prohibitive quantities of rHSV for optimal rAAV production, making rHSV MOI reduction a principal consideration in scale-up (Kang et al., 2009). Here, we did simultaneous coinfection of adherent HEK-293 cells in 75-cm2 flasks and sBHK cells in 25-ml spinner flask cultures at various MOI ratios, using rHSV-rep2/cap1 and rHSV-AAT vectors to produce rAAV1/AAT. The rHSV-rep2/cap1 MOI was 12, 8, or 4, whereas the rHSV-AAT MOI was held constant at 2. As expected, adherent HEK-293 cell rAAV1/AAT production was sensitive to the rHSV-rep2/cap1 MOI, decreasing from 74,600 to 42,100 DRP/cell with an MOI decrease from 12 to 4 (Fig. 3). By contrast, rAAV1/AAT production in sBHK cells was insensitive to rHSV-rep2/cap1 vector MOI inputs of 12, 8, and 4, resulting in rAAV1/AAT specific yields of 77,200, 93,400, and 85,100 DRP/cell, respectively (Fig. 3). As expected, continued reduction of the rHSV-rep2/cap1 MOI to 2 and 1 at a constant rHSV-AAT MOI of 2 showed marked reduction in the rAAV1/AAT specific yield, resulting in only 31,400 and 17,800 DRP/cell, respectively. Furthermore, a 4:2 MOI ratio (rHSV-rep2/cap1:rHSV-AAT) resulted in about 80% of the rAAV1/AAT DRP/cell production observed for a 5:5 MOI ratio (data not shown).

rAAV1/AAT production as a function of rHSV-rep2/cap1 vector MOI. The rHSV-rep2/cap1 MOI was varied (12, 8, or 4) while holding the rHSV-AAT vector MOI constant at 2 in 75-cm2 flask cultures of HEK-293 cells (1 × 107 cells, open columns) and 25-ml spinner flask cultures of sBHK cells (1 × 106 VC/ml, shaded columns). Error bars indicate standard deviation; n = 3 for spinner flask sBHK rAAV production and n = 4 for 75-cm2 flask HEK-293 rAAV production.
Production of multiple rAAV serotype–transgene combinations
Production of rAAV serotypes 1, 2, 5, and 8 in sBHK cells was investigated by coinfection of the corresponding rHSV-rep2/capX vector at an MOI of 4 with either rHSV-AAT or rHSV-sFLT01 at an MOI of 2 (Ye et al., 2006). The results are shown in Fig. 4, and demonstrate that high specific yields of rAAV (DRP/cell) can be consistently achieved in sBHK cells independent of the AAV serotype and packaged transgene, without genetic modification of either the AAV genes or the producer cell line. Furthermore, the rAAV DRP/ip ratios were low when infectivity of representative samples was determined in an end-point dilution TCID50 assay. rHSV coinfection-generated rAAV1/AAT, rAAV2/sFLT01, and rAAV5/sFLT01 viral lysates had average DRP/ip ratios of 110 ± 30, 6 ± 4, and 82 ± 25 to 1 (n = 3 for each), respectively, with the difference between serotypes most likely reflecting their in vitro infectivity variation (see Discussion). The DRP/ip ratio generated here for the prototypical rAAV2 (6 ± 4) is highly favorable when compared with other production methods, such as transient transfection (Aucoin et al., 2008).

rAAV production yields for various serotype–transgene combinations on sBHK cells. sBHK cells (1–2 × 106 VC/ml) in 25-ml spinner flask cultures were coinfected with rHSV-rep2/capX (where X is AAV cap serotype 1, 2, 5, or 8) and rHSV-GOI (where GOI is either AAT or sFLT01) at MOIs of 4 and 2, respectively. Error bars indicate the standard deviation and n = 5, 8, 6, and 3 for rAAV1/AAT, rAAV2/sFLT01, rAAV5/sFLT01, and rAAV8/AAT, respectively.
Bioreactor production of rAAV
The production of rAAV in sBHK cells by rHSV coinfection was executed in disposable bioreactors to exploit the increased scalability of suspension cells relative to adherent HEK-293 cell rAAV production. Initial sBHK growth and rAAV production experiments were performed in 1-liter working volume Sartorius BIOSTAT CultiBag RM Wave disposable bioreactors and resulted in high rAAV specific yields, using the optimized spinner flask production parameters. Cells were seeded at 2.5 × 105 VC/ml and fed with a bolus of 300 ml of 5 × DMEM (containing 1× salts) to prevent

Disposable bioreactor production of rAAV1/AAT by rHSV coinfection. Error bars indicate the standard deviation for 1-liter (n = 3), 10-liter (n = 4 with combination A of rHSV stocks), and 10-liter (n = 3 with combination B of rHSV stocks) working volume scales.
rAAV production was scaled to 5- and 10-liter disposable bioreactors and resulted in similar cell growth and rAAV specific yields as at the 1-liter scale. The initial seed volume was 2 or 4 liters and the cultures were diluted to 5 or 10 liters, respectively, at 24 hr postseeding. Cells were grown (doubling time of 12 ± 1 hr, n = 7) to a density of 1.6–2.0 × 106 VC/ml, coinfected with rHSV, and medium was exchanged 2 hr postinfection to DMEM lacking FBS. The average specific yields of 69,000 ± 7500 DRP/cell (n = 4) and 113,000 ± 16,400 DRP/cell (n = 3) for 10-liter runs (Fig. 5) were similar to 25-ml spinner flask and 1-liter disposable bioreactor specific yields, resulting in volumetric productivities of 1.3 × 1014 ± 2.0 × 1013 and 2.4 × 1014 ± 5.5 × 1013 DRP/liter. The approximately 2-fold variation in 10-liter disposable bioreactor specific yield was rHSV stock specific, as indicated in Fig. 5. Infectivity was determined for several 10-liter bioreactor productions of rAAV1/AAT and resulted in a DRP/ip ratio of 210 ± 60 (n = 5), within 2-fold of the 25-ml spinner flask results.
Medium exchange was accomplished by one of three methods: centrifugation, flat sheet tangential flow filtration (TFF), or integrated membrane perfusion of the reactor, the latter two being relevant to scale-up. Cells grown in 5-liter disposable bioreactors were collected by centrifugation (640 × g, 4 min) and resuspended in DMEM lacking FBS. The recovery of viable cells was 84 ± 11% (n = 10). A scalable process for medium exchange in 5- and 10-liter disposable bioreactors was pursued initially by flat sheet TFF in which the postinfection volume was reduced 5- to 10-fold and then diluted to the original volume with DMEM lacking FBS. The TFF medium exchange operational parameters are listed in Materials and Methods. Recovery of viable cells by TFF was 99 ± 1% (n = 5). Medium exchange was also examined using 10-liter disposable bioreactors with integrated perfusion membranes (1.2 μm), permitting medium exchange without additional equipment. Two to 4 hr postinfection, the bioreactor volume was reduced 3-fold by removing medium through the perfusion membrane (100 ml/min). The concentrated cell culture was then perfused (100 ml/min) at constant volume with 3 volumes of DMEM lacking FBS and then diluted back to the initial volume with the same. The recovery of viable cells was 93 ± 4% (n = 7).
Typical cell growth, viability, and ammonium concentrations for 10-liter disposable bioreactor sBHK cell growth and rAAV production by rHSV coinfection are shown in Fig. 6. Viability at harvest was typically 90–93%. Contrary to a previous report, cell growth did not appear to be substantially inhibited by ammonium accumulation of nearly 4 mM (Christie and Butler, 1999). Christie and Butler have reported that ammonium accumulation as low as 3 mM can significantly inhibit sBHK growth in ammoniagenic medium; however, no apparent cell growth inhibition was observed here (e.g., constant doubling time throughout the cell growth period).

Typical growth, viability, and ammonium concentrations during 10-liter bioreactor sBHK cell cultivation and rAAV production by rHSV coinfection. Arrows indicate time of (
Purification and in vivo evaluation of rAAV1/AAT 10-liter production runs
rAAV produced in 10-liter Wave bioreactors was prepared for column chromatography by clarification followed by concentration via TFF and buffer exchange via diafiltration (DF). Clarification was accomplished via depth filtration (1.2 μm) followed by dead end filtration (0.8/0.45 μm) and resulted in an average recovery of 98 ± 4% (n = 10). TFF and DF were executed with both hollow fiber filter (HFF) cartridges (0.2 or 1.15 m2) and flat sheet membranes (4 × 0.1 m2 membranes). Protein was routinely observed to precipitate during HFF DF, regardless of scale. As a result, recovery of rAAV by HFF TFF and DF was correspondingly low (Table 1). Flat sheet TFF was also examined for concentration and buffer exchange of rAAV and found to result in substantially higher recovery of rAAV, without protein precipitation, when the same operational parameters (recirculation rate, TMP, etc.) as used in HFF TFF and DF were employed (Table 1). The large protein reduction (29-fold) and standard deviation (σ = 16) observed during HFF TFF/DF was most likely a result of the protein precipitation observed in the diafiltered product. Flat sheet TFF/DF resulted in high recovery of vector (96%) with a substantial reduction in total protein (7.9-fold).
Abbreviations: DF, diafiltration; TFF, tangential flow filtration.
n = 6.
In vitro and in vivo evaluation of purified rAAV1/AAT vector generated nearly identical results to those previously seen for rHSV-produced rAAV1/AAT. Clarified, concentrated, and buffer-exchanged vector was purified by two-column chromatography, followed by buffer exchange into lactated Ringer's solution by TFF/DF. The resulting purified vector was characterized by AAV- and HSV-specific assays, and administered to C57BL/6 mice (1 × 1011 DRP/mouse) to ascertain biological activity. Table 2 shows the results for the in vitro and in vivo evaluation of purified rAAV1/AAT produced by the rHSV coinfection of a 10-liter Wave disposable bioreactor sBHK culture. The residual HSV DNA (11 ng/ml) and protein (3.2 ng/ml) were nearly identical to the values obtained for rAAV1/AAT produced by rHSV coinfection of adherent HEK-293 cells in cell factories and purified by three-column chromatography (Kang et al., 2009). In addition, the serum hAAT level observed by 1 month in C57BL/6 mice (2.8 × 104 ng/ml serum) was within the experimental error of that previously observed for rAAV1/AAT made by rHSV coinfection of HEK-293 cells (3.1 × 104 ng/ml serum) at the same dose (1 × 1011 DRP/mouse) at 1 month postinjection (Kang et al., 2009).
Abbreviations: DRP, DNase-resistant particles; hAAT, human α1-antitrypsin; HSV, herpes simplex virus; qPCR, quantitative polymerase chain reaction; rAAV, recombinant adeno-associated virus.
In C57BL/6 mice 1 month postinjection of 1 × 1011 DRP, n = 6, σ = 6.7 × 103.
Discussion
Manufacturing technologies for rAAV continue to be a limiting factor in its application to gene therapy protocols. The surface area required for the culturing of adherent mammalian cell lines and subsequent rAAV production is a daunting obstacle to gene delivery protocols aimed at treatment of systemic diseases with high dosing requirements such as Duchenne muscular dystrophy or α1-antitrypsin deficiency. Historic methodologies for rAAV production that use adherent cell lines will not be able to accomplish vector production on the required scale. Therefore, manufacture of rAAV in a suspension format is paramount to the future success of gene therapy disease indications that require large-scale vector production.
Here we have extended our rHSV coinfection platform production of rAAV by application to mammalian cells adapted to grow in suspension. The sBHK cells described in this paper produced similar specific yields (DRP/cell) of rAAV as both adherent HEK-293 and adherent BHK cells when coinfected with HSV/AAV hybrid vectors (Kang et al., 2009), but sBHK cells were easily scaled in disposable bioreactors to volumes necessary for clinical manufacturing requirements. sBHK cells are attractive from a process perspective because of their short doubling time of about 12–13 hr. For example, a seed train initiated with 1 × 107 total viable cells can be expanded to a 10-liter culture with a density of 2 × 109 viable cells/liter in 5 to 6 days. The sBHK system allowed 2- to 4-fold increases in cell density relative to adherent cell culture of HEK-293 cells, resulting in a similar increase in volumetric productivity, while allowing a decrease in rHSV vector input. The 400-fold scale-up from 25-ml spinner flasks to 10-liter bioreactors resulted in the same doubling time (12 hr) and similar specific yields, depending on the rHSV stocks used. The resulting average volumetric productivity for 10-liter bioreactors runs was as high as 2.4 ×1014 DRP/liter. These data are promising for further scale-up of the rHSV coinfection of sBHK cells for production of rAAV to meet increasing clinical requirements for human gene transfer. Finally, rAAV1/AAT produced by rHSV coinfection of sBHK cells in 10-liter disposable bioreactors and purified by two-column chromatography was similar to adherent cell factory HEK-293-produced and three column-purified rAAV1/AAT with respect to both residual HSV components and biological activity of the rAAV1/AAT produced (Kang et al., 2009).
Previous reports of rAAV production in suspension-adapted mammalian cell lines have been limited to HEK-293 cells (Park et al., 2006; Durocher et al., 2007; Feng et al., 2007; Hildinger et al., 2007) and the A549 packaging cell line, which expresses the AAV rep and cap genes (Farson et al., 2004). The HEK-293 cell systems employed the transient transfection method, which remains technically difficult to execute, results in low specific yields (DRP/cell), and contaminates rAAV with microgram quantities of exogenous DNA. A disadvantage of the A549 packaging cell line platform is that a new packaging cell line must be developed for each serotype–gene-of-interest combination, and often requires large numbers of potential clones to be screened (e.g., 1 rep/cap-positive cell per 1300 cells approximately) (Farson et al., 2004).
The sBHK rAAV production platform resulted in equivalent rAAV specific yields as adherent HEK-293 cells, but used only one third of the rHSV-rep2/capX vector input and was amenable to scale-up. The Poisson distribution is often used to characterize viral infection probability; however, it does not account for transport resistances or cell–virus contact limitations observed in T-flask culture of mammalian cells, making its application to adherent HEK-293 cells difficult. In addition, the kinetics of infection with multiple viruses can increase the difficulty in predicting viral infection by Poisson analysis even in suspension culture (Mena et al., 2007). Therefore, we employed an empirical approach to rAAV specific yield optimization in the sBHK system via MOI (total and ratio) modulation. An MOI ratio of 12:2 for the rHSV-rep2/cap2 and rHSV-GFP vectors, respectively, resulted in the maximal specific yield (ip/cell) of rAAV2/GFP in adherent HEK-293 cell culture (Kang et al., 2009). Reduction in the rHSV-rep2/capX MOI from 12 to 4 on sBHK cells did not reduce the specific yield of rAAV1/AAT (Fig. 3), and permitted the total MOI to be reduced from 14 to 6, or a 60% decrease. This most likely reflects the rHSV vector transport kinetics in the respective cultures, since the Poisson distribution predicts greater than 98% of cells should still be infected at an MOI of 4. The MOI ratio of 4:2 for rHSV coinfection production of rAAV1/AAT produced about 80% as much rAAV1/AAT as at a 5:5 MOI ratio (data not shown), and agreed closely with theoretical expectation with respect to infected cells (about 85%) because these MOIs would be predicted to infect 98 and 86% of the cell population, respectively. The MOI ratio of 4:2 was attractive from an rHSV raw material usage and purification burden standpoint, permitting the total rHSV MOI to be reduced 40%, from 10 (5:5) to 6 (4:2), with only a 20% loss of productivity (DRP/cell).
rHSV coinfection of sBHK cells for rAAV production compares favorably with the baculovirus infection production of rAAV in suspension insect cells. Cecchini and colleagues demonstrated production of 2 × 1014 DRP/liter at a 10-liter scale in a stirred-tank bioreactor at 72 hr postinfection, using three recombinant baculoviruses (Cecchini et al., 2008). By comparison, the average volumetric productivities we achieved with the rHSV coinfection of suspension BHK cells in 10-liter disposable bioreactors was as high as 2.4 × 1014DRP/liter; however, fewer recombinant viruses were required and the sBHK cells grow faster than insect cells. Another advantage of the rHSV coinfection platform is that all AAV capsid serotypes inserted into the rHSV-rep2/capX vector tested to date remain infectious; unlike the baculovirus system, which requires alteration of the capsid sequences for serotypes other than serotype 2 to remain infectious on mammalian cells. For example, rAAV1 produced by baculovirus infection of insect cells requires replacement of the N-terminal portion of VP1 with the analogous portion from serotype 2 to generate a functional phospholipase domain required for infectivity (Kohlbrenner et al., 2005). rAAV vectors of various serotypes were produced here by rHSV coinfection of sBHK cells at a low DRP/ip ratio, with the differences between serotypes most likely reflecting the differences in infectivity on the HeLaRC32 cell line used for infectious titering. This difference is exemplified by comparing the infectivity between serotypes 2 and 5, where expression of the platelet-derived growth factor receptor (α or β) was previously required for increased infectivity of serotype 5 on HeLa cells (Di Pasquale et al., 2003).
The advent of suspension rAAV manufacture provides a method for generating therapeutic quantities of vector for the treatment of a variety of diseases, inclusive of systemic vector applications requiring high DRP doses. Packaging/producer cell lines that use adenovirus infection and insect cell lines that use baculoviruses have previously been employed for suspension rAAV production. We have now extended this list to include the rHSV coinfection of sBHK cells for suspension production of rAAV, demonstrating high specific and volumetric productivities, short production cycle times, and substantially reduced helper vector requirements. These improvements should establish rHSV coinfection production of rAAV as another important production technology for preclinical and clinical rAAV gene therapy protocols. Future research will focus on removal of animal-derived products from the upstream process and additional characterization of rHSV stocks to ascertain attributes (particle-to-infectivity ratio, concentration, formulation, etc.) that enhance or diminish rAAV production.
Footnotes
Acknowledgment
The authors thank Ms. Robin Griffis for technical discussions and contributions to this work.
Author Disclosure Statement
The authors all hold share options in AGTC and have a conflict of interest to the extent that this work potentially increases their personal financial interests.