
Abstract
An elevated level of hypoxia-inducible factor 1 (HIF-1) is common in solid tumors and correlates with poor prognosis. Therefore, targeting of HIF-1 presents an appealing approach for cancer therapy. In this study, we developed an adenoviral vector carrying a fusion of human WW domain-containing oxidoreductase (hWWOX) and the HIF-1α oxygen-dependent degradation domain (ODD) under the control of a synthetic human recombinant telomerase reverse transcriptase promoter (hrTRTP). Luciferase reporter assay showed elevated promoter activity of the synthetic hrTRTP in tested tumor cell lines, but not in WI-38, a nontransformed cell line. Furthermore, adenoviral hrTRTP-hWWOX-Linker-ODD (Ad-TWLH) expression induced apoptosis in a variety of human cancer cell lines under hypoxic conditions dose dependently. Importantly, Ad-TWLH injection into xenografts of A549 tumor cells dramatically reduced tumor size in vivo. Western blot and immunohistochemistry assays also confirmed that hWWOX-Linker-ODD fusion protein was expressed in the A549 xenografts. And the protein level in center part was much higher than that of the circumjacent area. In conclusion, our dual-regulated adenovirus specifically induced apoptosis in human cancer cell lines under hypoxic conditions in vitro and repressed ectopic xenograft tumor growth in vivo, thus providing a novel strategy for hypoxia-targeted cancer gene therapy.
Introduction
WW domain-containing oxidoreductase (WWOX) was first identified in the chromosome fragile region 16q23.3–24.1, which was commonly affected in breast cancer (Bednarek et al., 2000), and subsequent studies indicated WWOX to be a potential tumor suppressor gene in ovarian cancer and other cancer types (Paige et al., 2001). WWOX contains two WW domains and a short-chain dehydrogenase/reductase-homologous domain. It physically interacts with p53, FADD, and TRADD to activate caspase pathways and induces apoptosis synergistically (Chang et al., 2001; Chang, 2002). WWOX also quenches proto-oncogenes such as c-Jun (Chang et al., 2003) and Ap-2γ (Aqeilan et al., 2004) by restraining their partitioning to the cytoplasm. Recently, adenoviral hormone-inducible WWOX expression has been used in cancer gene therapy for lung cancer cell lines (Fabbri et al., 2005). A major drawback of this system is that cytotoxicity of WWOX nonspecifically affects all transduced cells. Therefore, highly restricted WWOX expression has the potential to improve the safety profile of clinical cancer gene therapy vectors.
Human telomerase reverse transcriptase (hTRT) is a component of the telomerase complex. Because increased telomerase activity is ubiquitous in tumor cells, the hTRT promoter (hTRTP) can be utilized as a tumor-selective transcriptional control element for targeted cancer gene therapy. In this study, we developed a tumor-specific adenoviral vector dual-regulated at the transcriptional and posttranslational levels, to restrict hWWOX expression to hypoxic tumor cells. To achieve this, the HIF-1α ODD was fused to the C terminus of hWWOX. The expression of the fusion gene was controlled by a synthetic human recombinant telomerase reverse transcriptase promoter (hrTRTP), consisting of three copies of the proximal region of hTRTP in tandem. We showed that the synthetic hrTRTP has tumor-specific activity. Further, we showed that the hWWOX-Linker-ODD protein was stabilized under hypoxic conditions and activated caspase-dependent apoptosis. Significantly, adenovirus-mediated hWWOX-Linker-ODD expression (Ad-TWLH) induced tumor cell apoptosis in vitro and inhibited ectopic tumor xenograft growth in vivo. Our study demonstrated that this tumor-specific, hypoxia-regulated adenoviral vector showed promise for cancer gene therapy.
Materials and Methods
Cell culture and hypoxic treatment
Human cancer cell lines (A549, Hela, HepG2, and Hep3B), human embryo lung epithelial cell line (WI-38), and human embryonic kidney cell line (293T) were purchased from the American Type Tissue Culture Collection (ATCC, Manassas, VA). Cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% Fetal Bovine Serum (Hyclone, Waltham, MA), 100 U/ml penicillin, and 100 μg/ml streptomycin in a standard 37°C incubator with 5% CO2. For hypoxic culture, cells were plated at least 24 hr prior to transfer into a 37°C incubator with 1% O2, 5% CO2, and 94% N2 (3111; Thermo Forma, Rockford, IL) for the indicated time. In some experiments, cells were treated with 100 μg/ml cycloheximide (CHX) for the indicated time.
Plasmid construction and transfection
A 207 bp of hTRTP was cloned by polymerase chain reaction (PCR; forward primer: 5′-CAGTGAATTCCAGGACGCGCTCCCACGTG-3′; reverse primer: 5′-CAGTGGATCCCCAGGGCTTCCCACGTGCGCAGCAGGACGCAGGGCGGGGCCGCGGAAAG-3′) from pGL3-hTRTP as a single copy, which contained the core promoter region of E-box-GCislands-E-box sites and was cut with EcoRI (GAATTC) and BamHI (GGATCC) and then inserted into the pBlueScriptII vector. Two more copies of hTRTP were ligated downstream of the first copy of hTRTP. Synthetic hrTRTPs with different copy numbers of core region were then subcloned into the pGL3-Basic vector. These reporter plasmids were named as pGL3-Basic-TRTP207-1, pGL3-Basic-TRTP207-3, pGL3-Basic-TRTP207-5, and pGL3-Basic-TRTP207-3-TATA according to copy number of hTRTP core region and TATA box, respectively. hWWOX was cloned with primers according to hWWOX mRNA sequence (GenBank no. AF211943) from human fetal spleen cDNA (forward primer: 5′-ACGAAGCTTTTGGAGCGGGAGTGAGT-3′; reverse primer: 5′- AGCGGATCCGTTGGAGGGACATTTGG-3′) and then cut with HindIII (AAGCTT) and BamHI (GGATCC) and inserted into the pcDNA3 vector. The other reverse primer for hWWOX was designed for the hWWOX-Linker-ODD fusion gene, which lacked stop codon (reverse primer: 5′-AGCGGATCCGCCGGACTGGCTGCCAAG-3′). ODD was cloned from HepG2 cDNA with correspondent primer pair (forward primer: 5′-GACGGATCC
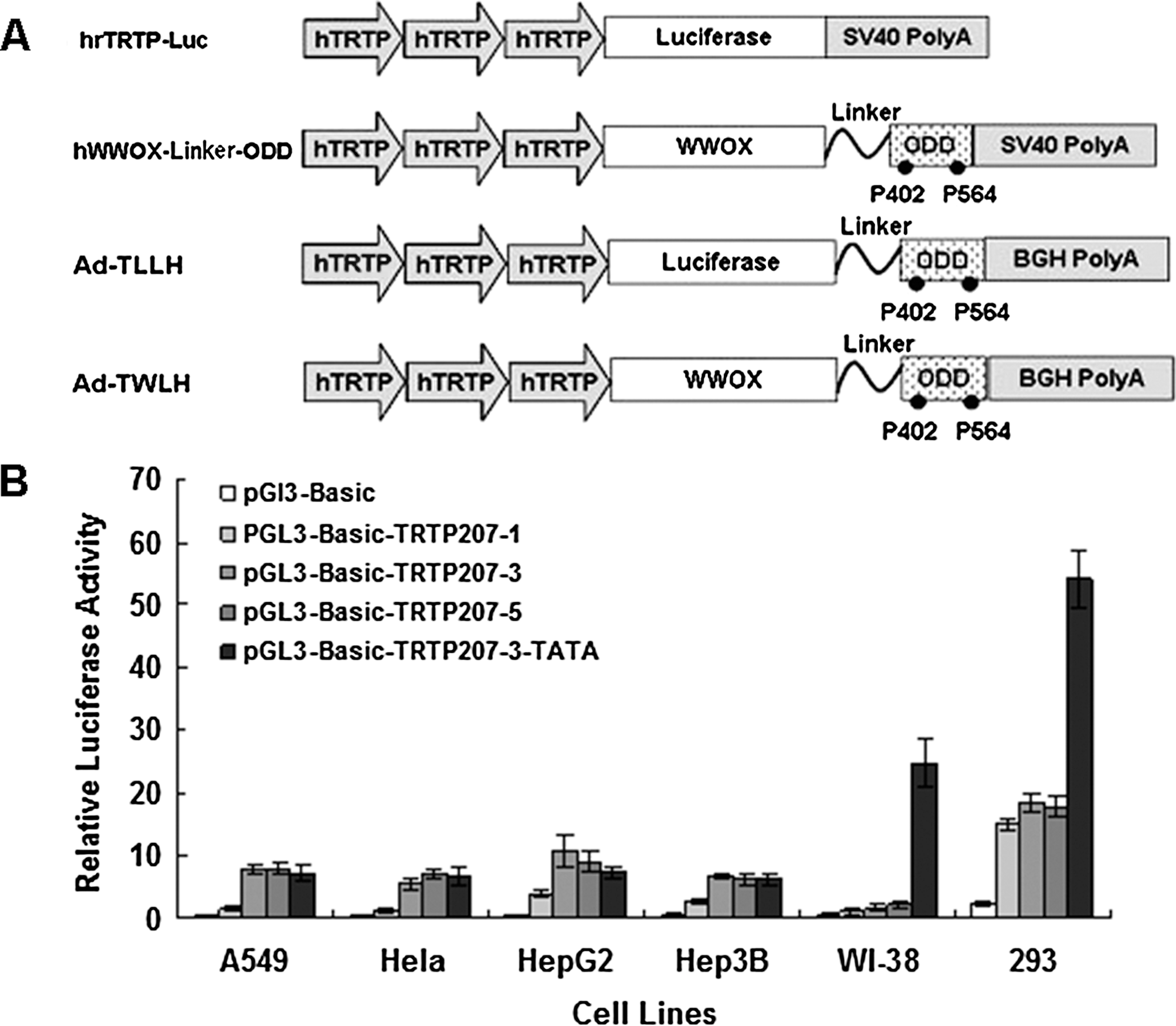
Construction of tumor-specific, hypoxia-regulated WW domain-containing oxidoreductase (WWOX)-expressing adenovirus vector. (
Human tumor cell lines were seeded in 24-well plates for 24 hr and were tranfected using Lipofectamine2000 (Invitrogen, Carlsbad, CA). For the luciferase assay, renilla reference plasmid was cotranfected as transfection efficiency control. The luciferase assay was done 24–48 hr after transfection using Dual Luciferase Reporter Assay System (Promega, Madison, WI) according to the user manual. Briefly, 20 μl of cell lysates were mixed with 50 μl of luciferase substrates and the luminal intensity was measured by luminometer (LB9507; Berthold Lumat, Bad Wildbad, Germany). Then, Stop and Glo reagent was added to the same sample to determine renilla luciferase activity.
Adenovirus preparation
Replicative-deficient adenovirus vectors were constructed with AdEasyXL adenoviral expression system (Stratagene) and virus was made according to the manufacturer's protocol. Briefly, recombined adenovirus vectors were linearized with PacI and purified with phenol–chloroform extraction. Ten microliters of linearized DNA was transfected into 293T cells with Lipofectamine2000 (Invitrogen). When the viral plaque was obvious, packaging cells were lysed to release adenovirus. The first generation of virus was used to infect 293T cells to propagate adenovirus. Then, 5 × 108 293T cells were harvested and lysed with 0.05% sodium deoxycholate. After DNaseI digestion, supernatants were subjected to cesium chloride (CsCl) super centrifugation at 45,000 rpm for 90 min. Lower adenovirus band was collected and dialyzed in PBS containing 10% glycerol overnight at 4°C. The titer of concentrated adenovirus was determined by the end-point dilution method mentioned in manufacturer's protocol. Purified adenovirus was stored at −70°C.
Western blot analysis for hWWOX-Linker-ODD expression
A549, Hela, HepG2, and Hep3B cells were seeded in six-well plates at a density of 5 × 105 cells per well and then cultured for 24 hr. Cells were transfected with pcDNA3-hWWOX-Linker-ODD and then incubated under either hypoxic or normoxic conditions for the indicated time. Whole cell lysates were extracted using cell lysate buffer (50 mM Tris-HCl [pH 7.4], 150 mM NaCl, 1% CHAPS (3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate), 100 μg/μl PMSF (phenylmethylsulfonyl fluoride), 1 μg/μl aprotinin, 1 μg/μl leupeptin). Equal amounts of proteins were loaded onto 10% SDS–PAGE gel and transferred to polyvinyl difluoride membrane. The membrane was blocked with 5% nonfat milk in TBS-T (pH 7.6, Tris-buffered saline with 0.1% Tween 20) and incubated with specific primary antibody overnight at 4°C. After washing, peroxidase-conjugated secondary antibody was applied. The signal was detected by Super Signal WestPico chemiluminescence system (Pierce, Rockford, IL) and developed on X-ray film. Antibodies used in this study were anti-human WWOX antibody (Chemicon, Billerica, MA), anti-caspase-3 antibody (Cell Signaling, Danvers, MA), and anti-β-tubulin antibody (Santa Cruz Biotechnology, Santa Cruz, CA).
Total RNA extraction and reverse transcriptional PCR (RT-PCR)
Total RNA was extracted with TRIzol reagent (Invitrogen) as indicated. Briefly, cells were harvested after appropriate treatment. About 5 × 106 cells were lysed with 1 ml TRIzol reagent. RNA concentration was quantified using UV absorbance at 280 and 260 nm. Five micrograms of total RNA was used for reverse transcription by SuperScriptII reverse transcription system (Invitrogen) with standard procedure. For hWWOX-Linker-ODD transcript detection, we designed a primer pair that overlaps hWWOX and ODD region (forward primer: 5′-CGACTATTGGGCGATGC-3′; reverse primer: 5′-CAGGGTCAGCACTACTTCG-3′). hGAPDH was used as internal control (forward primer: 5′-TGGGGAAGGTGAAGGTCGG-3′; reverse primer: 5′-CTGGAAGATGGTGATGGGA-3′).
Caspase activation assay
A549 cells were seeded onto 3.5-cm dishes and transiently infected with Ad-TLLH or Ad-TWLH. Infected cells were cultured under either normoxic or hypoxic conditions for 24 hr. In time-course experiment, A549 cells were infected with Ad-TWLH at a multiplicity of infection (MOI) of 50 and then incubated under hypoxic conditions for indicated time. In dose–response experiment, A549 cells were infected with Ad-TWLH at different MOIs and then incubated under hypoxic conditions for 24 hr. Cells were lysed and immunoblotted with anti-caspase-3 antibody.
Clonogenesis assay
A549 cells were seeded in 24-well plates for 5 × 104 cells per well and transfected with pcDNA3, pcDNA3-hWWOX, and pcDNA3-hWWOX-Linker-ODD, respectively. Transfected cells were replated into six-well plates at a ratio of 1:10 after 24 hr and selected with 600 μg/ml G418 for 14 days. Colonies were then stained with 0.2% crystal violet and counted.
Apoptosis assay: annexin-V-FITC (fluorescein isothiocyanate, FITC)
A549, Hela, HepG2, and Hep3B cells were plated into 24-well plates with 5 × 104 cells per well. Cells were infected with Ad-TLLH and Ad-TWLH for 24 hr at an MOI of 50. Infected cells were cultured under normoxic or hypoxic conditions, respectively, for another 24 hr. In time-course experiment, A549 cells were also infected with Ad-TWLH at an MOI of 50 and then incubated under hypoxic conditions for indicated time (three independent experiments). In dose–response experiment, A549 cells were infected with Ad-TWLH at different MOIs and then incubated under hypoxic conditions for 24 hr (three independent experiments). Cells were harvested and rinsed with PBS twice. Then early apoptotic cells were detected with annexin-V-FITC Apoptosis Detection Kit (BD Pharmingen, San Diego, CA) according to manufacturer's instructions. Briefly, cells were resuspended with 1 × Binding Buffer to 1 × 106 cells/ml. About 100 μl of cells were mixed with 5 μl of Annexin-V-FITC and 5 μl of propidium iodide (PI) and incubated for 5 min at room temperature, and then diluted with 400 μl of 1 × Binding Buffer. Labeled cells were analyzed by flow cytometry (LSRII; BD Pharmingen).
Generation of ectopic xenograft
A549 cells were trypsinized and washed with PBS twice. Cells were resuspended in PBS at 5 × 107 cells/ml. Six-week-old male BALB/c (nu/nu) mice were injected with 100 μl of aforementioned cells subcutaneously at right or left dorsal. Tumor volume = 0.5 × length × width2 (mm3). After 14 days, the average size of ectopic xenograft tumors reached 1000 mm3 and mice were divided into three groups randomly. The negative group received Ad-TLLH, the treatment group received Ad-TWLH, and H101, a p53 deficiency-dependent oncolytic adenovirus, was used as positive control. One hundred microliters of adenovirus (virus titer was about 1 × 109 pfu/ml) was injected intratumorally every 2 days for five times. Tumor volume was recorded after initiation of adenovirus injection every 4 days with vernier calipers.
Immunohistochemistry
After the Ad-TWLH injection treatment, A549 xenografts were fixed with formalin and embedded in paraffin. For immunohistochemistry, 4-μm tissue sections were incubated with anti-human WWOX antibody (diluted 1:200; Chemicon) and developed according to the standard protocol.
Statistical analysis
The data were presented as mean ± standard error of the mean. Difference between the groups was analyzed with analysis of variance or Student's t-test. Statistical significance was set at p < 0.05.
Results
Synthetic hrTRTP has increased promoter activity in tumor cells
As aforementioned, hTRTP has served as a tumor-specific promoter for cancer gene therapy. The core region of hTRTP is characterized as E-box-GC islands-E-box, which is located around 230 bp upstream of the hTRT transcription start site (Takakura et al., 1999). We cloned this proximal core region of hTRTP with PCR. We constructed a series of synthetic hTRTP (hrTRTP) with one, three, or five copies of the hTRTP core region to test the dose–efficacy relationship between hTRTP core region copy numbers and the expression level of the downstream gene (Fig. 1A). Transient transfection assays showed that the novel hrTRTP containing three copies of the core region displayed increased activity in A549, Hela, HepG2, and Hep3B cancer cell lines and low activity in WI-38 cell, an untransformed cell line (Fig. 1B). When we scaled up core region to five copies, promoter activity did not increase remarkably when compared with three copies (Fig. 1B). The promoter activity reached its plateau by three copies of core promoter region. Five copies of core region could not further enhance its transcriptional activity. According to reporter assay data in our study, we concluded that the synthetic hrTRTP with three copies of core region had optimal promoter activity.
hTRTP is highly GC rich and lacks TATA and CAAT boxes (Takakura et al., 1999). TATA box is a universal core element that recruits TATA-binding protein, RNA polymerase II, and transfactor II D (TFIID) and facilitates gene transcription initiation. To test if an extra TATA box can enhance hrTRTP activity, we ligated three copies of hTRTP core region with a nonspecific TATA box. Luciferase assay showed that TATA box could remarkably increase hrTRTP transcription activity in all tested cell lines, but the specificity of hrTRTP was significantly disrupted in the presence of TATA box as hrTRTP activity also dramatically increased in WI-38 cell line (Fig. 1B). To increase its activity while maintaining its specificity in tumor cells, we ligated three copies of core region in tandem and this synthetic hrTRTP was used for subsequent experiments.
ODD mediates inducible reporter gene expression under hypoxic conditions
To test if HIF-1α ODD could effectively induce fusion protein degradation under normoxic conditions, we made EGFP-Linker-ODD (Enhanced Green Fluorescent Protein, EGFP) and luciferase-Linker-ODD fusion proteins to test protein stability under hypoxic and normoxic conditions. A549 cells were seeded in 24-well plates for 24 hr prior to transfection with pcDNA3-EGFP-Linker-ODD or pcDNA3-luciferase-Linker-ODD plasmids. Cells were cultured under hypoxic or normoxic conditions. EGFP was observed under fluorescent microscopy. Under normoxic conditions, EGFP almost disappeared after 42 hr, whereas under hypoxic culture conditions, EGFP levels remained stable (Fig. 2A). To measure luciferase activity, cells were harvested 12 hr after transfection. Cell lysates were subjected to Dual Luciferase Assay System for luciferase activity analysis. 12 hr after transfection, the luciferase activity of fusion protein was approximately two times higher in hypoxia-treated cells than that in normoxia-treated cells. At longer incubation time, luciferase activity from normoxia-cultured cells decreased further and was almost exhausted at 42 hr, while the luciferase activity remained unchanged under hypoxic conditions (Fig. 2B). Thus the ODD fusion reporter protein could be effectively degraded by built-in cell regulation machinery under normoxic conditions and kept stable under hypoxic conditions.
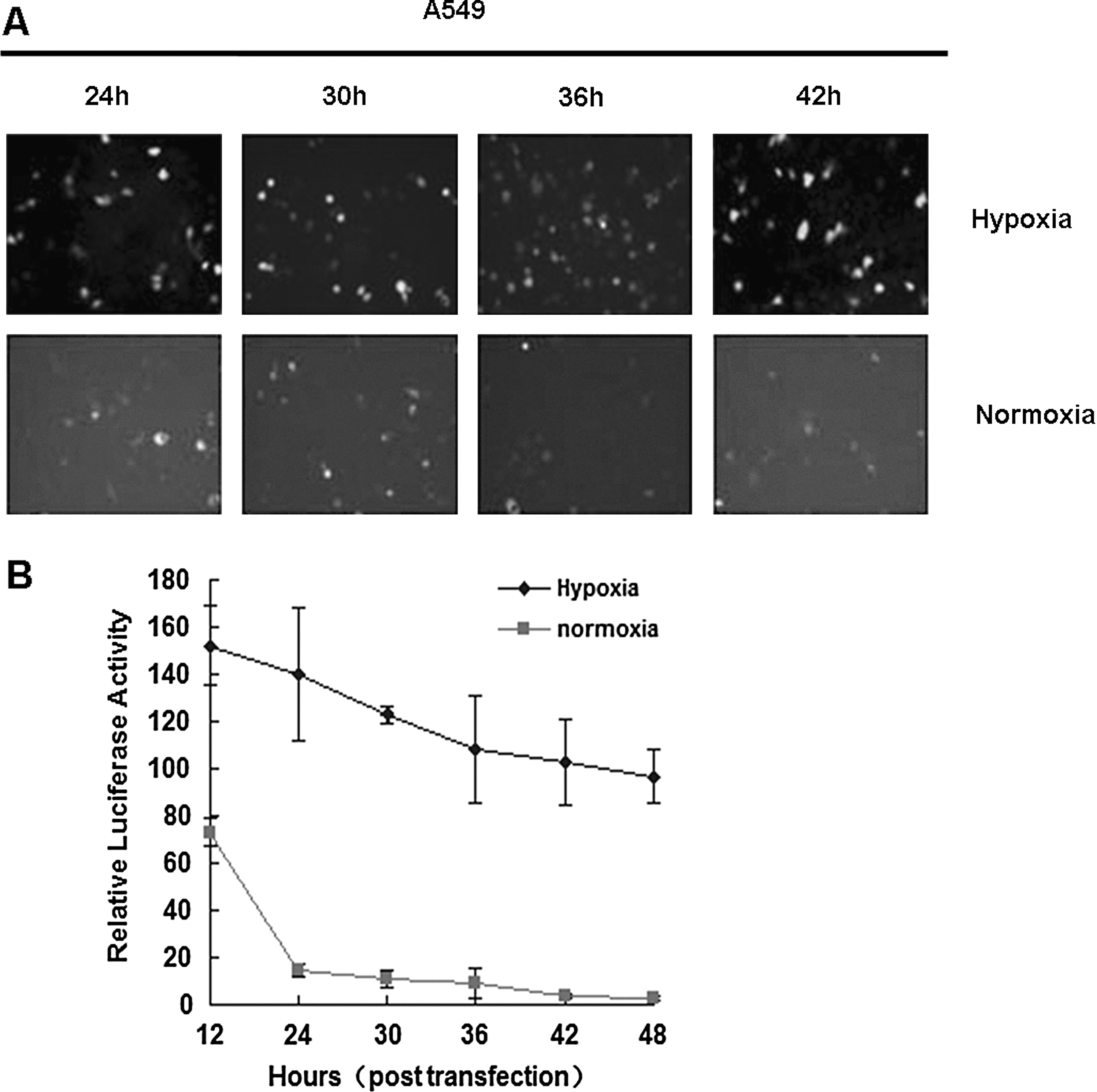
ODD-fused reporter protein is stable in hypoxic tumor cells. (
hWWOX-Linker-ODD fusion protein expression is strictly regulated posttranslationally
ODD fusion reporters showed hypoxia-specific stability. Based on this result, we proceeded to make hWWOX-Linker-ODD fusion proteins to obtain hWWOX expression only under hypoxic conditions. We investigated exogenous gene mRNA level and protein levels under different culture conditions to find out if hypoxia-specific stability occurs at the transcriptional or posttranslational levels.
A549, Hela, HepG2, and Hep3B were transiently transfected with pcDNA3-hWWOX-Linker-ODD and cultured under normoxic or hypoxic conditions for 24 hr. Then cells were harvested for total RNA extraction and RT-PCR analysis. Whether cultured in normoxia or hypoxia, the mRNA levels of hWWOX-Linker-ODD fusion gene remained the same in all cell types (Fig. 3A). The mRNA itself was stable and only regulated by the promoter.
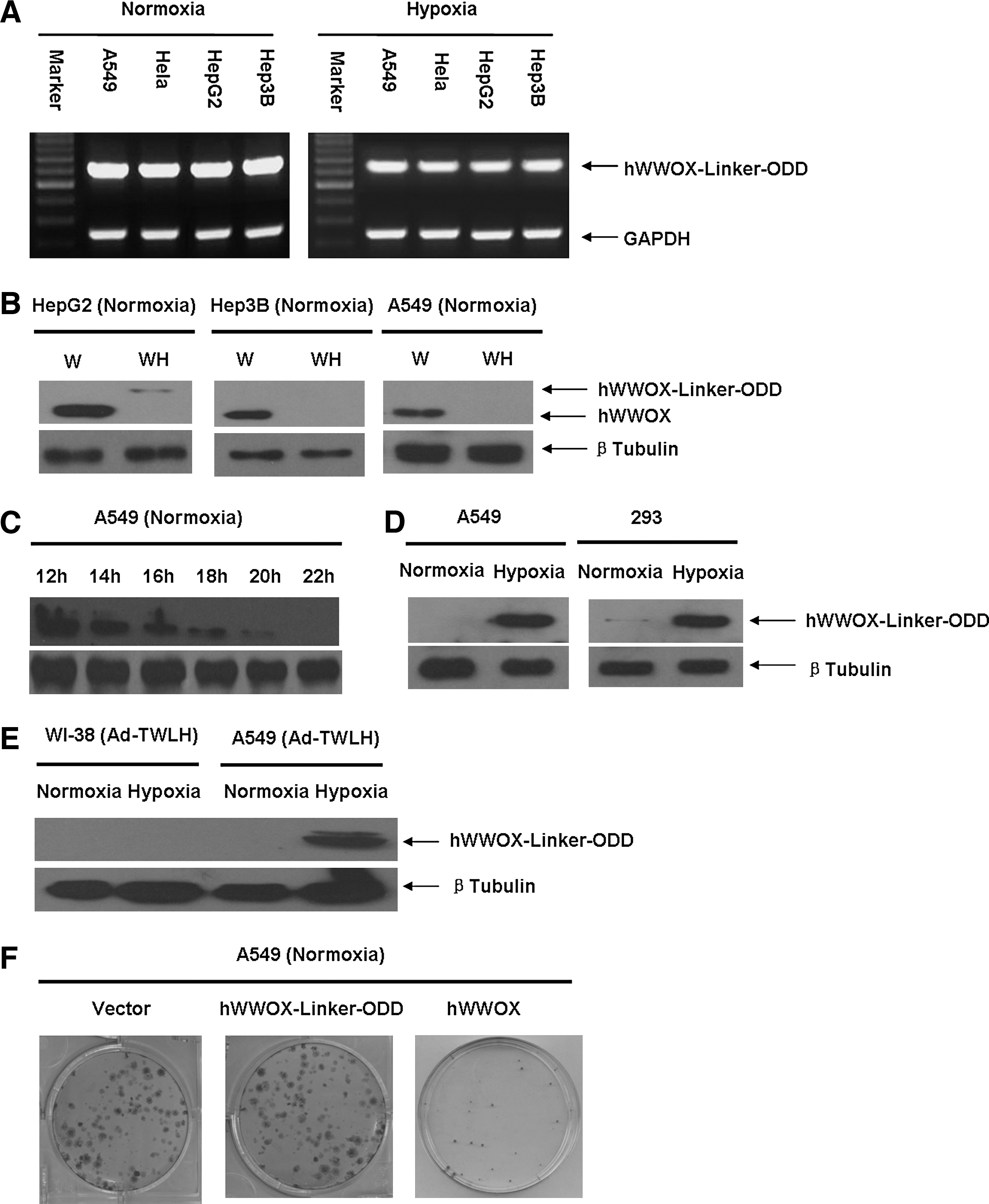
hWWOX is specifically expressed in hypoxic tumor cells. (
Next, human cancer cell lines were transiently transfected with pcDNA3-hWWOX or pcDNA3-hWWOX-Linker-ODD and then cultured under normoxic conditions. Whole cell lysates were applied to SDS-PAGE and blotted with anti-human WWOX antibody. HepG2, Hep3B, and A549 are all WWOX-negative cell lines. After transfection, the 46-kDa wild-type human WWOX was highly expressed in all three cell types under normoxic conditions, whereas the 70-kDa hWWOX-Linker-ODD fusion protein expression levels in all tested cancer cell lines were much lower than the wild-type counterpart (Fig. 3B). Therefore, hWWOX-Linker-ODD fusion protein could not accumulate within cancer cells under normoxic conditions. Then, we analyzed hWWOX-Linker-ODD fusion protein stability in A549 cells. A549 cells were transfected with pcDNA3-hWWOX-Linker-ODD and then cultured under normoxic conditions in the presence of 100 μg/ml CHX which is a potent protein synthesis inhibitor. Cells were harvested from 12 to 22 hr at an interval of 2 hr and whole cell lysates were loaded to SDS-PAGE gel and blotted with anti-human WWOX antibody. Under normoxic conditions in the presence of CHX, the fusion protein degraded progressively (Fig. 3C). In contrast, the fusion protein expression level was significantly higher in hypoxia-treated A549 cells than that in normoxic conditions (Fig. 3D).
To test if hWWOX-Linker-ODD fusion protein could inhibit A549 cell colony formation, A549 cells were transfected with pcDNA3, pcDNA3-hWWOX, or pcDNA3-hWWOX-Linker-ODD and then selected with G418 under normoxic conditions. After 14 days, A549 colonies were stained with crystal violet and counted. Under normoxic conditions, wild-type hWWOX markedly inhibited A549 colony formation. pcDNA3-hWWOX-Linker-ODD-transfected cells formed as many colonies as did the cells tranfected with empty vector (Fig. 3F). Thus, under normoxic conditions, hWWOX-Linker-ODD had little cytotoxic effect on cancer cells.
Adenovirus-mediated hWWOX-Linker-ODD expression promotes cancer cell apoptosis under hypoxic conditions
To investigate the cytotoxic effect of hrTRTP-hWWOX-Linker-ODD on cancer cells, hrTRTP-hWWOX-Linker-ODD was inserted into pShuttle and then recombined with AdEasy-1 backbone in BJ5183 using AdEasy XL adenoviral expression system (Stratagene), named Ad-TWLH. hWWOX was replaced by luciferase as a negative control, named Ad-TLLH. Adenovirus was produced by 293T cells and was purified by double CsCl density supercentrifugation. The final adenovirus titer was about 1 × 109 pfu/ml. A549, Hela, HepG2, and Hep3B cells were infected at an MOI of 50. The infection efficiency was approximately 80%. Infected cells were cultured under normoxic conditions for 24 hr and then moved to normoxic or hypoxic conditions for another 24 hr. First, we tested the stringency of the adenovirus-mediated system. An hTRT-negative cell line, WI-38, was included as control. The 70-kDa hWWOX-Linker-ODD fusion protein could be detected only in hypoxia-cultured A549 cells because of the high telomerase activity and the stability of fusion protein in hypoxic conditions. In WI-38 cells, even under hypoxic conditions, the fusion protein could not be detected. This result meant that there was no transcription in WI-38 cells because of the low telomerase activity (Fig. 3E). Next, we analyzed proapoptotic effects of Ad-TWLH. A549 whole cell lysates were harvested and immunoblotted with anti-caspase-3 antibody. Under normoxic conditions, the 17-kDa cleaved form of caspase-3 was not present in significant quantities in either Ad-TLLH or Ad-TWLH samples. Under hypoxic conditions, however, caspase-3 was greatly activated in Ad-TWLH-infected A549 cells (Fig. 4A). Infected A549, Hela, HepG2, and Hep3B cells were also subjected to annexin-V-FITC/PI analysis to detect early apoptosis represented in the lower right quadrant. In normoxic conditions, Ad-TLLH-infected cells had 1.43%, 1.9%, 2.12%, and 1.93% of early apoptosis rate in Hela, A549, HepG2, and Hep3B cells, respectively; Ad-TWLH-infected cells had 3.77%, 3.49%, 2.83%, and 3.6% of early apoptosis rate in Hela, A549, HepG2, and Hep3B cells, respectively. Hypoxia treatment itself increased the early apoptosis rate to 2.66%, 1.94%, 1.82% and 1.58% in Ad-TLLH-infected Hela, A549, HepG2, and Hep3B cells, respectively. Ad-TWLH-infected cells showed remarkable early apoptosis under hypoxic conditions in vitro: 27.11%, 32.6%, 33.1%, and 26.44% in Hela, A549, HepG2, and Hep3B cells, respectively, which were significantly higher than Ad-TWLH-infected cells under normoxic conditions (Fig. 4B). Representative flow cytometry analysis graph showed a notable shift of annexin-V-FITC signal in hypoxia-treated, Ad-TWLH-infected A549 cells (Fig. 4C). The in vitro result revealed that hypoxia could greatly stimulate the cytotoxic effects of Ad-TWLH in cancer cell lines. This cytotoxic effect was dose dependent because the early-stage apoptosis percentage increased from 0.01% to 36.99% when MOI was elevated from 0 to 200 (Fig. 5A and B). Caspase-3 activation assay further confirmed dose-dependent proapoptotic effects of Ad-TWLH (Fig. 5C). Next we checked the apoptosis onset time point. A549 cells were infected with Ad-TWLH at an MOI of 50 and then incubated under hypoxic conditions for indicated time. Annexin-V-FITC staining showed that a remarkable early apoptotic population emerged 12 hr after infection (Fig. 5D and E). Caspase-3 cleavage could be detected around 12 hr and peaked at 48 hr (Fig. 5F).
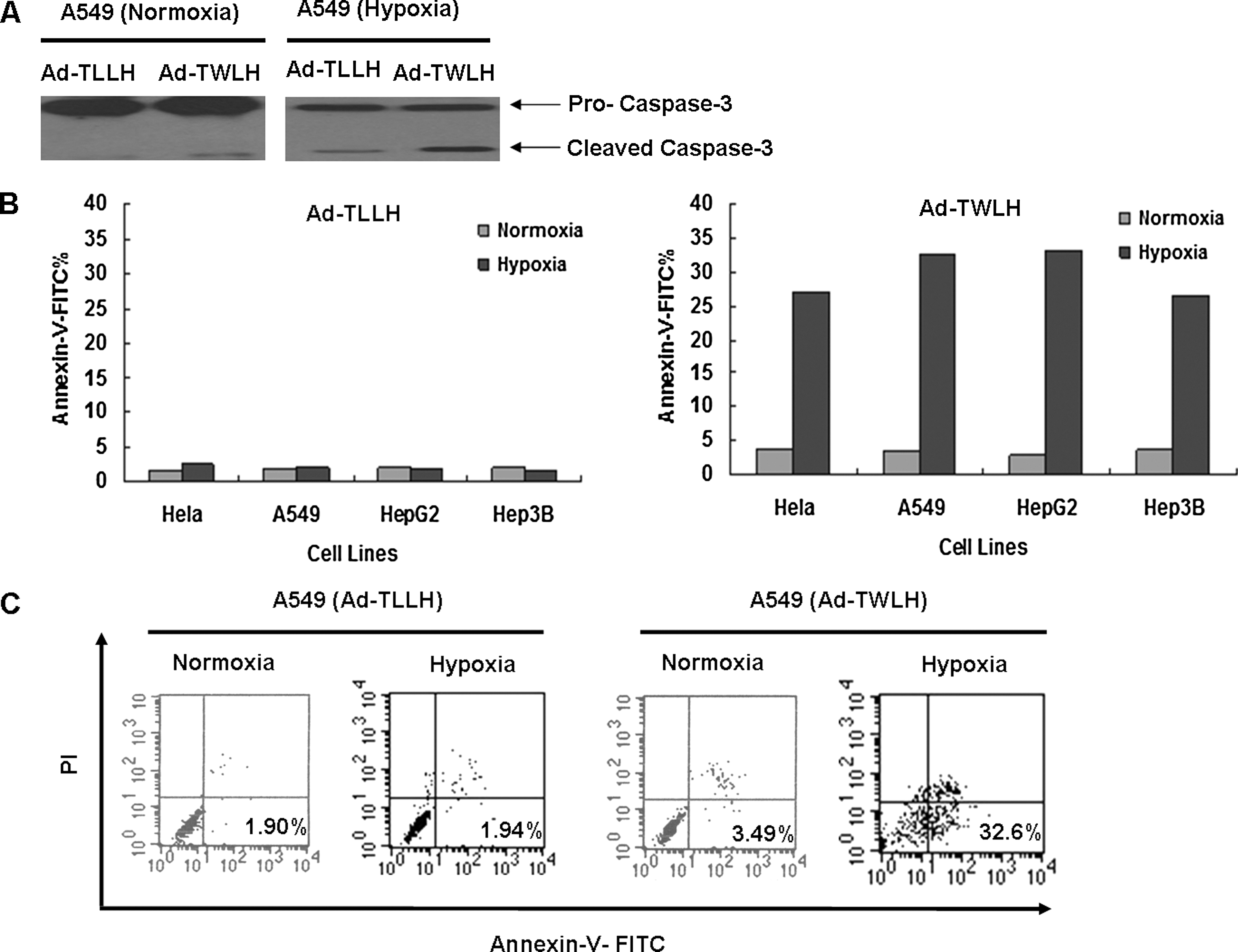
Ad-TWLH induces apoptosis in hypoxic tumor cells. (
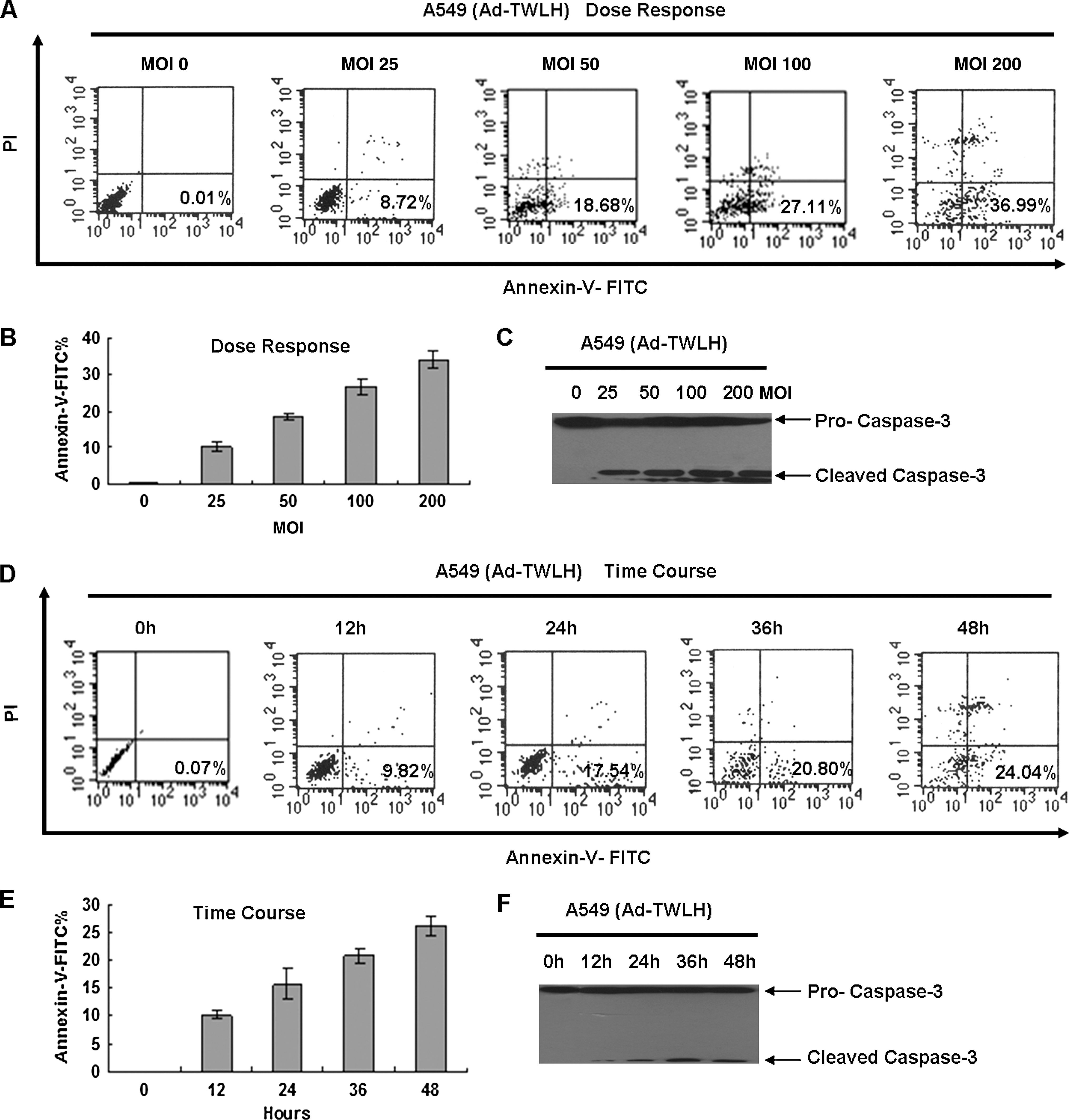
Ad-TWLH induces A549 cell apoptosis in a time-dependent and dose-dependent manner. (
Ad-TWLH significantly inhibits xenograft A549 tumor growth in vivo
To investigate the antitumor effect of Ad-TWLH in vivo, an ectopic A549 xenograft tumor model was established in BALB/c (nu/nu) mice. About 5 × 106 viable tumor cells were injected subcutaneously into the right or left dorsal area of 6-week-old male mice. After 14 days, the average xenograft tumor size reached 1000 mm3. Tumor-bearing mice were divided into three groups and received adenovirus intratumoral injections five times every 2 days. The dose of adenovirus was 1 × 108 pfu in 100 μl PBS each time. The negative control group received Ad-TLLH, the positive control group received H101, an oncolytic adenovirus (Heise et al., 1997), and the treatment group was injected with Ad-TWLH. Tumor size was measured every 4 days after the initiation of virus injection. Twenty-one days later, the average tumor sizes of the negative group, positive group, and treatment group were 3383.4 ± 787.2, 1579.8 ± 272.1, and 1302.4 ± 413.2 mm3, respectively (Fig. 6A). Statistical analysis showed that Ad-TWLH achieved significant antitumor efficacy (p = 0.00039 for negative control group, p = 0.016 for positive control group) (Fig. 6B). Ad-TWLH-injected tumor xenografts were characterized by typical necrotic appearance (Fig. 6C), compared with negative control group tumor grafts.
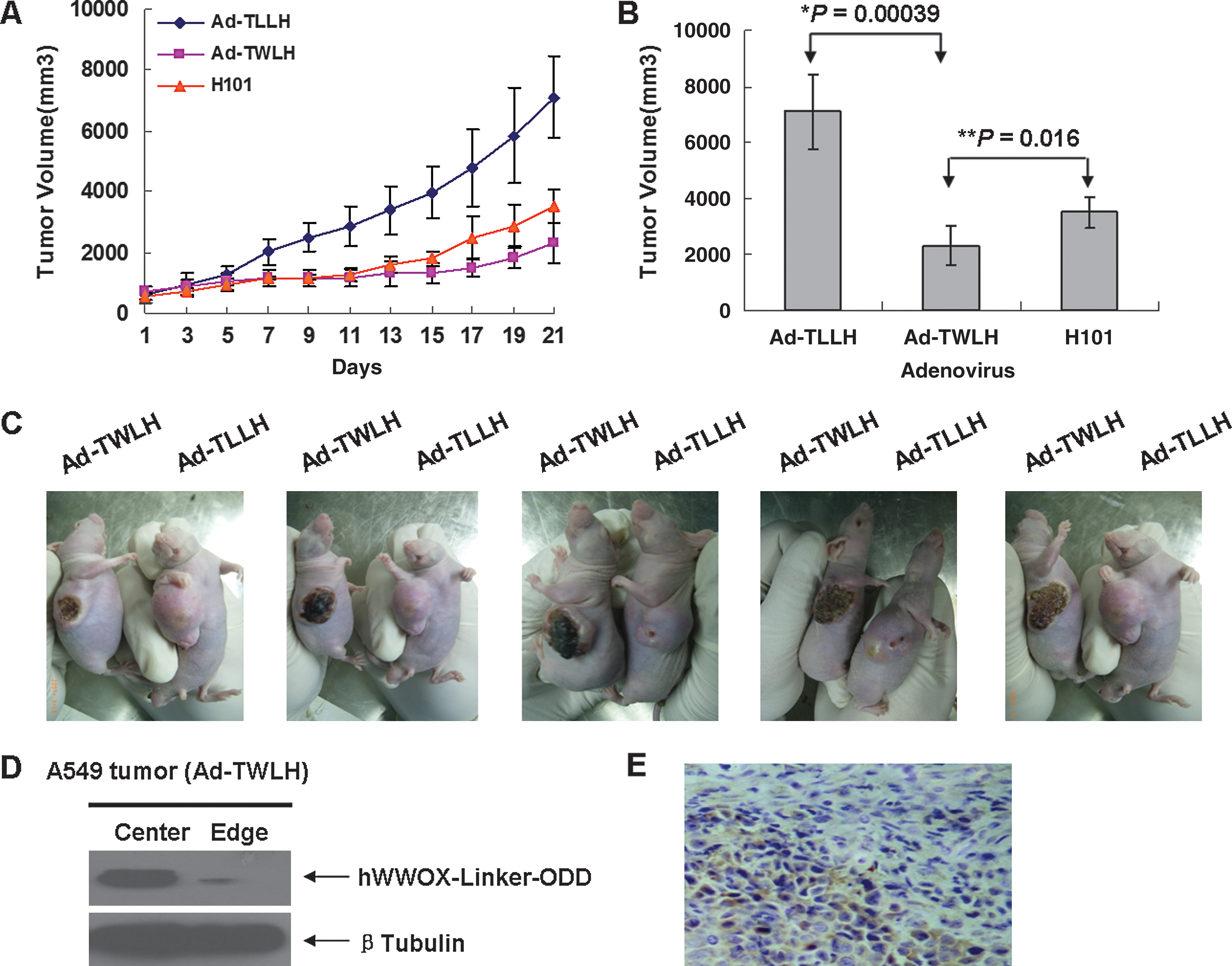
Ad-TWLH inhibits tumor growth in ectopic tumor xenograft in nude mice. (
To further validate the temporal and spacial specificity of Ad-TWLH, we separated central and peripheral portions of the xenograft tumor and subjected samples to Western blot. hWWOX-Linker-ODD was dominantly expressed at the center which was considered the hypoxic region (Fig. 6D); this was confirmed by immunohistochemistry analysis of tumor sections (Fig. 6E). Taken together, the above results showed that Ad-TWLH could effectively inhibit tumor growth in vivo.
Discussion
Hypoxia and activation of telomerase are two major features of tumors and are interesting targets for exploiting novel tumor target therapies. In this study, we took advantage of these two properties to construct Ad-TWLH, a dual-regulated adenovirus that specifically expressed a cytotoxic gene in hypoxic tumor cells.
We successfully made recombinant human TRTP (hrTRTP) with much higher activity without compromising its specificity. hTRT is the active subunit of telomerase which is positive in stem cells and over 90% of tumor cells but inactive in most normal somatic cells. hTRT expression is strictly regulated at the transcriptional level by hTRTP with a core promoter region composed of a 59-bp sequence located at −208 to −150 bp upstream of the transcriptional start site (Horikawa et al., 1999). Tumor-specific therapy strategies based on hTRTP have achieved success in the past decade (Pan and Koeneman, 1999; Abdul-Ghani et al., 2000; Gu et al., 2002; Kirch et al., 2002; Zhang et al., 2008). We cloned a 207 bp of hTRTP proximal core region, and the reporter assay showed that it specifically promoted reporter gene expression in tumor cells lines, such as A549, Hela, HepG2, and Hep3B, but not in WI-38, a normal human embryo lung epithelial cell line. Regarding the purpose of gene therapy, efficiency shares the same importance with specificity. To increase hTRTP activity, we first introduced a generic TATA box to the hTRTP core promoter. hTRTP is a TATA-free promoter with high GC content. An extra TATA box did facilitate transcription in our case, but also abolished tumor specificity. To recruit more tissue-specific transcriptional factors, we ligated three or five copies of 207 bp of hTRTP core region in tandem. Luciferase assay showed that hrTRTP with three copies of core region gained increased activity in tested cancer cell lines and maintained low activity in WI-38 cell line. However, two more copies of 207 bp did not increase hrTRTP activity. Based on this evidence, we made hrTRTP containing three copies of hTRTP core region as a useful promoter to drive target gene expression with high activity and specificity.
Poor vasculature and consequent decreased oxygen perfusion are another features in solid tumor. HIF-1α protein level is stable in most hypoxic tumor cells, which allows tumor cells to adapt to the hypoxic environment and gain predominance in proliferation and resistance to chemotherapy/radiotherapy. Under physiological conditions, ODD of HIF-1α mediates proteasome degradation. But in hypoxic conditions, the mechanism is shut off and HIF-1α is stabilized. We first evaluated two reporter genes (EGFP and luciferase) fused with ODD and found that the fusion proteins were sensitive to oxygen concentration in a visible way in A549 cells. EGFP and luciferase signals almost vanished after 36 hr in normoxic culture in the presence of CHX, a protein synthesis inhibitor (Fig. 2A and B). Then we fused the cytotoxic gene hWWOX with ODD in the same frame. Transient transfection experiments and subsequent Western blots proved that hWWOX-Linker-ODD protein levels were unstable under normoxic conditions in A549, HepG2, and Hep3B cells (Fig. 3B) despite equal transcription levels under normoxic and hypoxic conditions (Fig. 3A.). Time course degradation showed that the half-life time of fusion protein was around 16 hr in A549 cells (Fig. 3C). Decreased protein level under normoxia is regulated posttranslationally as expected. Safety is also one of the major concerns in cancer gene therapy; our clonogeneic assay showed that hWWOX-Linker-ODD-transfected cells had the same growth rate as empty vector control under normoxic conditions, and wild-type hWWOX greatly decreased cell survival (Fig. 3F). Above data showed that hWWOX-Linker-ODD was benign to cells under normoxic conditions. Another advantage of this fusion protein was that it will not induce 293T cells to apoptosis under normoxic conditions while packaging adenovirus. The packaging efficiency tremendously decreased if the exogenous protein was toxic to 293T cells. Many researches focused on the production of massive amount of adenovirus for in vivo application, such as ligand-inducible system (Fabbri et al., 2005). However, in our system, hWWOX-Linker-ODD degraded in 293T cells under normoxic conditions (Fig. 3D) and actually protected packaging cells from being damaged and ensured high titer adenovirus production.
WWOX is a WW domain-containing oxidoreductase cytotoxic protein. WWOX can operate synergistically with p53 to activate apoptosis through activation of capspase-3, -8, and -9 pathways (Chang et al., 2001, 2003) and exert more potent proapoptotic efficacy in WWOX-negative cells (Fabbri et al., 2005). In our tested tumor cells, HepG2 is p53 intact, Hep3B is p53 null, and both A549 and Hela have downregulated p53 levels. HepG2, Hep3B, and A549 are WWOX null (Fig. 3B). Early apoptosis rates are higher in HepG2 and A549 cell lines (Fig. 4B). In the future, combination therapy of WWOX and p53 is expected to obtain higher efficiency.
Next, we combined hrTRTP and hWWOX-Linker-ODD to make temporal and spatial-regulated Ad-TWLH. The specificity and inducibility of the dual-regulated adenovirus was evaluated in WI-38 and A549 cells under different oxygenation conditions. The result well matched our expectation that this virus only worked in hypoxic tumor cells (Fig. 3E). In our system, Ad-TWLH only activated caspase-3 cleavage and early apoptosis under hypoxic conditions in all tested tumor cells while exerting no effect under normoxic conditions (Fig. 4A–C). Additionally, the proapoptotic effect of Ad-TWLH was dose dependent (Fig. 5A–C) and time dependent (Fig. 5D–F).
In A549 xenograft mouse tumor models, Ad-TWLH exhibited excellent antitumoral activity by effectively depressing tumor growth, and the xenografts showed significant morphological change, whereas Ad-TLLH had no effect on tumor growth or morphological change (Fig. 6A–C). H101 is an oncolytic adenovirus featured with double deletion of E1B-55kDa and partial E3 gene. E1B deletion allows it to replicate selectively in p53null/mutant cancer cells because of absence of p53-mediated cell cycle arrest, and E3 partial deletion renders slower elimination rate by host immune system. A549, a p53lo tumor cell line, has the ability to host H101 replication. Compared with H101, Ad-TWLH had significantly increased antitumor activity (p = 0.016; Fig. 6B). A549 xenograft tumors underwent shrinking, bleeding, and even elimination. Although Ad-TWLH did not replicate, the stable expression of hWWOX-Linker-ODD in hypoxic cells contributed largely to its increasing antitumor activity. Importantly, hWWOX-Linker-ODD was only highly expressed in tumor samples from hypoxic region (center) but not normoxic region (periphery) (Fig. 6D). Immunohistochemistry staining affirmed that hWWOX-Linker-ODD was expressed preferentially at hypoxic tumor area (Fig. 6E).
In conclusion, we constructed a tumor-specific, hypoxia-regulated adenovirus, Ad-TWLH. This adenovirus had high specificity in hypoxic tumor cells and excellent antitumoral efficacy in vivo. The increased safety properties and remarkable antitumoral effects of Ad-TWLH makes it a promising strategy for cancer gene therapy.
Footnotes
Acknowledgments
This work was supported by 973 National Basic Science Research Project (2004CB518804 and 2010CB529902), the Natural Science Foundation of China (30400536 and 30973663), Basic Science Project from Shanghai Science and Technology Committee (07JC14034 and 08ZR1412500), the foundation from Shanghai Municipal Education Commission (09ZZ110 and S30205), Doctoral Foundation (BXJ0606) and Elevation Project for Young Teachers from Institutes of Medical Sciences, Shanghai Jiaotong University School of Medicine. We also thank Dr. Guang Qu, Joana Balcereck, Amy E. Campbell, and Stephan Kadauke for extensively reading the manuscript and for their precious comments.
Author Disclosure Statement
No competing financial interests exist.